











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introducción
La línea de árboles es uno de los ecotonos más vulnerables y sensibles al calentamiento global. Sin embargo, debido a que la localización de cualquier borde natural depende de la escala de observación y los criterios de medición, su delimitación precisa presenta una serie de desafíos conceptuales. Esta ambigüedad se puede apreciar en la variedad de términos para describir los límites altitudinales de los árboles. Por ejemplo, la línea de árboles (treeline) se refiere al límite altitudinal o latitudinal donde los árboles crecen formando parches discontinuos; por otro lado, la línea de madera (timberline) marca el borde del bosque cerrado continuo; mientras que la línea de especies arbóreas (tree species line) señala la presencia de los últimos individuos aislados en microambientes favorables (Körner, 2012; Öberg & Kullman, 2012).
La línea de árboles puede servir como un indicador de las condiciones ambientales en los sistemas de montaña, ya que su posición puede cambiar en respuesta a variaciones en la topografía y el clima, repercutiendo en la biodiversidad y otros servicios ecosistémicos (Harsch et al., 2009). En las últimas décadas se han observado cambios en la posición de la línea de árboles en muchas partes del mundo, con un desplazamiento mayoritario hacia altitudes y latitudes más altas (Harsch et al., 2009). Este desplazamiento se ha atribuido principalmente al calentamiento global y otros cambios en el clima que están modificando las condiciones ambientales en los sistemas de montaña volviéndolas más favorables para el crecimiento de los árboles. Sin embargo, los cambios mejor documentados corresponden a las zonas templadas del hemisferio norte, mientras que, en los trópicos, a pesar de su dinámica climática única, persiste un vacío de información. Allí, aunque las temperaturas invernales (asociadas a entradas de aire frío) afectan la línea de árboles, su relevancia ecológica es menor debido a la escasa persistencia de la nieve, un aspecto poco estudiado (Dandan et al., 2022; Hansson et al., 2021; Lauer, 1978; Wieser et al., 2014).
Un número cada vez mayor de estudios ha analizado los cambios de posición y patrones espaciales en la línea de árboles en diferentes escalas geográficas. A escala local, diversos estudios se han centrado en los cambios en la densidad de los árboles, la distribución de las especies y la composición de la zona de transición del límite arbóreo a escala del paisaje (Sigdel et al., 2020). Sin embargo, sigue habiendo una falta de investigación sobre la heterogeneidad espaciotemporal del límite arbóreo a escala de paisaje. La influencia de la topografía en la ubicación y distribución de la línea de árboles se ha observado en muchas montañas en el mundo, ya que condiciona directamente la radiación solar, la velocidad y dirección del viento, generando una diversidad de microclimas. Por ejemplo, las áreas soleadas y convexas tienden a ser más cálidas y propicias para el crecimiento de plántulas, mientras que el exceso de calor y la sequía pueden inhibir el establecimiento de árboles (Holtmeier & Broll, 2012). Por otro lado, la pendiente del terreno y la orografía que brinden refugio contra el viento también pueden influir en el establecimiento y supervivencia de los árboles (Autio, 2006; Autio & Colpaert, 2005; Bansal et al., 2011; Malanson et al., 2007). Lo anterior explica las diferencias en los patrones de la línea de árboles y su dinámica desde la escala local a la global (Bader et al., 2021). Gaire et al. (2014) revelaron que la línea de árboles está ascendiendo a un ritmo de 2,61 m/año en la escala de parcela, mientras que Mohapatra et al. (2019) descubrieron que la línea de árboles está ascendiendo a un ritmo de 11,3 m/año a escala de paisaje en el Himalaya central de Nepal. Sin embargo, un metanálisis de las tasas anuales de cambio de la línea de árboles en el hemisferio norte mostró que la tasa media de cambio fue de 0,354 m/año (Lu et al., 2021).
Recientemente, la percepción remota y los mapas históricos han sido utilizados para evaluar el cambio en la línea de árboles debido a la gran proporción de conjuntos de datos que cubren grandes áreas. El Índice de Vegetación de Diferencia Normalizada (NDVI por sus siglas en inglés) es el índice de vegetación más común y es bien conocido como un buen estimador de la biomasa y la densidad de los árboles. Por lo tanto, el seguimiento de los cambios en el NDVI proporciona una estimación cuantitativa de los cambios a lo largo de la línea de árboles. Sin embargo, se necesita más investigación para validar esta relación, y evaluar el efecto potencial del calentamiento global en los movimientos de la línea de árboles. Al mismo tiempo, debido a que la línea de árboles está íntimamente ligada al mosaico de formas del terreno y sus consecuentes efectos ambientales en las condiciones del sitio, es crucial evaluar el impacto de los patrones topográficos en el desplazamiento de la línea de árboles, ya que este movimiento no puede ser interpretado únicamente a través de modelos de isoterma (Holtmeier & Broll, 2005).
En México, el Eje Volcánico Transversal o Eje Neovolcánico comprende numerosos volcanes donde se ha estudiado la dinámica de la línea de árboles. Por ejemplo, se ha documentado que, a lo largo del último tercio del Holoceno, la línea de árboles se ha desplazado entre 700 y 500 metros debido a fluctuaciones climáticas asociadas a avances y retrocesos glaciales (Lozano-García & Vázquez-Selem, 2005). De manera similar, investigaciones dendrocronológicas sugieren que los árboles han migrado 300 metros hasta su límite actual en los últimos 500 años (Biondi, 2001), así como 70 metros en las últimas cinco décadas (Astudillo-Sánchez et al., 2019). Y registros satelitales (1986-2018) documentan un ascenso promedio de 553 metros (equivalente a 17.3 m/año) en volcanes del centro de México (Jiménez-García et al., 2021). Estos desplazamientos no son homogéneos, su magnitud varía según la topografía local. Las crestas suelen albergar la línea de árboles a mayor elevación, mientras que en depresiones este límite desciende, posiblemente por efectos de drenaje de aire frío o exceso de humedad edáfica (Beaman, 1962). Aunque la tendencia general es ascendente, como muestran los 67 m de avance registrados entre 1955 y 2012 en el estudio de Torres-Beltrán et al. (2017), persisten vacíos en la comprensión de los patrones a escala microtopográfica y su relación con la heterogeneidad ambiental. En este contexto nos planteamos las siguientes preguntas de investigación: ¿Cómo ha cambiado la posición de la línea de árboles en el volcán Iztaccíhuatl, ubicado en el Eje Neovolcánico? y ¿Qué condiciones ambientales han sido más favorables para el avance de la línea de árboles? Nuestras hipótesis fueron: Primero, que la posición de la línea de árboles ha avanzado mayoritariamente hacia mayores altitudes. Segundo, que las variaciones topográficas crean microclimas locales con un efecto significativo en la dinámica de la línea de árboles, por lo que solo ciertas zonas favorecerán el avance de la línea de árboles.
Finalmente, nuestros objetivos fueron los siguientes: 1) Comparar los cambios en la posición de la línea de árboles en un sistema montañoso en el Volcán Iztaccíhuatl durante los periodos 1985-2000, 2000-2014 y 2014-2020 utilizando imágenes satelitales. 2) Analizar la influencia de la topografía en la distribución de la línea de árboles, identificando las características del terreno que favorecen o inhiben el crecimiento y establecimiento de los árboles. Para abordar estos objetivos, utilizamos Sistemas de Información Geográfica con imágenes satelitales Landsat y un modelo de elevación digital. Evaluamos los cambios en el tipo de vegetación y clasificamos diferentes ambientes según variables del terreno. Nuestro estudio integra el papel del ambiente local con la capacidad de respuesta de los árboles para el avance o el retroceso de la cobertura forestal en la dinámica de la línea de árboles en el volcán Iztaccíhuatl.
Metodología
Zona de estudio
La zona de estudio se encuentra en el Eje Volcánico Transversal ubicado en el centro de México. Esta cadena montañosa engloba las cimas más elevadas del país, entre las cuales destaca el volcán Iztaccíhuatl (19.172°N -98.641°O), el tercer volcán más alto, con una altura de 5,284 metros sobre el nivel del mar. El volcán Iztaccíhuatl ha permanecido inactivo desde finales del Pleistoceno, lo que ha permitido que la migración de los bosques esté regulada principalmente por factores climáticos y humanos, más que por la actividad volcánica. Entre los factores antropogénicos destacan los incendios provocados para renovar pastos ganaderos y el pisoteo generado por las veredas de uso turístico (Lenero et al., 2007). El clima varía con la altitud, a 3,700 msnm la temperatura media anual es de 5.5°C, descendiendo a 5°C a 4,000 msnm, se ha reportado un gradiente térmico de -0.68 °C por cada 100 metros (Jasso-Flores et al., 2019). La precipitación anual acumulada oscila entre 874 mm (3,700 m) y 588 mm (4,000 m), concentrándose principalmente durante los meses de junio a octubre (CONANP, 2013).
La vegetación está compuesta principalmente por bosques de coníferas, con Pinus hartwegii como la especie que establece la línea de árboles. En las cotas inferiores de su distribución (3,400-3,700 m), P. hartwegii forma asociaciones con P. montezumae, donde el dosel semicerrado (15-20 m de altura) permite el desarrollo de un sotobosque diverso con géneros como Senecio, Ribes, Muhlenbergia, Agrostis, Lupinus y Festuca. A medida que se asciende (3,700-4,000 m), P. hartwegii se vuelve la especie arbórea exclusiva, con una conformación más abierta donde coexiste con zacatonales densos y leguminosas del género Lupinus (Giménez de Azcárate Cornide et al., 2009). En su límite altitudinal superior (4,000 - 4,200 m), la especie adopta una forma achaparrada (5-8 m de altura), con individuos aislados, entre los zacatonales, que resisten temperaturas bajo cero, nevadas recurrentes e incluso incendios superficiales (CONANP, 2013).
Entre los 4,000 y los 4,500 m se establece el pastizal alpino donde dominan las gramíneas amacolladas. Se pueden distinguir dos grupos de comunidades, una que se desarrolla en los llanos sobre sitios bajos y mal drenados, dedicados históricamente al pastoreo de ganado (Giménez de Azcárate Cornide et al., 2009). Y la comunidad del pastizal clímax, esta comunidad frecuentemente es resultado de la substitución de la vegetación original o de la destrucción del bosque de pino previa a su establecimiento. La composición del pastizal alpino puede ser muy homogénea pero sus especies dominantes varían según la altitud, formando cinturones definidos (Giménez de Azcárate Cornide et al., 2009). Entre el pastizal, resalta la presencia de arbustos enanos como Juniperus montícola en microambientes en los que permanecen protegidos de los incendios forestales (CONANP, 2013).
Selección y procesamiento de imágenes satelitales
Para comparar los cambios en la posición de la línea de árboles utilizamos imágenes satelitales Landsat seleccionadas en función de la visibilidad de la línea de árboles (Solár & Janiga, 2013; Zinnert et al., 2011). Obtuvimos imágenes Landsat con una resolución espacial media de 30 m de la ESA (European Space Agency), y realizamos procesos de clasificación supervisada que nos permitieran identificar la línea de árboles. También obtuvimos distintas variables predictivas que utilizamos en nuestros análisis, tales como el índice normalizado de diferencia de la vegetación (NDVI por sus siglas en inglés), la temperatura de la superficie terrestre, la radiación recibida, la radiación emitida y el índice de humedad del suelo (SMI, por sus siglas en inglés). Debido a la nubosidad, principalmente durante la temporada lluviosa, no logramos contar con una muestra representativa de los cambios estacionales, por lo que analizamos cambios anuales basándonos en la visibilidad de la línea de árboles y elegimos las imágenes de la época seca (diciembre- mayo) de los años 1985, 2000, 2014 y 2020. Para los primeros dos años usamos imágenes Landsat 5 TM Collection 2 con corrección atmosférica de reflectancia superficial Tier 1y Landsat 8 OLI/TIRS Collection 2 para las otras dos fechas. Para cada fecha generamos imágenes compuestas en las que eliminamos los píxeles contaminados por nubes y sombras de nubes en las imágenes.
Sobre las imágenes compuestas realizamos un proceso de clasificación supervisada utilizando un clasificador de bosques aleatorios y una selección aleatoria de puntos sobre mapas de tipos de vegetación de México según INEGI (1997, 2001, 2013, 2021) como datos de entrenamiento para el modelo. Las categorías consideradas fueron cobertura forestal, cobertura de pastizal y suelo desnudo, el glaciar se trató como pixeles sin datos. Posteriormente, destinamos el 70% de los datos al conjunto de entrenamiento y el 30% restante al conjunto de validación para evaluar la precisión del clasificador. Con los conjuntos de entrenamiento y validación definidos, entrenamos un clasificador con 10 árboles en el bosque aleatorio. La clasificación resultante se guardó como una imagen raster y se utilizó para evaluar cambios en la línea de árboles.
El NDVI ha sido ampliamente utilizado para la detección de cambio de vegetación (verdor y estado de la vegetación) (Myneni et al., 1998), que puede ayudan a distinguir la densidad de la vegetación y su dinámica (Singh et al., 2015). Por lo tanto, calculamos el NDVI de las imágenes compuestas de 1985 y 2020 restando el valor de la reflectancia en el infrarrojo cercano (NIR) del valor de la reflectancia en el rojo (RED), y luego dividiendo esta diferencia por la suma de ambos valores. Guardamos las imágenes resultantes como imágenes raster. Adicionalmente generamos imágenes de la temperatura de la superficie terrestre, la radiación recibida y la radiación emitida para cada uno de los años seleccionados. Estas variables se obtuvieron a partir de las bandas espectrales correspondientes a cada satélite. Para Landsat 5 TM, la temperatura de la superficie terrestre se derivó de la banda térmica ST_B6, aplicando un factor de escala de 0.00341802 para convertir los valores digitales a grados Celsius. En el caso de Landsat 8 TIRS, se utilizó la banda ST_B10 con el mismo factor de escala, asegurando consistencia en las unidades entre ambos sensores. Esta conversión permitió estimar la LST en grados Celsius, facilitando el análisis de las variaciones térmicas en el área de estudio a lo largo del tiempo. La radiación recibida y emitida se extrajeron de las bandas ST_DRAD y ST_URAD, respectivamente, tanto para Landsat 5 como para Landsat 8. Los valores de radiación se ajustaron multiplicándolos por 0.001, convirtiéndolos así a unidades estándar (W·m⁻²·sr⁻¹·µm⁻¹). Estas unidades representan la energía radiante por unidad de área, considerando la dirección de medición y el rango espectral del sensor, lo que garantizó la comparabilidad temporal de los datos. El SMI se calculó dividiendo el NDVI entre la radiación emitida para cada conjunto de datos correspondiente a los años 1985, 2000, 2014 y 2020. Los resultados de la extracción fueron verificados y revisados con Imágenes de alta resolución proporcionadas por la plataforma Google Earth. Además de las imágenes Landsat, utilizamos datos de un modelo de elevación digital (DEM). Procesamos datos del DEM proporcionado por el Shuttle Radar Topography Mission (SRTM). Realizamos la extracción de variables topográficas como pendiente, aspecto (dirección de la pendiente) y hillshade (o forma de ladera, destaca la presencia de colinas y montañas) de la región de interés definida mediante coordenadas que delimitan el área alrededor del Iztaccíhuatl. Calculamos las variables de pendiente, aspecto y forma ladera utilizando las funciones disponibles en la biblioteca ee.Terrain de Google Earth Engine.
Detección de cambio de la línea de árboles
Para calcular cuantitativamente los cambios en la cobertura vegetal entre los años 1985, 2000, 2014 y 2020 utilizamos los datos de NDVI de la elevación máxima de la cobertura forestal y la elevación mínima de la cobertura de pastizal como un indicador de la posición de la línea de árboles. Utilizamos las imágenes raster de los tipos de cobertura resultantes de nuestro proceso de clasificación supervisada para obtener la diferencia absoluta entre pares de años consecutivos (1985-2000, 2000-2014, y 2014-2020), al restar los valores de cada píxel de un año al valor del mismo píxel en el año siguiente. La diferencia absoluta nos permitió identificar cambios en la reflectancia que indicaran una variación en la cobertura vegetal. Con las diferencias absolutas creamos mapas binarios de cambio. En estos mapas, los píxeles con un valor diferente de cero se marcaron como cambio y los píxeles con un valor de cero se marcaron como no cambio. Posteriormente calculamos el porcentaje de píxeles que cambiaron, dividiendo el número de píxeles marcados como cambio por el número total de píxeles en la imagen, y multiplicando el resultado por 100. Por último, para analizar las dinámicas específicas de los cambios en la cobertura vegetal, realizamos matrices de transición para cada periodo, en las que se mostrara cómo las categorías de interés (cobertura forestal y cobertura de pastizal) se transformaron de un año a otro. Además de las matrices de transición, calculamos las proporciones de transición para obtener una perspectiva porcentual de los cambios. Esto implicó dividir la cantidad de píxeles que cambiaron de una categoría a otra por el total de píxeles en la categoría inicial, multiplicando el resultado por 100. Las proporciones de transición proporcionaron información detallada sobre la magnitud y dirección de los cambios en la cobertura vegetal.
También utilizamos un enfoque cuantitativo que nos permitió evaluar el cambio altitudinal de la línea de árboles en pixeles específicos. Usamos las imágenes raster de cobertura vegetal y la herramienta "Poligonizar" en QGIS para convertir los raster a polígonos, y así obtener una representación vectorial del límite entre la cobertura forestal y la cobertura de pastizal. Con los polígonos generados empleamos la herramienta "Extraer vértices" en QGIS para identificar los vértices de la línea de árboles. Para evaluar los cambios en la altitud de los de la línea de árboles, definimos un total de 155 cuadros de 100 x 100 metros distribuidos a lo largo de la línea de árboles del Volcán. Utilizamos el modelo de elevación digital (DEM) de la NASA SRTM con una resolución de 30 metros para obtener la elevación de cada vértice dentro de los cuadros definidos. Con los datos de elevación de los vértices, realizamos modelos de regresión lineal por cada uno de los 155 cuadros. Para cada cuadro, ajustamos una regresión lineal que relacionaba la altitud de los vértices con el tiempo según el año de cada imagen (1985, 2000, 2014 y 2020), y analizamos la pendiente de cada modelo de regresión lineal. La pendiente de la regresión nos indicó la dirección y magnitud del cambio en la altitud de la línea de árboles en cada cuadro: Una pendiente positiva indicaba un avance de la línea de árboles (aumento en altitud), y una pendiente negativa un retroceso de la línea de árboles (disminución en altitud), y una pendiente cercana a cero señalaba un mantenimiento de la posición de la línea de árboles.
Caracterización de los micrositios de colonización
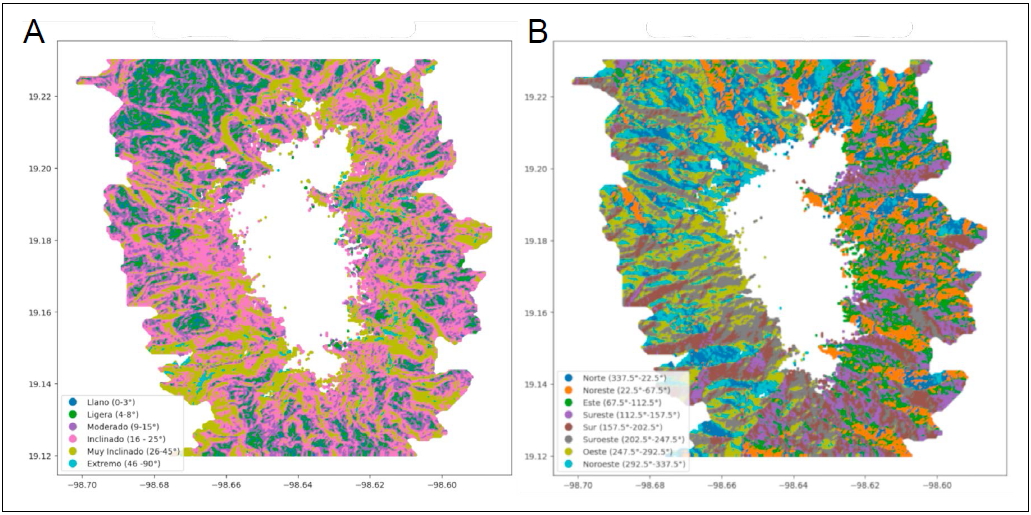
Para analizar la influencia de la topografía en la línea de árboles nos enfocamos en identificar los principales micrositios que favorecen o inhiben el crecimiento y establecimiento de los árboles. Consideramos a la pendiente y la orientación como variables clave, por su papel fundamental en la regulación hídrica y térmica, factores limitantes clave para el establecimiento de árboles. Clasificamos la pendiente en seis categorías: Llano (0-3°), Ligera (4-8°), Moderada (9-15°), Inclinada (16-25°), Muy Inclinada (26-45°) y Extrema (46-90°) (Fig. 1A). La orientación se dividió en ocho categorías: Norte (337.5°-22.5°), Noreste (22.5°-67.5°), Este (67.5°-112.5°), Sureste (112.5°-157.5°), Sur (157.5°-202.5°), Suroeste (202.5°-247.5°), Oeste (247.5°-292.5°) y Noroeste (292.5°-337.5°) (Fig. 5B). Combinamos estas dos clasificaciones para obtener 48 categorías topográficas con el objetivo de evaluar cómo la pendiente y la orientación, al influir en el microclima, afectan el establecimiento de árboles. Para cada una de estas categorías, calculamos los estadísticos descriptivos de las variables de temperatura, índice de humedad (SMI), radiación descendente (DownRad), radiación ascendente (UpRad) y NDVI, luego estandarizamos los datos utilizando la función StandardScaler de Scikit-Learn. Aplicamos un análisis de agrupamiento jerárquico con la función linkage del módulo scipy cluster hierarchy, empleando el método de Ward para generar un dendrograma y observar el agrupamiento de las categorías con base en los cambios de cobertura (de cobertura forestal a cobertura de pastizal o viceversa). Establecimos una distancia máxima de 8.5 para definir los grupos resultantes. Finalmente, comparamos las agrupaciones mediante el cálculo de estadísticos descriptivos (media, desviación estándar, valor mínimo y máximo) de las variables climáticas y de vegetación.
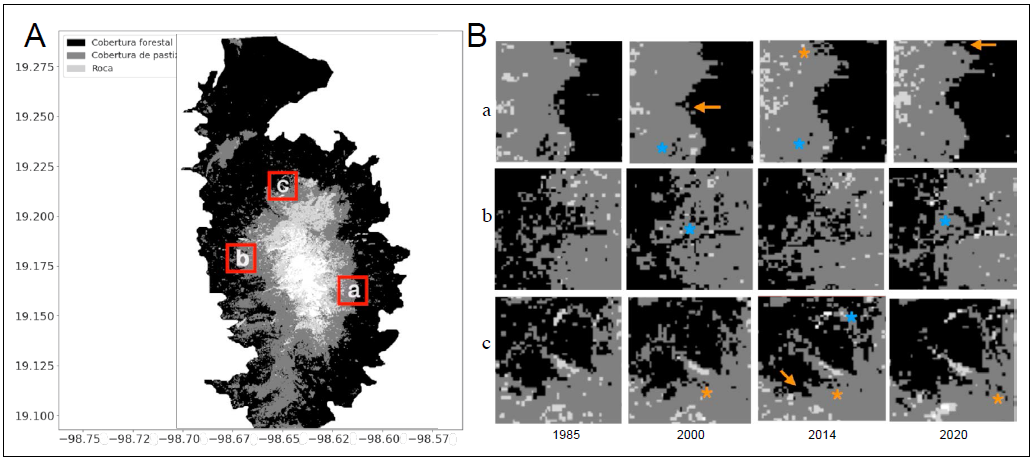
Figura 1 Clasificación de la cobertura vegetal y mapas de cambio. A) Ejemplo de clasificación supervisada según los tipos de vegetación para el 2020. Se observan tres categorías de cobertura: forestal, pastizal y roca. B) Detalles de cambio en tres cuadros, se observan cambios en la línea de árboles de 1985 a 2020, tales como aumento de la densidad (asterisco naranja) o la disminución de la densidad (asterisco azul) y el avance de la línea de árboles (flecha naranja).
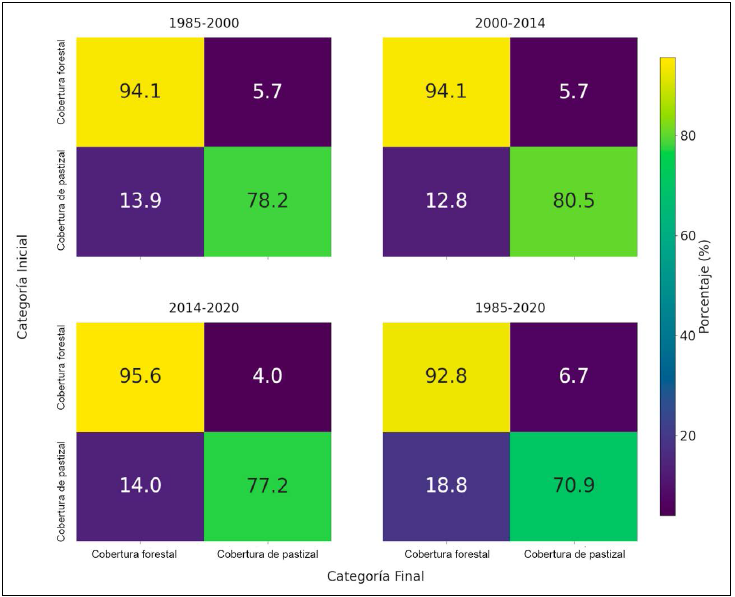
Figura 2 Matrices de transición de cobertura vegetal. Se observan las matrices de transición para los periodos analizados: 1985 a 2000, 2000 a 2014, 2014 a 2020 y 1985 a 2020. Cada matriz muestra el porcentaje de píxeles que cambiaron de categoría (cobertura forestal o pastizal) entre los años comparados. Los valores representan las transiciones entre las dos categorías disponibles, donde la escala de color indica la intensidad de las transiciones.
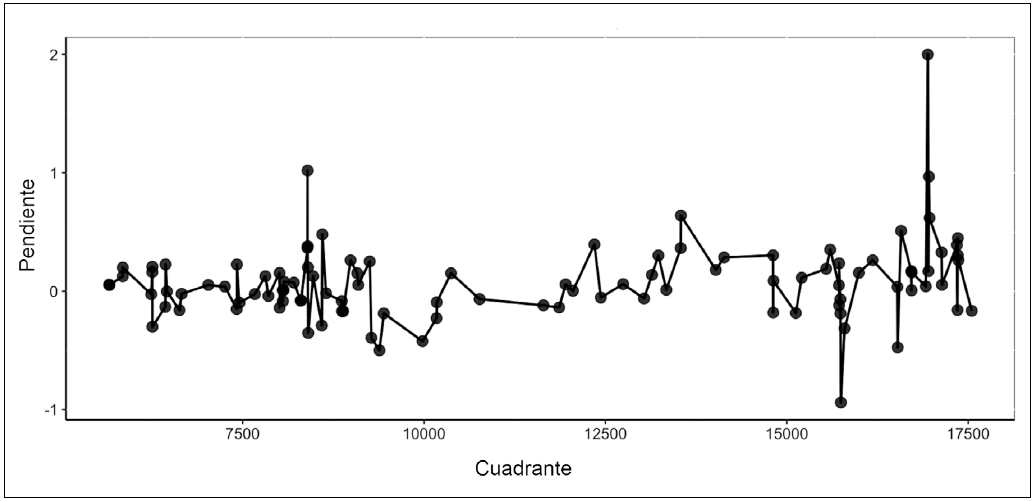
Figura 3 Dinámica altitudinal de la línea de árboles. Los valores de pendiente del modelo linela por cuadrante reflejan la dirección y magnitud del desplazamiento: pendientes cercanas a cero indican estabilidad, valores positivos muestran ascenso hacia mayores elevaciones, y valores negativos revelan descenso hacia cotas inferiores.
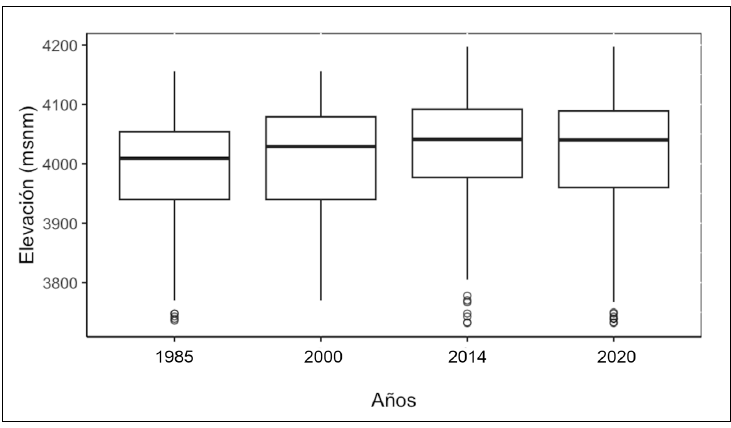
Figura 4 Distribución altitudinal de la línea de árboles (1984-2020). Los boxplots muestran media (línea central), rango intercuartílico (cajas) y valores extremos (puntos) para cada periodo evaluado.
Resultados
Detección de cambio en la línea de árboles
El proceso de clasificación supervisada (coeficiente Kappa de 0.6977) permitió identificar la línea de árboles como el límite entre la cobertura forestal y la cobertura de pastizal, así como cuantificar las transiciones entre los dos tipos de cobertura a lo largo del tiempo. La línea de árboles se presenta de forma irregular, con altitud variable y límites difusos que constituyen una zona de transición con numerosas ‘islas de árboles’, es decir, píxeles de cobertura forestal aislados. Los cambios en la distribución de los pixeles de cobertura forestal sugieren incrementos o disminuciones en la densidad de las poblaciones de P. hartwegii (Fig. 1B). Los píxeles de cobertura forestal que superan ampliamente la zona de transición se consideraron mal clasificados por el modelo, ya que no coincidían con la la línea de árboles que se establece a los 4000 metros (Fig. 1A).
El análisis de transiciones entre cobertura forestal y de pastizal reveló que entre 1985-2000, el 94.08% de la cobertura forestal se mantuvo estable, mientras que un 5.7% se transformó en pastizal. En el mismo periodo, el 78.17% de la cobertura de pastizal permaneció sin cambios, y un 13.94% fue reemplazado por cobertura forestal. En el periodo de 2000 a 2014, la persistencia de la cobertura forestal fue similar (94.11%), con una ligera reducción en su conversión a pastizal (5.65%). La cobertura de pastizal mostró mayor estabilidad (80.45%), aunque con menor avance de la cobertura forestal (12.83%). El periodo 2014-2020 registró la mayor estabilidad forestal (95.64%) y la menor pérdida (4.02%). Los pastizales, en cambio, presentaron mayor dinámica, solo 77.2% se mantuvo estable, mientras que 13.97% cambió a cobertura forestal.
A escala del estudio completo (1985-2020), la cobertura forestal mostró alta persistencia (92.84%), pero con una pérdida acumulada del 6.74%, lo que podría estar relacionado con la apertura de claros en el bosque. Destaca la expansión forestal sobre pastizales (18.83%), que redujo su cobertura estable a 70.87% (Fig. 2).
Observamos tanto avances como retrocesos en la línea de árboles, con predominio del aumento en altitud. La magnitud del avance de la línea de árboles fue generalmente pequeña. De los 155 cuadros evaluados, el 60% presentaron pendientes positivas en sus respectivos modelos de regresión lineal, lo que sugiere un avance de la línea de árboles durante todo el periodo. De este 60%, cerca de la mitad presentaron pendientes menores a 0.1, lo que indica que el aumento altitudinal fue sutil. Solo en tres casos se observaron pendientes cercanas o superiores a 1, lo que refleja un avance más significativo de la línea de árboles en esos cuadrantes. En el 40% de los cuadros con pendientes negativas, ninguna pendiente alcanzó un valor de -1, sugiriendo que los retrocesos altitudinales también fueron sutiles (Fig. 3).
La altitud promedio de los cuadros utilizados para la detección del avance de la línea de árboles presentó un incremento desde 1985 hasta 2014, seguido de una estabilización en el último periodo. Entre 1985 y 2000 se registró un aumento en la media de la altitud límite de los árboles de 20 m, mientras que entre 2000 y 2014 el incremento fue de 12 metros, y para el año 2020 se observó una disminución de 1 metro. También se identificó un aumento en la altitud mínima durante el primer periodo; entre 1985 y 2000 el valor mínimo se incrementó en 34 metros, aunque disminuyó en 38 metros hacia 2014, manteniéndose sin cambios en 2020. Los valores de altitud máxima llegaron a los 4200 metros, lo cual no coincide con las observaciones de campo en los que establecen el límite del bosque continuo (timberline) a los 4000 metros (Fig. 4). Esta discrepancia podría deberse a que los datos satelitales están captando parches discontinuos de árboles (treeline) en altitudes superiores, los cuales también pueden observarse actualmente en campo incluso hasta los 4300 metros.
Caracterización de los micrositos de colonización
En el área de estudio predominan las pendientes inclinadas, seguidas de las moderadas y muy inclinadas. En contraste, se observa una menor cantidad de pixeles clasificados como llanos o de pendientes extremas (Fig. 5A). En cuanto a los pixeles de transición, encontramos que cambios de cobertura de bosque a cobertura de pastizal y de cobertura de pastizal a cobertura de bosque) pueden ocurrir dentro de la misma categoría, aunque también se observó una tendencia hacia un tipo de cambio específico (Fig. 6). Por ejemplo, en pendientes extremas con orientación oeste, categoría con el menor número de pixeles, el 100% de los cambios correspondió a la transición de cobertura de bosque a cobertura de pastizal (Fig. 6).
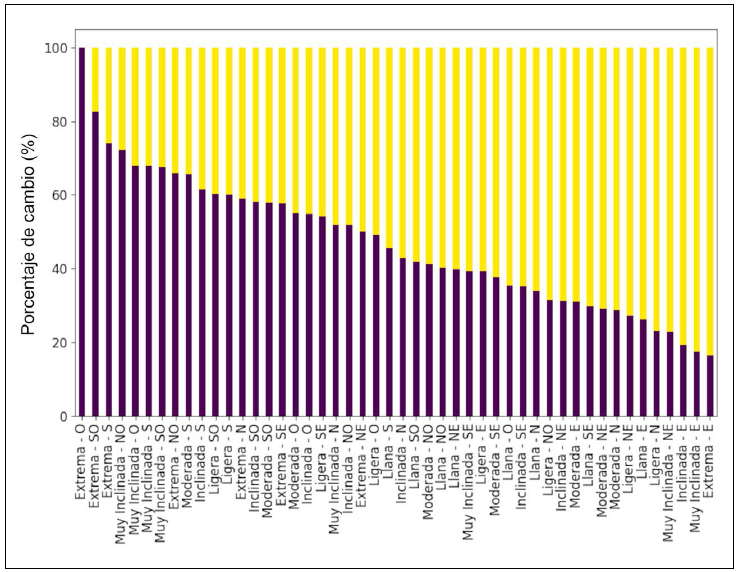
Figura 6 Porcentaje de cambio entre cobertura forestal y cobertura de pastizal (1985-2020) distribuido según las 48 categorías topográficas combinadas (pendiente-orientación). El análisis distingue dos dinámicas principales: la transición de cobertura forestal a cobertura de pastizal (representada en azul, ordenada de mayor a menor porcentaje) y su proceso inverso de cobertura de pastizal a cobertura forestal (en amarillo, ordenado de menor a mayor). Los valores porcentuales reflejan la proporción relativa de píxeles que experimentaron cada transición, donde el 100% equivale al total de los pixeles modificados, excluyendo los que mantuvieron su cobertura original.
Las 48 categorías topográficas se agruparon en siete conjuntos con base en sus similitudes climáticas y de vegetación. Los grupos 1, 2 y 3 están compuestos mayormente por sitios con pendientes extremas a muy inclinadas, con orientaciones noroeste, oeste y suroeste, en estos grupos predominó el cambio de cobertura forestal a cobertura de pastizal. El grupo 4 se caracterizó por pendientes extremas y con orientaciones noreste y este, donde el cambio predominante fue de cobertura de pastizal a cobertura forestal.
El grupo 5 mostró mayor heterogeneidad, incluyendo categorías de pendiente ligera, moderada, inclinada, muy inclinada y extrema, con orientaciones sur, este y sureste; en este grupo se presentaron ambos tipos de cambio sin una tendencia clara hacia uno en particular. El grupo 6 se compone de pendientes bajas, clasificadas como ligeras o llanas, con predominancia del cambio de cobertura de pastizal a cobertura forestal. Finalmente, el grupo 7 incluye pendientes ligeras, moderadas e inclinadas, principalmente con orientación norte, y en él también predominó el cambio de cobertura pastizal a cobertura forestal (Fig. 7).
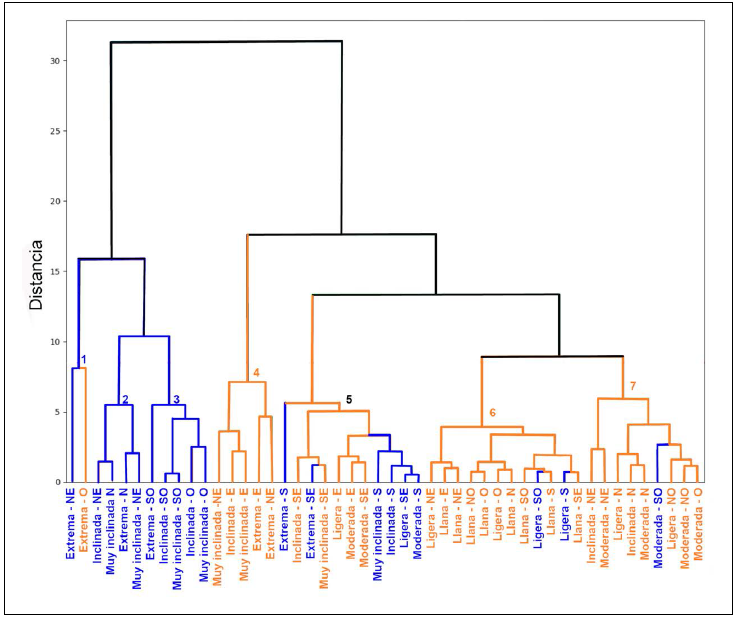
Figura 7 Dendrograma de las 48 categorías combinadas de pendiente y orientación, agrupadas según variables ambientales (Temperatura, SMI, Radiación y NDVI). El color de las categorías destaca el tipo de cambio predominante: en azul, transición de cobertura forestal a pastizal; en naranja, de pastizal a forestal. Cada grupo está marcado con un número (generado automáticamente por el análisis), cuyo color refleja la tendencia principal de ese cluster. En casos como el grupo 5 (negro), el color neutro indica ausencia de una dirección de cambio clara. Las líneas del dendrograma también siguen el patrón de color.
Los menores valores de temperatura del aire y del índice de humedad del suelo (SMI) se observaron en los grupos 1 y 2, coincidiendo con la predominancia del cambio de cobertura forestal a cobertura de pastizal. El grupo 5 presentó los valores más altos de temperatura, donde no se identificó una tendencia clara en el tipo de cambio. Aunque el grupo 5 presentó los valores atípicos más altos de SMI, el valor promedio más elevado se encontró en el grupo 4. Entre los grupos con predominancia de cobertura de pastizal a cobertura forestal, el grupo 4 presentó la temperatura media más alta (18.54 °C) entre los tres, mientras que los grupos 6 y 7 presentan medias de 17.42 °C y 16.01 °C, respectivamente. Por otro lado, los grupos 6 y 7 comparten una mayor homogeneidad en sus variaciones, con desviaciones estándar relativamente bajas en ambos parámetros, lo que sugiere condiciones más estables (Fig. 8)
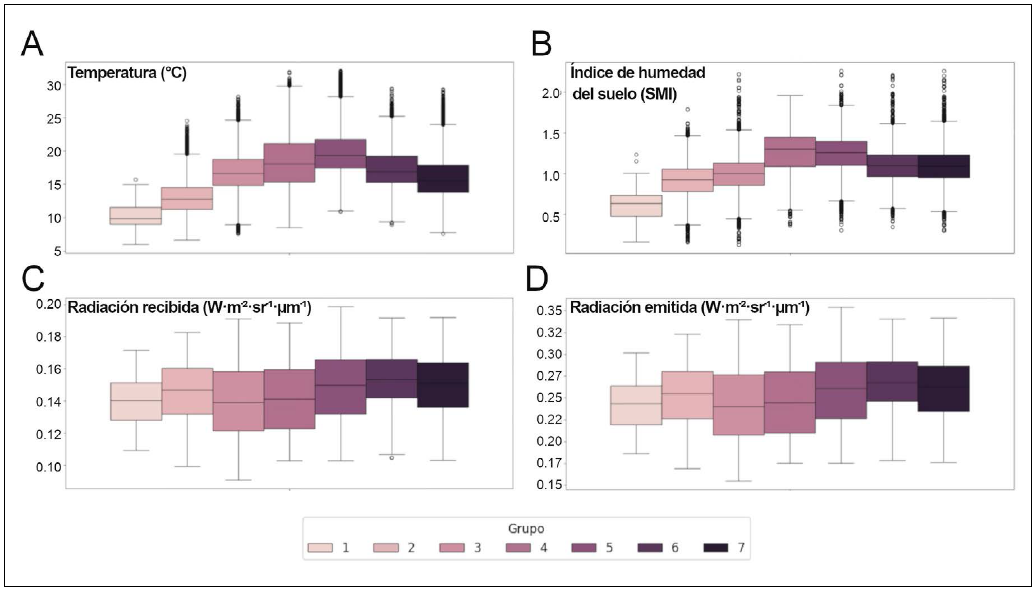
Figura 8 Distribución por agrupaciones de (A) temperatura superficial (°C), (B) índice de humedad del suelo (SMI), (C) radiación solar recibida (W·m⁻²·sr⁻¹·µm⁻¹), y (D) radiación emitida (W·m⁻²·sr⁻¹·µm⁻¹). Cada diagrama de caja muestra la media (línea central), rango intercuartílico (caja), valores extremos (bigotes) y outliers (puntos). Los colores distinguen las siete agrupaciones según se indica en la leyenda.
Discusión
Cambios en la línea de árboles
La topografía y el clima explican el avance de la línea de árboles, el clima (temperatura del aire y los patrones de precipitación) es el factor principal que limita el crecimiento de los árboles, y la topografía (temperatura y humedad del suelo) influye en la dirección y distribución del límite de dicha línea. Los análisis de las imágenes de satélite de la distribución altitudinal de la cobertura forestal y la cobertura de pastizal combinados con variables ambientales permiten entender la distribución de la cobertura vegetal en gradientes altitudinales (Mu et al., 2022). Nuestros resultados confirmaron la hipótesis de que la posición de la línea de árboles en el Volcán Iztaccíhuatl ha avanzado hacia altitudes mayores entre 20 y 38 m, lo que es consistente con estudios previos que reportan aumentos de 20 a 65 m en la línea de árboles en otros sistemas montañosos del Eje Neovolcánico (Alfaro-Ramírez et al., 2017). Aunque observamos una tendencia general de avance hacia altitudes superiores, los incrementos no fueron ni uniformes ni continuos en las fechas evaluadas, sugiriendo el avance de la línea de árboles solo se presenta en condiciones microambientales específicas y depende de las condiciones metereológicas anuales. Además, se identifican píxeles aislados formando islas distantes de la masa cobertura forestal indicando que hay retrocesos causados por factores como la mortalidad. Los avances y retrocesos de la línea de árboles sugieren que la migración de individuos es por saltos y difuso. Alfaro-Ramírez et al. (2017) clasificaron la línea de árboles de P. hartwegii en el Nevado de Toluca como difusa siguiendo lo propuesta por Harsch & Bader (2011), lo cual concuerda mayormente con nuestras observaciones en el Iztaccíhuatl, donde se observa una disminución gradual en la densidad de los píxeles. Investigaciones sobre la conectividad de los fragmentos de bosque de P. hartwegii reportan distancias entre fragmentos de 19.1 a 52.7 metros (García-Amorena et al., 2021), esto reduce el flujo de dispersión y la fuerza de las conexiones con otros parches, favoreciendo el aislamiento de parches. Este fenómeno podría ser objeto de estudios más detallados con una mayor cantidad de imágenes anuales.
Otro factor importante para considerar en cuanto al avance de la línea de árboles son los incendios forestales. En el Parque Nacional Izta-Popo, la alta frecuencia de incendios ha favorecido la expansión de pastizales y la colonización de especies exógenas, desplazando a la vegetación original, incluidos los árboles del bosque (Gutiérrez Martínez et al., 2015). Además, los incendios recurrentes, especialmente en ecosistemas templados como los bosques de P. hartwegii, incrementan la mortalidad de individuos juveniles, lo que impide el reclutamiento natural y acelera la degradación del bosque. Estudios como el de Robles-Gutiérrez et al. (2016) demuestran que, tras un incendio de mediana a alta intensidad, la mortalidad de los pinos jóvenes puede alcanzar hasta un 28%, en comparación con solo un 4% en áreas no quemadas. Estos factores, combinados con la alteración del régimen natural del fuego, generan cambios profundos en la estructura y composición del bosque, favoreciendo la transición de bosques a pastizales.
A nivel global, el desplazamiento de la línea de árboles hacia elevaciones más altas se ha asociado con el calentamiento global, especialmente el aumento en la temperatura del aire propicia que las isotermas (líneas de igual temperatura) se desplacen a mayor altitud, creando una «ventana» para que los árboles amplíen su área de distribución, en la que los árboles pueden sobrevivir, ya que la temperatura suele ser un factor limitante para su crecimiento (Monleon & Lintz, 2015). En el Eje Neovolcánico, se ha reportado un aumento de la temperatura máxima de 2 °C desde 1955, con un incremento adicional de 3 °C (de 23 a 26 °C) desde finales de la década de 1970 hasta 2009, coincidiendo con un período de mayor reclutamiento de P. hartwegii (Astudillo-Sánchez et al., 2017). Los cambios temporales en el reclutamiento de árboles sugieren que el avance de la línea de árboles es por pulsos. El mayor avance promedio de la línea de árboles se registró entre 1985 y 2000, mientras que de 2014 a 2020, la línea de árboles retrocedió en promedio 1 m. Según Astudillo-Sánchez et al. (2017), los fenómenos del sistema climático como los eventos de El Niño y La Niña influyen en el reclutamiento de nuevos árboles. Durante los eventos de El Niño, caracterizado por un calentamiento anormal superior a 0.5°C en el Pacífico Tropical, el aumento de lluvias invernales (entre diciembre y marzo) combinado con veranos más secos crean condiciones ideales para el establecimiento de nuevos árboles, al reducir las heladas y proporcionar humedad crítica en etapas tempranas. Por el contrario, los episodios de La Niña, consecuencia de un enfriamiento de al menos -0.5°C en el Pacífico Tropical, traen consigo inviernos particularmente fríos y secos que generalmente dificultan la supervivencia de las plántulas. Esta relación se refleja claramente en los datos de reclutamiento: mientras que en el periodo 1947-1979, dominado por condiciones frías, apenas el 7% de los individuos lograron establecerse, entre 1980 y 2012, cuando los eventos de El Niño fueron más frecuentes e intensos, se registró el 93% del reclutamiento total, con una tasa promedio anual del 5%. Este aumento coincide parcialmente con el periodo de mayor avance en la línea de árboles que nosotros observamos. Estos patrones subrayan la importancia de la estacionalidad y los eventos de lluvia en el reclutamiento de nuevos árboles, lo que influye en los cambios en la línea de árboles. Sin embargo, la observación tanto de avances como retrocesos en la línea de árboles en nuestros resultados sugiere que factores adicionales al aumento de temperatura podrían estar influyendo en estos cambios. Los fenómenos de variación local para la posición de la línea de árboles y el desplazamiento ascendente indica que la temperatura no es el principal parámetro ambiental que promueve avance hacia arriba de la línea de árboles (Xu et al., 2020). Esto subraya la importancia de considerar elementos como la topografía y las interacciones ecológicas específicas de cada micrositio, para una explicación más completa de la dinámica de la línea de árboles.
Micrositos de colonización
Las irregularidades en el terreno (crestas y depresiones) crean microclimas locales que afectan de manera desigual el crecimiento de los árboles, lo que lleva a que la línea de árboles se presente a diferentes altitudes dentro de una misma zona (Beaman, 1962). La interacción entre microclimas locales y la capacidad de respuesta de las plantas resulta fundamental para comprender el avance y retroceso del bosque, especialmente en áreas montañosas. En otros estudios se han encontrado diferencias significativas en la altitud del ecotono entre exposiciones noroeste y este-sureste, donde la cara norte ha registrado la mayor variación altitudinal (Alfaro-Ramírez et al., 2017). En particular, la orientación de las pendientes juega un papel crucial en la dinámica de la línea de árboles. Al agrupar las 48 categorías topográficas se revelaron patrones interesantes en la dinámica del cambio de vegetación. El Grupo 4, con pendientes extremas a inclinadas orientadas hacia el noreste y el este, se caracterizó principalmente por el cambio de pastizal a bosque, mientras que los grupos con pendientes extremas y orientaciones noroeste, oeste y suroeste (Grupos 1, 2 y 3) mostraron una predominancia del cambio de bosque a pastizal. Un resultado similar se observó en el Nevado de Toluca, donde se reportó una mayor abundancia de reclutamiento en exposiciones norte y oeste, contrastando con las exposiciones sur y este caracterizadas por un menor reclutamiento (Astudillo-Sánchez et al., 2019). De manera similar, Torres-Beltrán et al. (2017) documentaron que la orientación E favorece particularmente el ascenso de individuos arbóreos. Se ha comprobado que la orientación de la pendiente influye notablemente en el rango de temperatura de cada micrositio según su altitud. En el Cerro Jocotitlán del Estado de México se han reportado diferencias notables en las temperaturas máximas entre sitios, especialmente en los sitios con orientación SO los cuales suelen mostrar menor oscilación térmica y menos eventos cercanos a 0 °C comparados con otras exposiciones (Núñez García, 2019). Estos hallazgos sugieren que la creación de micrositios es fundamental.
En el Iztaccíhuatl se ha medido que la temperatura anual media es de 1 a 4 °C más alta en las pendientes S en comparación con las N a la misma elevación (López López et al., 2023). En términos generales, nuestros resultados parecen indicar que los máximos de temperatura no favorecen el avance de la línea de árboles, ya que en las orientaciones S se observó principalmente el cambio de bosque a pastizal. Se ha sugerido que la exposición directa al sol (sur y este) suelen representar mayor riesgo de deshidratación y daño térmico, lo que afecta negativamente la supervivencia de plántulas (Astudillo-Sánchez et al., 2019). No obstante, es importante considerar que todos los micrositios presentaron los dos tipos de cambio y que inclusive hubo grupos, como el Grupo 5, con una gran heterogeneidad de pendientes y orientaciones, no mostraron una tendencia clara hacia un tipo de cambio particular. Si bien los máximos de temperatura pueden aumentar la mortalidad de la plántula, una mayor temperatura, especialmente en otoño, es crucial para la actividad del cambium y la xilogénesis (Núñez García, 2019), proceso esencial para el crecimiento y posterior establecimiento de los juveniles. En el Iztaccíhuatl también se han medido las variaciones estacionales de temperatura. En general, las pendientes sur tienen temperaturas anuales promedio 1 a 4 °C más altas en comparación con las pendientes norte a la misma altitud. (López López et al., 2023) demostró que existe un gradiente térmico de 0.55°C /100 m de altitud para el suelo de umbría, y de 0.31°C /100 m de altitud para el de solana, sugiriendo que la exposición en la umbría tiene un control significativo en la marcha de la temperatura del suelo. Sin embargo, durante junio, julio y agosto, las diferencias de temperatura entre pendientes sur y norte son menores y, en algunas altitudes, las temperaturas son incluso ligeramente más altas en las pendientes norte debido a la mayor insolación veraniega (López López et al., 2023). Este tipo de fenómenos podría explicar la predominancia del avance de la línea de árboles en los Grupos 4, 6 y 7. Elliott & and Cowell, (2015) demostraron que la regeneración arbórea de la parte superior del límite arbóreo se explica debido a los regímenes térmicos, sin embargo, aunque las diferencias de temperatura fueron insignificantes entre las laderas, el avance de la línea de árboles varia a lo largo de una misma montaña, como ocurre en nuestro sitio de estudio.
La pendiente es otro factor relevante que también ha mostrado tener un efecto en el avance de la línea de árboles debido a que influye en el microclima, la humedad del suelo y la exposición al viento, que as u vez pueden afectar el establecimiento y crecimiento de los árboles (Zheng et al., 2021). Estudios anteriores han sugerido que las pendientes moderadas (15°-25°) son óptimas para el crecimiento de P. hartwegii, mientras que las pendientes más pronunciadas (>35°) limitan el establecimiento por la inestabilidad del terreno y la falta de suelo desarrollado (Torres-Beltrán et al., 2017). Por otro lado, se ha documentado que las pendientes suaves retienen mejor la humedad del suelo, facilitando el desarrollo inicial de los árboles jóvenes (Astudillo-Sánchez et al., 2019). Y a su vez, también se ha observado que en los bosques de la línea de árboles estable tiende a ubicarse en pendientes más pronunciadas, donde la capacidad de acumulación de agua derivada de la escorrentía favorece su crecimiento (Xu et al., 2020). En nuestro estudio, encontramos evidencia de avance de la línea de árboles en una amplia gama de pendientes, predominando en micrositios con pendientes llanas y ligeras, pero también ocurriendo en pendientes muy inclinadas y extremas.
La temperatura y la humedad también influyen en la dinámica de la vegetación. Los valores más bajos de temperatura y del índice de humedad del suelo (SMI) en los Grupos 1 y 2 que mostraron un predominio del cambio de bosque a pastizal, refuerzan la idea de que las condiciones más secas y cálidas pueden promover la conversión de cobertura forestal a cobertura de pastizal. Aunque las especies de alta montaña están adaptadas a bajas temperaturas, la temperatura media anual (6 °C) podría no ser suficiente para alcanzar el nivel óptimo de fotosíntesis, que solo ocurre en los momentos de máxima irradiación (Körner, 2012; Sanfuentes et al., 2012). En contraste, el Grupo 4 mostró los valores más altos de SMI, lo que coincide con la predominancia del cambio de pastizal a bosque en estas áreas, subrayando la importancia de la humedad en el establecimiento de los árboles. Pompa-García et al. (2023) sugieren que en la línea de árboles la menor temperatura y disponibilidad de agua en el suelo limitan la actividad fotosintética. La vegetación se concentra en áreas donde la orientación reduce la sombra de ladera y permite mayor acceso a la luz solar. Por lo que, tanto la cantidad como la estacionalidad de la precipitación juegan un papel crucial en la distribución de los sistemas vegetales tras el establecimiento, una vez que la humedad queda almacenada en el suelo y las temperaturas son óptimas (Astudillo-Sánchez et al., 2017).
Conclusiones
La topografía y el clima son factores complementarios que explican el avance de la línea de árboles. Los resultados de nuestro estudio revelan un patrón de avance espacialmente heterogéneo y en pulsos temporales que sugieren la influencia de factores adicionales más allá del calentamiento global. El análisis de los cambios en la línea de árboles y en la vegetación indica una tendencia general hacia el avance del bosque y una reducción del pastizal desde 1985. Sin embargo, los resultados también destacan la complejidad y variabilidad de los cambios en función de factores topográficos y climáticos. El movimiento de la línea de árboles presenta variaciones espaciales locales debido a las diferencias en temperatura y humedad del suelo que se traducen en la presencia de micrositios de colonización que favorecen el establecimiento. Estos sitios podrían representar zonas en las que se favorece el avance de los árboles debido a que amortiguan las condiciones adversas y permiten el establecimiento. Se requiere seguir explorando la interacción de los árboles en sus primeras etapas del desarrollo con la variedad de ambientes presentes en la línea de árboles, para lograr un mejor entendimiento de las dinámicas de la línea de árboles.

