











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El desplazamiento de especies fuera de sus áreas de distribución natural, con la intervención humana de forma intencional o accidental, ha favorecido las invasiones biológicas (Gutiérrez 2006). Las invasiones biológicas son procesos naturales, que no son exclusivamente provocadas por los humanos (Vantarová et al. 2023). Sin embargo, en los últimos 150 años, la frecuencia de estas invasiones ha experimentado un incremento, resultando en la homogenización biótica de una porción de la biodiversidad terrestre y acuática, fenómeno raramente observado en la historia de la vida en la Tierra (Olden y Poff 2003, Daly et al. 2023). Esta situación se debe a la concurrencia de múltiples factores, como la expansión de la frontera agrícola, la predominancia del monocultivo, la deforestación, la fragmentación de hábitats, la desertización, las demandas alimentarias asociadas al crecimiento demográfico, el cambio climático, la contaminación, la sobreexplotación de los recursos, el comercio internacional, la acuicultura, la pesca y el turismo (McNeely et al. 2001).
América Latina y el Caribe han enfrentado las invasiones biológicas de manera similar al resto del mundo (Pauchard et al. 2011). En particular, la biodiversidad de países megadiversos como Ecuador podría estar en riesgo debido a las especies exóticas invasoras, ya que es una de las más ricas del planeta y está concentrada en un espacio bastante reducido (Ballesteros-Mejia et al. 2021, Kleemann et al. 2022). La gran diversidad biológica de Ecuador se debe a tres factores clave: su ubicación geográfica en la zona ecuatorial, la elevación de la cordillera de Los Andes y la influencia de las corrientes marinas en la zona costera (Mittermeier et al. 1997). Especialmente en las islas Galápagos, se han reportado numerosas invasiones de especies exóticas, como consecuencia del incremento de las actividades turísticas y el desarrollo económico (Muñoz-Barriga 2015). El aumento en la cantidad de invasores que ocupan los diversos ecosistemas en las Galápagos representa la amenaza principal para la biodiversidad, ya que desplazan a las especies autóctonas y endémicas, provocando su pérdida (Ballesteros-Mejia et al. 2021). También tienen un impacto indirecto al competir por alimentos, sitios de reproducción y anidamiento, degradar el hábitat y perturbar los procesos ecológicos y evolutivos (Torres et al. 2023).
Entre las especies exóticas invasoras que están causando graves daños a nivel mundial y en Ecuador están algunas especies de hormigas (Salazar-Basurto et al. 2023). De forma general, en el país existen registradas 11 subfamilias, 97 géneros, 829 especies y subespecies de hormigas, de las cuales 75 son endémicas y 21 son introducidas (Antweb 2023). Nylanderia fulva (Mayr), conocida como la hormiga loca sudamericana, se encuentra entre las especies más peligrosas a nivel global debido a su potencial invasivo (Wang et al. 2016), y su presencia ha sido registrada recientemente en Ecuador continental (Pazmiño-Palomino et al. 2020). Es una hormiga pequeña, colonial que se comporta como plaga invasora en varios países, causando impactos económicos, ecológicos y sociales (Mendoza et al. 2019). Principalmente, esta especie invasora ataca y desplaza a la fauna silvestre; además, invade cultivos donde se asocia en simbiosis con insectos chupadores para alimentarse. Se ha documentado que también ataca animales domésticos, impide o limita las labores agrícolas y ocupa las habitaciones humanas (Arcila y Quintero 2005, Gruber et al. 2022).
Ante esta situación alarmante, es necesario identificar a priori las posibles áreas que puedan ser invadidas por N. fulva en Ecuador, de manera tal que pueda servir como línea base para el desarrollo de estrategias de prevención o control. Tomando en consideración lo expuesto, el objetivo de esta investigación fue evaluar el riesgo de invasión de la hormiga loca sudamericana en Ecuador. Para ello, se modeló la distribución geográfica potencial de N. fulva en Ecuador.
Materiales y métodos
Área de estudio
La investigación se llevó a cabo en Ecuador, país que incluye una porción continental y las Islas Galápagos. La parte continental se sitúa al Noroeste de América del Sur (01° 28’ N y 05° 01’ S; 75° 11’ O y 81° 01’ O), limitando al norte con Colombia, al sur y al este con Perú (Farfán 2018). El territorio ecuatoriano se encuentra dividido en tres regiones naturales distintas en términos de topografía, clima, vegetación y población, conocidas como la Costa o Litoral, la Sierra o Región Andina y el Oriente o Amazonía. Adicionalmente, las Islas Galápagos forman la cuarta región del país, ubicada en el océano Pacífico, a una distancia de 972 km de la costa continental de Ecuador.
Datos de presencia de la especie
Se realizó una búsqueda en la base de datos Global Biodiversity Information Facility (GBIF) con la herramienta NicheToolBox (Osorio-Olvera et al. 2020), para obtener registros (coordenadas geográficas) de la hormiga loca dentro de su rango nativo y de invasión a nivel mundial. Conjuntamente, se utilizaron los registros de presencia documentados en el informe final del proyecto: Digitalización de datos de especies invasoras del Ecuador a nivel nacional y regional (CDC-Ecuador 2008), en el informe de Especies Exóticas Invasoras en Reservas de Biosfera de América Latina y el Caribe (Schüttler y Karez 2008), en el Cuarto informe para la Convención de Diversidad Biológica (Ministerio del Ambiente de Ecuador 2010) y de la lista GRIIS (GRIIS Checklist of Introduced and Invasive Species - Ecuador) en GBIF (Herrera et al. 2019). Adicionalmente, se obtuvieron otros datos de presencia en la región nativa y de invasión a partir de literatura científica (Herrera 2013, Kumar et al. 2015).
De manera preliminar, se obtuvieron 503 registros de presencia de la hormiga a nivel mundial, los cuales se organizaron por países y años. Este conjunto inicial de registros de presencia se depuró para reducir posibles sesgos de muestreo y de calibración de los modelos de nicho ecológico (Syfert et al. 2013) que conducen a agrupaciones artificiales (Alkishe et al. 2017). Primero, se descartaron aquellos registros con referencias geográficas desconocidas, o con errores como coordenadas incompletas, ubicaciones en el océano y desajustes entre metadatos y coordenadas. Posteriormente, con la herramienta NicheToolBox se eliminaron todos los registros duplicados dentro de un tamaño de píxel de 1 km para minimizar la autocorrelación espacial. Este filtrado de los datos por distancia omite registros redundantes y permite una mejor estimación de las características del nicho (Fourcade et al. 2013). Al final, los registros se redujeron a 82, de los cuales 49 se localizaron en territorios invadidos (solo cuatro en Ecuador) y 33 en el área nativa de distribución.
Variables climáticas
Se tomaron 15 de las 19 variables bioclimáticas disponibles en el WorldClim 2.0, con una resolución de 30 arc-segundos (~1 km) (Fick y Hijmans 2017), como base para la representación ambiental y climática del área de distribución de la especie. Se descartaron cuatro variables: temperatura promedio del trimestre más húmedo (bio8), temperatura promedio del trimestre más seco (bio9), precipitación en el trimestre más caliente (bio18) y precipitación en el trimestre más frío (bio19), ya que está comprobado que forman parte de artificios matemáticos que no se relacionan con la especie (Alkishe et al. 2017). La información de clima se extrajo de los puntos de presencia de la hormiga en la región nativa y de invasión, con ArcGIS v10.1 (ESRI 2011).
Posteriormente, se combinaron diferentes métodos para seleccionar solo las variables adecuadas para obtener el modelo de N. fulva, ya que la eliminación de variables predictivas correlacionadas aumenta el rendimiento de los modelos (Cooper et al. 2016). En primer lugar, se revisó la ecología de la especie, para así poder determinar cuáles variables climáticas aparecen descritas como más importantes para su desarrollo (Eyer et al. 2018). En segundo lugar, se revisó la literatura científica donde se modeló con anterioridad la distribución potencial de la especie, para así comprobar cuáles variables ambientales han sido más importantes en estos modelos (Kumar et al. 2015). En tercer lugar, se realizó un análisis de correlación bivariada entre las 15 variables climáticas, para identificar las correlaciones más fuertes y seleccionar una variable de cada par correlacionado, con un umbral de r = 0.8. En cuarto lugar, se realizó un Análisis de Componentes Principales (ACP), para usar el peso de cada variable en relación a los componentes como una vía para identificar su relevancia y aporte a la varianza de los datos. Se consideraron como más relevantes las variables que tuvieron un peso mayor a 0.7. En quinto lugar, se calibró un modelo de máxima entropía con las 15 variables para identificar aquellas de mayor aporte. Una vez resumidas las variables más importantes por cada método (Tabla 1), se eliminaron las de menor contribución en cada uno. Finalmente, quedaron solo las más contribuyentes desde el punto de vista ecológico y estadístico: temperatura media anual, temperatura máxima del mes más cálido, precipitación en el mes más húmedo y precipitación del trimestre más húmedo (Tabla 1), las cuales se utilizaron para obtener los modelos con el algoritmo MaxEnt.
Tabla 1 Variables climáticas más importantes para Nylanderia fulva según diferentes formas de análisis.
| Influencia en la ecología de la especie | Literatura de modelación para la especie | Correlación entre variables | Análisis de Componentes Principales | Algoritmo de máxima entropía (MaxEnt) |
|---|---|---|---|---|
| Temperatura media anual (Bio1) | Temperatura media anual (Bio1) | Estacionalidad de la temperatura (Bio4) | Temperatura media anual (Bio1) | Temperatura media anual (Bio1) |
| Rango de temperatura anual (Bio7) | Temperatura máxima del mes más cálido (Bio5) | Temperatura máxima del mes más cálido (Bio5) | Estacionalidad de la temperatura (Bio4) | Temperatura máxima del mes más cálido (Bio5) |
| Precipitación anual (Bio12) | Temperatura mínima del mes más frío (Bio6) | Temperatura mínima del mes más frío (Bio6) | Temperatura máxima del mes más cálido (Bio5) | Precipitación en el mes más húmedo (Bio13) |
| Estacionalidad de la precipitación (Bio15) | Precipitación anual (Bio12) | Rango de temperatura anual (Bio7) | Temperatura mínima del mes más frío (Bio6) | Precipitación del trimestre más húmedo (Bio16) |
| Precipitación en el mes más húmedo (Bio13) | Precipitación en el mes más húmedo (Bio13) | Rango de temperatura anual (Bio7) | ||
| Precipitación en el mes más seco (Bio14) | Precipitación en el mes más seco (Bio14) | Precipitación anual (Bio12) | ||
| Precipitación del trimestre más húmedo (Bio16) | Precipitación en el mes más húmedo (Bio13) | |||
| Precipitación del trimestre más húmedo (Bio16) |
Modelación de nicho ecológico
Se empleó el algoritmo de máxima entropía MaxEnt 3.4.4. (Peterson 2003) para modelar el nicho ecológico de la hormiga loca. Se usaron las asociaciones entre los puntos de presencia de la especie y las variables climáticas para construir el nicho ecológico. Para calibrar el modelo se determinó el área accesible adecuada (M), que se define como el área a la que la especie probablemente tuvo acceso a través de la dispersión (Barve et al. 2011). La calibración del modelo se debe realizar a través de esta área ya que mejora el rendimiento del mismo (Anderson y Raza 2010) y minimiza los impactos de supuestos sobre la ausencia de la especie en áreas que no son accesibles para la misma. Se consideró que el área accesible incluye los países nativos y de invasión donde existieron puntos de presencia de la hormiga. A esto se suma el número restringido de registros de N. fulva en Ecuador, que no capturaban la representación climática en la que es posible encontrar a la especie. Por ello, se calibraron los modelos con los datos de presencia en la región nativa y de invasión mundial y se proyectaron en el espacio geográfico de invasión en Ecuador, para así identificar las áreas cuyas condiciones climáticas, tanto en las regiones nativas como en las de invasión, son similares a las de Ecuador y adecuadas para la especie.
Se generaron 100 réplicas del modelo, para evitar asunciones incorrectas por artefactos estadísticos. Se usó el boostrap como método de remuestreo. El 80% de los registros de presencia fue utilizado para calibrar el modelo y el otro 20% para evaluarlo. Dentro de las 100 iteraciones, se eligió el modelo que presenta la mediana de los valores, evitando así la influencia de los valores atípicos sobre la media. Para validar el modelo se tuvo en cuenta el valor del AUC, y se tomó como guía para la clasificación de los modelos la propuesta realizada por Araujo y Guisan (2006): AUC = 0.5-0.6 insuficiente; 0.6-0.7 pobre; 0.7-0.8 promedio; 0.8-0.9 bueno; 0.9-1 excelente. Además, se usó el partialROC (Peterson et al. 2011) para probar la robustez de los modelos a través de Niche Toolbox (Osorio-Olvera et al. 2020). Se consideraron como parámetros una tasa de omisión de 5%, un porcentaje de puntos aleatorios de 50% y 500 iteraciones de bootstrap (Peterson et al. 2008).
A menudo, las predicciones de umbral reflejan las suposiciones de los investigadores sobre los valores de umbral apropiados y no los atributos de la distribución de especies (Merow et al. 2013). Sin embargo, para evitar problemas con el sobreajuste de los modelos (Peterson et al. 2007), se estableció como umbral de corte el 10mo percentil (Liu et al. 2013). A todos los píxeles con un valor por debajo de este umbral se les asignó un valor de cero (0), lo que representaría la ausencia de condiciones adecuadas para la especie. Los mapas binarios (favorable/no favorable) obtenidos permitieron la identificación de las regiones geográficas con condiciones climáticas adecuadas para N. fulva. La diferencia entre el valor de umbral de corte (0.29) y el valor de probabilidad máxima (0.948) se dividió por tres, lo que arrojó un nuevo valor de corte que se utilizó para clasificar el mapa en tres nuevas categorías. Estas categorías incluyen la distribución de píxeles por encima del valor umbral (0.29): baja (29 - 51%), media (51 - 73%) y alta probabilidad (73% - 95%) de condiciones favorables para la invasión de la especie.
Se evaluó la importancia de las variables climáticas en los modelos comparando los valores obtenidos a través del jackknife de MaxEnt con los datos de entrenamiento, al correr el modelo con las variables individuales. Asimismo, se llevaron a cabo análisis de Similitud de Superficie Ambiental Multivariada para determinar la similitud entre los climas presentes en las regiones geográficas donde se proyectaron los modelos (Zurell et al. 2012).
Resultados
Se obtuvieron registros de presencia de N. fulva en Guayana Francesa y 11 países, tres de distribución nativa (Argentina, Brasil y Paraguay) y ocho que ha invadido (Costa Rica, Cuba, Colombia, Ecuador, México, Estados Unidos, España y Granada). Para los datos de entrenamiento, el valor del AUC para el modelo de distribución geográfica potencial de N. fulva fue 0.9 ± 0.01, mientras que el de validación fue 0.88 ± 0.03, por lo que tuvo buen rendimiento a la hora de predecir los sitios con condiciones ambientales adecuadas para la especie. Las predicciones dieron proporciones de área bajo la curva (AUCratio) mayores que las expectativas nulas en el análisis ROCparcial (p < 0.001). El AUCratio tuvo un valor promedio de 1.5 ± 0.33, lo que indica buen rendimiento según las variables utilizadas, y predice de forma adecuada y mejor que un modelo al azar.
La temperatura promedio anual fue la variable que más aportó al modelo de N. fulva, con un 73.8 ± 12% de contribución y un 74.6 ± 14.4% de importancia en la permutación. La contribución de la precipitación del mes más lluvioso fue baja con 22.8 ± 11.9% y 15.7 ± 11.4% de importancia. De acuerdo con la respuesta de la idoneidad ambiental para la especie, en relación con estas dos variables, se observa que los lugares con una temperatura media anual inferior a 20 °C presentan una baja idoneidad para la especie (Figura 1A), de igual forma los que presentan temperaturas superiores. Pero se observó un segundo pico de respuesta óptima a la favorabilidad a los 28 °C (Figura 1A). La respuesta a la precipitación en el mes más lluvioso tuvo un comportamiento similar, sitios con valores de alrededor de los 200 mm de precipitación tienen una respuesta óptima de la favorabilidad, por encima o por debajo de este valor la respuesta disminuye (Figura 1B).
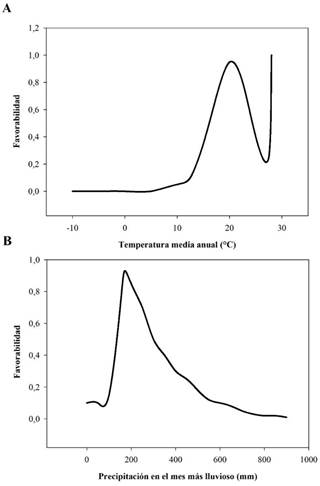
Figura 1 Respuesta de la favorabilidad ambiental para la hormiga loca (Nylanderia fulva) ante variaciones de la temperatura media anual (A) y la precipitación en el mes más lluvioso (B) en Ecuador.
El área favorable para la invasión de N. fulva en Ecuador tuvo una extensión de ~110 227 km2. En gran parte de la región Costa y la región Amazónica, N. fulva podría encontrar áreas con condiciones climáticas adecuadas para su establecimiento, zonas que tienen un clima tropical y condiciones climáticas semejantes a la región nativa de la especie. La región Sierra apareció como no adecuada para esta especie y en las Islas Galápagos se identificaron zonas núcleos con condiciones climáticas adecuadas dentro de cada isla (Figura 2).
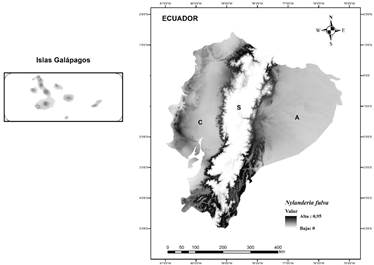
Figura 2 Favorabilidad ambiental para la distribución potencial de Nylanderia fulva (hormiga loca) en Ecuador. C: región Costa; S: región Sierra; A: región Amazonía.
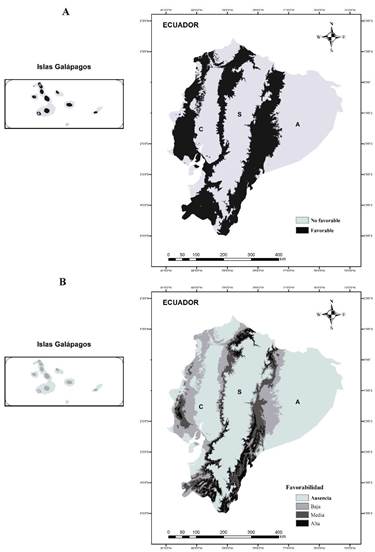
Se distinguieron de forma clara zonas que bordean a la región Sierra y en la franja costera como sitios favorables (Figura 3A), de igual forma dentro de cada isla del archipiélago de Galápagos. Aun así, dentro de estos sitios favorables, las regiones con mayor valor de favorabilidad se extienden solamente en ~18 892 km2 bordeando a la sierra en la porción norte y sur (Figura 3B). Esta extensión representa solamente el 17% de las zonas con condiciones adecuadas en el país. El mayor porcentaje (53.4%) corresponde con áreas de baja favorabilidad (~58 884 km2), fundamentalmente hacia las Islas Galápagos, la zona costera continental y parte de la Amazonía (Figura 3B).
Discusión
La distribución potencial de N. fulva en Ecuador se ajusta a las ocurrencias que actualmente se conocen en los países invadidos de América del Sur, en los cuales hay climas similares. Por esta razón, y como lo predijo el modelo, en territorio ecuatoriano la hormiga loca tiene disponibles extensas áreas con clima adecuado fuera de su región de distribución nativa (Kumar et al. 2015). La metodología aplicada, identificó las zonas con mayor probabilidad de invasión. Los territorios que potencialmente pueden ser invadidos en Ecuador por la especie N. fulva, con condiciones climáticas adecuadas para su desarrollo, están definidos. Este trabajo toma como antecedente el primer reporte realizado en el país para esta especie en el año 2020 (Pazmiño-Palomino et al. 2020), el cual debe ser tomado en serio dada la gravedad. La introducción accidental o intencional de hormigas, mediante la intervención humana, en zonas donde dichas especies no se encuentran presentes de manera natural, puede provocar una invasión biológica con consecuencias irreparables para los ecosistemas invadidos, la salud humana y la economía (Ballesteros-Mejia et al. 2021, Gruber et al. 2022, Wong et al. 2023). Dentro de las especies invasoras, las hormigas constituyen uno de los grupos más peligrosos y algunas especies, como Linepithema humile, Pheidole megacephala, Anoplolepis gracilipes, Solenopsis invicta, han sido catalogadas entre las peores a nivel mundial (Lowe et al. 2000, Rabitsch 2011). La capacidad invasora de la hormiga loca se atribuye a sus características anatómicas y ecológicas, como su pequeño tamaño, alta movilidad, altas densidades, capacidad de adaptación y dominancia dentro de las comunidades de invertebrados (Wang et al. 2016, Sunyong et al. 2018).
El antecedente de invasión más cercano a Ecuador, que demuestra el peligro que representa N. fulva, se registró en Colombia, donde fue introducida entre 1969 y 1970 en la zona cafetera del centro y en el Valle Medio del Magdalena, como agente de control biológico contra las hormigas cortadoras de hojas y serpientes (Zenner-Polania 1991). La capacidad de la hormiga loca para colonizar eficazmente nuevos hábitats se debe a su ventaja sobre las especies nativas, con tasas de expansión del área de distribución de aproximadamente 100 m al mes (Zenner-Polania 1991). En Ecuador, se encuentran condiciones climáticas y ecológicas similares a Colombia, lo que favorece la proliferación de esta especie invasora. Por esta razón, transcurridos cinco años a partir de su primer reporte en territorio ecuatoriano continental, N. fulva puede estar en un proceso inicial de colonización, que de no detenerse puede conllevar a consecuencias desastrosas.
El primer registro de la hormiga loca en Ecuador fue en áreas de cultivo de caña de azúcar (Pazmiño-Palomino et al. 2020) y es precisamente en este tipo de plantaciones donde se ha reportado como una de las especies más frecuentes, así como en los domicilios humanos en ambientes urbanos (Fontenla-Rizo y Matienzo-Brito 2011, Zenner-Polanía 2019). Uno de los principales cultivos del Ecuador es la caña de azúcar, si se considera la superficie cultivada, el valor producido y la cantidad de empleos directos que genera (Cartay-Ángulo et al. 2019). La caña de azúcar se cultiva en casi todo el país, sin embargo, la producción está concentrada en las provincias Guayas, Cañar, Carchi e Imbabura. En solo tres provincias Guayas, Loja y Cañar, se produce cerca de un 97% del volumen total de todo Ecuador (Cartay-Ángulo et al. 2019). Varias de estas áreas azucareras por excelencia se localizan dentro de las localidades que el modelo predice como potencialmente adecuadas para el desarrollo de la especie. Por esta razón, las probabilidades para que la especie se expanda son mucho mayores, ya que no solo cuenta con las condiciones climáticas adecuadas, sino con hábitats que son de su preferencia. De propagarse N. fulva en el interior de los campos de caña de azúcar, pudiera provocar serias afectaciones a uno de los renglones productivos de mayor importancia en el país.
Según el modelo obtenido, las regiones idóneas para N. fulva aparecen principalmente en la región Costa y la región Amazónica del Ecuador. En estas regiones, desde el punto de vista ecológico, se concentra gran parte de la biodiversidad del país. La región de la Amazonía se ha catalogado como uno de los sitios con mayor diversidad biológica a nivel mundial (Funk et al. 2012) y la presencia de esta especie de hormiga invasora pudiera afectar el equilibrio en estas zonas tan sensibles. La región Costa se caracteriza por incluir zonas muy húmedas, las cuales, según el modelo, son muy adecuadas para soportar poblaciones de N. fulva.
La presencia de N. fulva en Ecuador continental fue predicha por un modelo de distribución potencial global más general (Kumar et al. 2015). Estos autores propusieron que esta especie se ha trasladado a lugares más cálidos y secos entre el rango nativo e invasivo, adoptando un mayor potencial para colonizar nuevas áreas. Por estas razones, es fundamental monitorear el progreso de la invasión de N. fulva en Ecuador, especialmente con un escenario de cambio climático que podría facilitar su colonización. Ante esta situación, se recomienda incentivar el diseño y la realización de investigaciones con relación a la invasión de esta especie en Ecuador, que den seguimiento al desarrollo y evolución de sus poblaciones en su área de distribución en el país, así como el impacto provocado a nivel ecológico y económico. Además, crear alianzas entre las entidades ambientales y gubernamentales para promover y hacer cumplir las medidas preventivas y de control en torno a su invasión (LeBrun et al. 2022), ya que, de no llevarse a cabo con suficiente tiempo, las consecuencias pueden ser catastróficas.
Conclusiones
Las regiones idóneas para N. fulva se encuentran en la región Costa y la región Amazónica de Ecuador, donde se concentra gran parte de la biodiversidad del país. La hormiga loca se podría trasladar a lugares más cálidos y secos por su potencial para colonizar nuevas áreas, lo que implica un llamado de atención a las autoridades ecuatorianas. Es necesario establecer un plan para monitorear el progreso de la invasión de N. fulva en Ecuador, especialmente con un escenario de cambio climático que podría facilitar su colonización.

