











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La zooarqueología o arqueozoología es la ciencia que investiga los restos de la fauna del pasado preservados en contextos arqueológicos. Se centra en la identificación taxonómica y anatómica de las evidencias arqueobiológicas y el análisis de las interacciones entre las sociedades pasadas, el medio ambiente y las poblaciones animales en el tiempo y el espacio. Este enfoque particular, se interesa en conocer las dimensiones culturales, biológicas y ecológicas de los usos históricos de la fauna y su repercusión en la formación y distribución de los depósitos de restos arqueofaunísticos y en la conservación de la biodiversidad actual (Davis, 1987; Peña-León, 2006; Reitz y Wing, 2008; Gutiérrez et al., 2010; Gifford-González, 2018).
En la búsqueda de reconstruir la historia de procesamiento de los restos óseos de los animales cazados y domesticados por los seres humanos en el pasado, la zooarqueología adopta las herramientas metodológicas y preguntas epistemológicas de la tafonomía, una ciencia transdisciplinar que estudia los procesos físicos, químicos y biológicos que suceden con posterioridad a la muerte de los organismos, abarcando las fases de descomposición, desarticulación, dispersión, transporte, depositación, inhumación e incorporación de los restos orgánicos en la matriz sedimentaria, dando origen a la fase diagenética del proceso tafonómico. En esta fase continúa la modificación de las características físicas y químicas del hueso hasta su fosilización o eventual traslocación y reelaboración (Efremov, 1940; Behrensmeyer et al., 1985; Aguirre, 1997; Lyman, 2010; Lloveras et al., 2016).
Los estudios zooarqueológicos llevados a cabo en el Caribe colombiano se han centrado principalmente en las siguientes líneas de investigación: 1) estudios sobre subsistencia y alimentación humana en contextos arqueológicos, los cuales presentan menciones específicas de evidencias tafonómicas en los restos de fauna (Rojas-Mora & Montejo-Gaitán, 1999; Stahl & Oyuela-Caycedo, 2007; Ramos-Roca & Archila, 2008; Álvarez-León & Maldonado-Pachón, 2009; Lozano, 2014; Castro et al., 2018; Flórez-Correa 2018a; 2020; Niño-Méndez, 2018; Ramos-Roca & Niño-Méndez, 2019; Gutiérrez-Ferro, 2022; 2024); 2) estudios paleoambientales, paleoecológicos y paleoclimáticos sobre peces (Rodríguez & Peña-León, 2001; Peña-León, 2001; Diaz-Chauvigne, 2016) algunos de los cuales han aplicado estudios físico-químicos para la caracterización de variables tafonómicas (Flórez-Correa, 2023; Flórez-Correa et al., 2024), y sobre malacofauna en concheros litorales (Carvajal-Contreras, 2012; 2013a; 2013b; 2019) y en contextos funerarios (Flórez-Correa, 2019), los cuales han evidenciado la modificación de las conchas marinas y dulceacuícolas para la elaboración de artefactos; 3) estudios etnoarqueológicos y etnohistóricos sobre el conocimiento y uso de la fauna local (Carvajal-Contreras, 2015; 2022; Ramos-Roca, 2014; 2020; Ramos-Roca & Jiménez, 2015; Márquez-Prieto, 2017) entre los cuales, trabajos como los de Flórez-Correa (2019b) y Baquero-Soto (2022) profundizan en la cacería, procesamiento y formación de depósitos óseos de fauna analizando las variaciones tafonómicas de los animales sacrificados y aprovechados por comunidades actuales; y 4) otros trabajos se han centrado puntualmente en estudios experimentales y arqueométricos sobre alteraciones térmicas (Ramos-Roca & Campos, 2014), caracterizando evidencias tafonómicas de la exposición al fuego en la histología ósea, el colágeno, la hidroxiapatita (bioapatita) y el ADN antiguo (ADNa) (Rojas-Mora & Flórez-Correa, 2023; Flórez-Correa, 2023; Flórez-Correa et al., 2024).
La presente investigación zooarqueológica y tafonómica se contextualiza en el sitio San Pedro de la Depresión Momposina, un antiguo asentamiento indígena localizado en el municipio de San Benito Abad en Sucre, Colombia, donde se han hallado depósitos arqueológicos con abundante cerámica, carbón y arqueofauna acuática. De esta última línea de evidencia, destaca la abundancia relativa de una especie en particular, el ponche (Hydrochoerus isthmius Goldman 1912, Rodentia, Mammalia). Su distribución en los depósitos derivados de los residuos de la alimentación de las comunidades, da cuenta de su aprovechamiento intensivo en la paleodieta humana. Estas evidencias llevan a plantear la pregunta acerca de la importancia cultural de los ponches y su depositación en el contexto arqueológico de estudio, así como las implicaciones para la conservación pasada y presente de la especie. En ese sentido, el objetivo de esta investigación fue caracterizar las evidencias tafonómicas del procesamiento antrópico de H. isthmius en los depósitos zooarqueológicos del sitio San Pedro y reconstruir las formas de aprovechamiento llevadas a cabo por la población humana que habitó el yacimiento (Flórez-Correa, 2018a; 2023; Flórez-Correa et al., 2024).
También conocido como poncho, chigüiro, cacó, carpincho o capibara menor según la región, es considerado uno de los roedores más grandes del mundo pertenecientes al género Hydrochoerus Brisson 1762, de la familia Caviidae. Anteriormente se sostenía la existencia de una especie, Hydrochoerus hydrochaeris, y dos subespecies: Hydrochoerus hydrochaeris hydrochaeris Linnaeus 1766, así como Hydrochoerus hydrochaeris isthmius. Sin embargo, sus diferencias cariotípicas (H. hydrochaeris 2n=66; H. isthmius 2n=64) y moleculares, indican que corresponden a dos especies distintas (Wilson & Reeder, 2005; Carrascal et al., 2011; Chacón et al., 2013; Moreira et al., 2013). Ambas especies varían en tamaño, forma y peso, en asociación con sus preferencias de hábitat y dieta particulares. Los pesos máximos reportados incluyen valores de hasta 48 kg para H. isthmius, y 50 kg para H. hydrochaeris, en los Llanos de Venezuela (Mones & Ojasti, 1986), y hasta 100 kg en zonas agrícolas del Medio Oeste de Brasil (Felix et al., 2014). Morfológicamente se diferencian en el ángulo que se forma entre el neurocráneo y el esplacnocráneo. H. hydrochaeris exhibe una transición recta y frontales más anchos. H. isthmius presenta una inclinación de la región nasal en sentido ventral y diastema inferior proporcionalmente más larga, así como un proceso pterigoideo más grueso y corto que H. hidrochaeris (Fig. 1). Sin embargo, ambas especies tienen trayectorias ontogenéticas similares (Aeschbach et al., 2016).

Adaptado de: https://colombia.inaturalist.org/taxa/74443-Hydrochoerus-isthmius/ Link de licencia: https://creativecommons.org/licenses/by-nc/4.0/
Figura 1. Hydrochoerus isthmius. Fotografía de Jesús Arana.
El origen del género Hydrochoerus spp. se remonta al Pleistoceno en el sur de los Estados Unidos, Centro América y Sur América (Ojasti, 1973). La distribución biogeográfica de H. hydrochaeris se extiende al oriente de la Cordillera de los Andes en gran parte de Suramérica en países como Colombia, Venezuela, Suriname, Guyana, Guyana Francesa, Brasil, Ecuador, Bolivia, Uruguay, Paraguay y Argentina. Su registro fósil se ha reportado desde el Pleistoceno Tardío en países como Curaçao, Uruguay, Brasil y Argentina (Ballesteros & Jorgenson, 2009). Por su parte, H. isthmius de menor tamaño, se distribuye desde el área central de Panamá hasta el norte de Colombia y el noroeste de Venezuela en la cuenca del Lago de Maracaibo, aislada por los Andes de Mérida y la Serranía del Perijá (Mones, 1991; Kerber & Ribeiro, 2011; Solari et al., 2013) (Fig. 2). En Colombia las poblaciones son frecuentes en las vertientes del océano Atlántico, en las sabanas ubicadas al norte de la cordillera de Los Andes y en las áreas inundables del río Magdalena, así como en las cuencas del río Mira, el Atrato, el Medio y Alto Sinú, y el Medio San Jorge en el piso térmico cálido de 0-500 msnm. Esta especie no presenta registro fósil conocido (Ballesteros & Jorgenson, 2009; Chacón et al., 2013; Moreira et al., 2013; Carbot-Chanona et al., 2020).
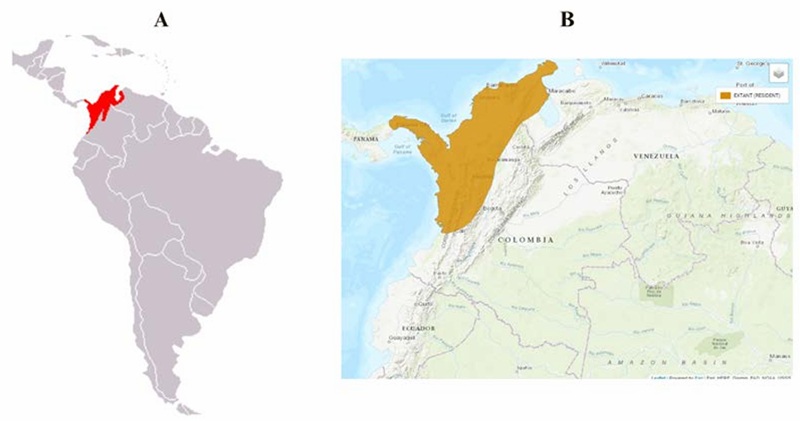
Figura 2. Distribución biogeográfica de H. isthmius. A) Adaptado de: https://colombia.inaturalist.org/taxa/74443-Hydrochoerus-isthmius. B) Adaptado de https://www.iucnredlist.org/species/136277/22189896
Los registros del género Hydrochoerus en contextos arqueológicos suramericanos incluyen reportes de H. hydrochaeris desde Argentina al sur, principalmente en sitios del Holoceno Tardío vinculados con zonas bajas de la llanura aluvial del Paraná en la provincia de Buenos Aires (Acosta et al., 2010; Sartori & Colasurdo, 2012; Servin, 2016), como Río Luján, Cañada, Honda e Isla (Politis, 1985; Salemme, 1987), al igual que en varios sitios de la provincia de Santa Fe como Playa Mansa (Feuillet-Terzaghi, 2002), Barrancas del Paranacito, Cerro Aguará, y el sitio histórico Juan Manuel de Rosas de la ciudad de Rosario datado entre el siglo XVIII y finales del XIX d.C. (Colasurdo, 2014). En Brasil se ha registrado el uso de H. hydrochaeris desde 8000 a 7500 A.P. a partir de pictografías al interior de sistemas kársticos en Piauí (Guidon & Delibrias, 1986), y hay reportes de restos óseos en distintos sitios arqueológicos (Prous, 1992) como el SC PRV 02 de la Isla de Santa Catarina (1067-1735 años A.P.), donde se presentó la extinción local de la especie hace apenas 40 años (De Castilho & Simões-Lopes, 2005).
La especie H. isthmius ha sido reportada en sitios arqueológicos de la región Noroccidental de Colombia como el yacimiento Arrancaplumas del período Formativo Tardío (siglos V-I a.C.), localizado en la zona de Raudales del río Magdalena en el municipio de Honda, Tolima (Peña, 2010). También se ha reportado en el área arqueológica de Malambo del período Formativo Tardío (3070±200 años A.P.), localizada en la zona occidental del río Magdalena en el departamento del Atlántico (Angulo, 1981). Otros sitios incluyen el Valle de Santiago (900-1210 años a.C.) (Angulo, 1983) y a 20 km de allí, en la Serranía del Caballo, sobre la margen Occidental de la Ciénaga del Guájaro (Angulo, 1988), al igual que en el sitio Santa Fé del período Tardío (siglos XIV-XVI d.C.) ubicado a orillas del río Magdalena en el municipio de Magangué, Bolívar (Álvarez, 2022). A su vez, hay registros en la cuenca del Bajo río San Jorge en yacimientos de la Depresión Momposina (Plazas et al., 1993), como el sitio San Pedro del Caño Rabón (Flórez-Correa, 2018) y el sitio Tronconudo del Caño Carate (Flórez-Correa, 2025). También en el sitio Momil de la etapa Formativa (2150±60 años A.P.), ubicado en la cuenca del río Sinú (Reichel-Dolmatoff & Dusán, 1956), y en el contexto funerario hallado en el Colegio Nacional José María Córdoba, localizado en la ciudad de Montería, Córdoba. En este yacimiento se identificó un fémur izquierdo de H. isthmius junto con cinco individuos humanos adultos y dos subadultos, inhumados en una urna funeraria de un total de siete recuperadas en el sitio, que datan del período comprendido entre los siglos II a.C. y X d.C., indicando la importancia simbólica y cultural de esta especie para las comunidades que se asentaron en la región (Castillo & Lalinde, 2010; Puche, 2018).
Biología y Ecología de Hydrochoerus isthmius . El ponche es un animal herbívoro que se adapta a distintos tipos de hábitat que poseen en común un cuerpo de agua, principalmente ciénagas, pantanos, meandros abandonados, manglares, esteros, canales de drenaje, praderas inundables, madres viejas, ríos y arroyos, los cuales usan para bañarse, beber agua, copular y refugiarse de sus depredadores, al igual que áreas de esparcimiento como pastizales, bosques de galería, terrenos de ganadería extensiva y zonas de vegetación arbustiva o hidrófila, donde se alimentan, descansan y duermen. En la Costa Caribe colombiana los hábitats más frecuentados corresponden a humedales en bacines inundables del sistema hidráulico (Ballesteros & Jorgenson, 2009). Su alimentación se basa en gramíneas y ciperáceas tiernas (Corriale, 2010). Estudios de isótopos estables de Carbono realizados en huesos de H. hydrochaeris de los Esteros del Iberá, Corrientes, han demostrado una alta variabilidad en su conducta forrajeadora facilitando su adaptación a los cambios en la vegetación (Corriale & Loponte, 2015).
El comportamiento de estos roedores se ve influenciado por factores como la geomorfología de los hábitats, el clima, la disponibilidad de recursos, el manejo antrópico de las poblaciones y otras actividades humanas. Generalmente se agrupan en manadas relativamente estables, de entre 5-30 individuos, caracterizadas por un sistema social estratificado. Esto depende de la presión de caza llegándose a presentar individuos aislados. Su comportamiento suele ser diurno, pero debido al mismo factor pueden volverse principalmente nocturnos. Se caracterizan por su territorialidad, pero esta condición puede variar en la época seca cuando pueden llegar a congregarse hasta más de 100 individuos de distintos grupos alrededor de los humedales remanentes (Carrascal et al., 2011; Herrera, 2013). Su reproducción se caracteriza por la prolificidad ya que se da durante todo el año con gestaciones de 150,6 días para Hydrochoerus hydrochaeris y 108 días para Hydrochoerus isthmius.Trapido (1949) reporta rangos de 119-126 días y 104-111 días respectivamente. Las camadas suelen ser de entre uno y ocho individuos con pesos neonatales de alrededor de 1500 g para H. hydrochaeris y de 1100 g para H. isthmius (Moreira et al., 2013). Cada hembra puede tener dos gestaciones anuales y las crías tienen una rápida tasa de crecimiento. La madurez sexual de las hembras ocurre alrededor del año y medio, mientras que los machos tardan de tres a cuatro semanas (Nogueira-Filho, 1996). Los depredadores en orden de importancia suelen ser los perros domésticos (Canis lupus familiaris), la boa (Boa constrictor), los cocodrilianos (Caiman crocodilus fuscus y Crocodylus acutus), el jaguar (Panthera onca), las águilas crestadas (Morphnus guianensis) y otras aves de rapiña, las cuales atacan principalmente a las crías y juveniles (Carrascal et al., 2011; Nogueira-Filho & Nogueira, 2013).
Estado de Conservación de Hydrochoerus isthmius. Las principales amenazas para la conservación biológica de H. isthmius están asociadas con la cacería comercial y de subsistencia, la destrucción y contaminación del hábitat, la deforestación y la modificación de los sistemas hídricos que alteran la dinámica natural de los humedales. De acuerdo con la Unión Internacional para la Conservación de la Naturaleza (UICN) en su revisión de las Listas Rojas, esta especie se cataloga con Datos Insuficientes (DD), mientras que H. hydrochaeris se cataloga como de Preocupación Menor (LC). El estado de conservación de los ponches en la región noroccidental de Colombia y en las zonas bajas de las cuencas Magdalena-Cauca evidencia poblaciones muy diezmadas y con altos índices de amenaza, representando un peligro de extinción local en las subregiones del Bajo Sinú, Costanera y Sabanas en el departamento de Córdoba (Cortés, 1993; Ballesteros & Jorgenson, 2009).
En distintas regiones de América del Sur múltiples comunidades indígenas y campesinas han utilizado históricamente a los roedores silvestres como fuente de proteína, principalmente a especies grandes como la guartinaja (Cuniculus paca) y a otras de la familia Caviidae, subfamilia Hydrochoerinae (Salcedo-Rivera et al., 2018). Su carne se considera saludable por el bajo contenido de colesterol y grasas saturadas, y por la presencia de altas cantidades de grasas poliinsaturadas (Nogueira-Filho & Nogueira, 2018). De igual forma, su piel es comercializada para marroquinería y su grasa suele utilizarse en medicina tradicional. La cacería y la intensificación comercial de los ponches en la zona Norte de Colombia se incrementan principalmente hacia la época seca del año, asociadas con la concentración de las poblaciones en los relictos de agua (Ojasti, 1991; De La Ossa-Lacayo & De La Ossa-V, 2015). Este fenómeno concuerda con la temporada previa a la Semana Santa en Colombia, celebrada en el mes de abril, época en la que el ponche es ícono gastronómico al igual que otras especies silvestres, ya que en 1784 el Vaticano instó a los católicos romanos a consumir su carne durante este periodo, en acto de abstinencia de la carne roja (Herrera & Barreto, 2013). A esta amenaza se le añade una alta mortalidad relacionada con la disminución de la calidad del hábitat, y la aparición de enfermedades parasitarias, infecciosas y nutricionales (Ballesteros & Jorgenson, 2009), lo que hace necesario profundizar en su conocimiento biológico, ecológico y zooarqueológico contribuyendo a su conservación.
Materiales y métodos
Área de estudio. La Depresión Momposina es un sistema fluvial anastomosado que se encuentra ubicado en el Caribe colombiano sobre el valle del Bajo río Magdalena en confluencia con los ríos San Jorge, Cauca y Cesar en jurisdicción de los departamentos de Córdoba, Sucre y Bolívar. Al sur limita con Tierra Alta, al norte con San Benito Abad, hacia el oriente con el río Cauca y al occidente con las sabanas del Caribe. Su formación se dio a partir del hundimiento progresivo de una cuenca transtensiva hacia el límite entre el Pleistoceno y el Holoceno, y en la actualidad se encuentra a 20 msnm. Su forma es cóncava y longitudinal y abarca una extensión de entre 380.000 y un millón de hectáreas (Van der Hammen, 1986; Herrera & Berrío, 1998; Rojas-Mora & Montejo-Gaitán, 2015). Debido a sus características geomorfológicas la región permanece inundada durante siete meses al año, entre abril y noviembre, a causa de un periodo de abundantes precipitaciones, siendo una de las más grandes áreas de inundación del mundo (HIMAT, 1977). La precipitación anual de la región oscila entre los 2000-3000 mm, con una temperatura media anual de 27,6ºC. Las mayores precipitaciones se presentan en los meses de junio, julio, agosto, septiembre y octubre, y los meses más secos suelen ser enero, febrero y marzo (Edit, 1983; Herrera et al., 2001). La Depresión Momposina consiste en un delta interior donde se concentra alrededor del 80% de las ciénagas de la región Caribe, al igual que diversos humedales como pantanos, zapales, caños menores y madres viejas (Plazas et al., 1979). Este sistema está rodeado de llanuras pantanosas de bosque de galería y sabanas donde habitan las poblaciones de H. isthmius (Jimeno & Reichel-Dolmatoff, 1990; Chacón et al., 2013) (Fig. 3).
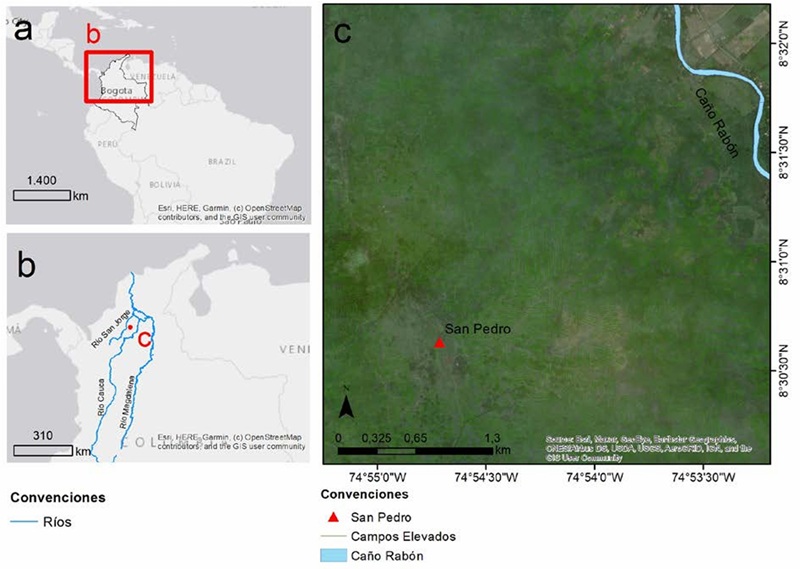
Figura 3 Localización del sitio arqueológico San Pedro en la Depresión Momposina, Colombia. Elaborado por la arqueóloga Ana María Aguirre.
Antecedentes Zooarqueológicos de la Depresión Momposina. La investigación arqueológica en esta región ha evidenciado la construcción de una obra de infraestructura hidráulica de más de 500.000 h., por las poblaciones humanas que habitaron la región entre los siglos IX a.C. a X d.C. Este sistema de canales y camellones o eras de cultivo permitía la conducción del agua en épocas de lluvia y su almacenamiento en épocas de sequía, lo que hizo posible el aprovechamiento anual de la tierra para la producción de alimentos basada en la agricultura, la pesca y la caza de mamíferos, aves, reptiles y anfibios mediante un manejo integral de los sistemas hidrobiológicos (Plazas & Falchetti, 1987; Plazas et al., 1988; Plazas et al., 1993; Rojas-Mora & Montejo-Gaitán, 1999; 2015; Rojas-Mora & Flórez-Correa, 2023).
Los canales sirvieron como vías de comunicación y transporte fluvial de alimentos y productos. Los poblados humanos se establecieron sobre plataformas de vivienda construidas a partir de la adición artificial de sedimentos por encima de las zonas inundables. También se han hallado áreas que se utilizaron para el cultivo extensivo. Una de ellas es el sitio San Pedro que se encuentra ubicado en el Municipio de San Benito Abad del departamento de Sucre, en la hacienda Tuloviste, a 30 km al sureste de San Marcos. El sitio tiene una extensión de más de 2000 h y al estar ubicado sobre las estribaciones del Cauca se encuentra a 20 m snm donde las inundaciones no son tan drásticas. Algunas fechas de radiocarbono derivadas de los basureros antropogénicos lo ubican en el siglo VII d.C. (Plazas et al., 1993; Rojas-Mora & Montejo-Gaitán, 2015; Flórez-Correa, 2023). En este lugar, el Programa de Investigación de la Depresión Momposina (PIDMO), adscrito a la Universidad de Antioquia (UdeA), ha realizado excavaciones arqueológicas encontrando abundantes restos de cerámica, carbón y arqueofauna compuesta por el ponche (H. isthmius), el tití cabeciblanco (Saguinus oedipus) tortugas (Trachemys venusta callirostris, Chelonoidis carbonarius y Podocnemys lewyana), cocodrilianos (Crocodylus acutus y Caiman crocodilus fuscus), peces (Pseudoplatystoma magdaleniatum, Prochilodus magdalenae, Ageneiosus pardalis, Caquetaia kraussii, Hoplias malabaricus y Synbranchus marmoratus) y gasterópodos terrestres (Marisa cornuarietis y Pomacea flagellata) (Flórez-Correa, 2018a; Flórez-Correa, 2023; Rojas-Mora & Flórez-Correa, 2023; Flórez-Correa et al., 2024).
Trabajo de Campo. La exploración arqueológica del antiguo poblado de San Pedro se llevó a cabo entre el 6 y el 25 de abril del año 2014. Allí se prospectaron tres plataformas de vivienda o unidades de exploración (U.E.) mediante la excavación de 115 unidades de recuperación (U.R.) o pozos de sondeo de 40x40 cm de ancho, por niveles arbitrarios de 10 cm, al igual que tres cortes de excavación estratigráfica con profundidades de hasta 2 m. En la Plataforma 1 se realizaron 61 pozos de sondeo (U.R. 1-61), y un corte (U.R. 100) con medidas de 6x1 m, dividido en tres cuadrículas de 2x1 m. En la Plataforma 2, 14 pozos (U.R. 62-75) y un corte (U.R. 103). En la Plataforma 3, 25 pozos (U.R.76-99 y 105) y dos cortes; la U.R. 101 compuesta por dos cuadrículas de 1 m2, y la U.R. 102 por una cuadrícula de 1 m2. Las acumulaciones de restos óseos y dentales fueron extraídas en bloques de sedimentos y rotuladas con códigos de contexto incluyendo el número de la plataforma (U.E.), la unidad de recuperación (U.R.), el número de cuadrícula y el nivel de profundidad en rangos de 10 cm.
Análisis de Laboratorio. La datación de la edad radiocarbónica del yacimiento se realizó a partir de muestras de carbón vegetal en el Laboratorio Beta Analytic. La calibración a años calendario se realizó con INTCAL13 empleando una curva de 0-50,000 años cal AP con 2 sigma. Las muestras de restos óseos fueron procesadas en el Laboratorio de Arqueobiología y Zooarqueología PALEO-ZOO en las instalaciones del Laboratorio de Arqueología de la UdeA realizando una limpieza con alcohol al 70% en atomizador y remoción mecánica de sedimentos con palillos de madera y pinceles sobre cernidores con luz de malla de 1 mm. Más adelante, se clasificó el material por unidad contextual, se individualizó cada fragmento con un código de secuencia y se ingresó la información de contexto en una base de datos de Excel.
Para la identificación anatómica y taxonómica de los restos de H. isthmius, se realizó un análisis de anatomía comparada con ejemplares (esqueletos completos) de la Colección Teriológica del Laboratorio de Mastozoología de la U. de A. Se revisaron las descripciones morfológicas disponibles del cráneo y el esqueleto axial (Macphee, 2011; Bode et al., 2013; Aeschbach et al., 2016), la pelvis y las extremidades posteriores (Araújo et al., 2013; García-Esponda & Candela, 2016; Brombini et al., 2018). Para la identificación de los restos de maxilar superior y mandíbula se registraron subvariables anatómicas como la superficie preservada del elemento (interna, externa o ambas superficies) y el número del alvéolo (1-4). En molares y premolares aislados se registraron variables como la posición del elemento (superior o inferior), el número del alvéolo de inserción (1-4) y el número de la lámina (1-11). En el análisis de los huesos largos se registró la porción conservada del elemento empleando las siguientes categorías: completo (C), epífisis proximal (EpX), epífisis proximal y diáfisis (EpX+Di), diáfisis (Di), diáfisis y epífisis distal (Di+EpT), y epífisis distal (EpT).
Se procedió con la cuantificación del Número de Especímenes Identificados por Taxón (NISP), el Mínimo Número de Elementos (MNE) y el Mínimo Número de Individuos (MNI) para cada depósito estratigráfico de acuerdo con Lyman (1994). Inicialmente se procedió con el conteo de cada uno de los fragmentos pertenecientes a H. isthmius (NISP), se establecieron conjuntos de fragmentos correspondientes al mismo elemento anatómico para establecer el MNE, se calculó su porcentaje multiplicando el valor obtenido en cada nivel de excavación por 100 dividido entre el total obtenido en cada U.R., y se estimó el MNI lateralizando los huesos largos de la extremidad inferior. Para la determinación de la edad relativa se evaluó el estado de desarrollo de los fémures a partir de la determinación del grado de fusión de la cabeza femoral de la epífisis proximal (EpX). Con este criterio se clasificaron los restos de acuerdo con el estado de maduración en categorías como juveniles (no fusionado), subadultos (parcialmente fusionado) y adultos (completamente fusionado) siguiendo las estimaciones de edad relativa propuestas para otros mamíferos silvestres (Lewall & Cowan, 1963; Carden & Hayden, 2015).
Se analizaron las alteraciones tafonómicas de los restos identificando los procesos que intervinieron en la modificación de los conjuntos óseos durante el procesamiento antrópico y la depositación y enterramiento de los restos. Se realizó una descripción de las marcas de corte según su frecuencia, dirección e intencionalidad, clasificadas como: en el hueso, cuando estuvieron destinadas a seccionar el elemento; o sobre el hueso, cuando su intención original fue procesar el tejido blando y se generaron marcas lineales en la superficie cortical. Se evaluaron los parámetros planteados por Shipman & Rose (1993) para identificar marcas de corte generadas por herramientas líticas. Para la determinación de fracturas en hueso fresco se aplicó el método propuesto por Alcántara-García et al. (2006), basado en la morfología del paño y ángulo de fractura.
Se estudiaron las alteraciones térmicas a partir de variables macroscópicas, tales como las variaciones cromáticas, textura, patrones de agrietamiento, las características de superficie, los grados de fragmentación de los elementos óseos empleando valores de 0%, 25%, 50%, 75% y 100%, y se determinaron los rangos de temperatura en °C de acuerdo con las categorías de Etxeberría (1994). Las alteraciones tafonómicas se registraron empleando un Estereomicroscopio Digital Brand NY con magnificaciones de entre 50x y 100x.
Se caracterizó la histología del tejido osteológico y odontológico en una muestra dental insertada en el alvéolo del maxilar y un fragmento de tibia evaluando el estado de preservación de las células óseas y dentales. Para esto se realizó un montaje en resina epóxica con un curado de 24 horas y postcurado de 4 horas a 60 °C, corte transversal con una cortadora de precisión IsoMet Buehler con disco de diamante, desbaste con papel abrasivo grado 600, 1000 y 1500, pulido en paño microcloth con alúmina de 0.3 μm con acabado de espejo, lavado en agua destilada y etanol, y secado con aire en el Centro de Investigación, Innovación y Desarrollo de Materiales (CIDEMAT) de la UdeA. Luego se analizaron en el Centro de Microscopía Avanzada de la UdeA donde se fijaron en cinta de grafito, se hizo un recubrimiento delgado en oro (Au) con el equipo Denton Vacuum Desk IV y se analizaron en Microscopía Electrónica de Barrido (MEB) en alto vacío (JEOL JSM 6490 LV) con el detector de electrones secundarios. Para la identificación de las estructuras histológicas se emplearon las convenciones del Cuadro 1, se evaluó el estado de alteración histológica (Prowse et al., 2004) y se aplicó el Índice Histológico de Oxford (IHO) para evaluar la degradación celular (Hedges et al., 1995; Millard, 2001).
Cuadro 1 Convenciones histológicas y celulares del tejido óseo y dental.
| Histología Ósea Histología Ósea | Histología Dental | ||
| Ha | Hueso alveolar (maxilar) | C | Cemento |
| Tc | Tejido compacto | E | Esmalte |
| Te | Tejido esponjoso o trabecular | D | Dentina |
| Cva | Cavidad vascular | Cd | Canalículos de la dentina |
| CH | Canal de Havers | Td | Túbulo dentinario |
| Cc | Canalículos calcóforos | Di | Dentina intertubular |
| L | Lamelas | Dp | Dentina peritubular |
| CV | Conducto de Volkmann | Pd | Predentina |
| Lo | Lagunas osteocíticas | P | Pulpa |
| Lcc | Líneas de cemento celular | Cp | Cámara Pulpar |
| G | Grietas | Cr | Conducto radicular |
| Os | Osteonas | Od | Odontoblastos |
| Li | Láminas intersticiales | Cc | Cuerpos celulares |
Se analizó la alteración nanoestructural de las fibras de colágeno y los cristales de hidroxiapatita en fragmentos de tibia y maxilar. Se trituraron las secciones de hueso en un macerador de ágata, se suspendió el material particulado en etanol grado analítico y se rompieron los aglomerados con ultrasonidos. Se tomaron alícuotas de cada suspensión y se colocaron en una rejilla de cobre Lacey/carbon 200 mesh para su análisis en Microscopio Electrónico de Transmisión (MET) Tecnai G2 F20. La composición química se determinó mediante la técnica de Espectroscopía de Rayos X de Energía Dispersiva (EDS-EDX) empleando una microsonda de referencia INCA PentaFETx3 Oxford Instruments.
Resultados
Abundancia relativa y representación anatómica de Hydrochoerus isthmius en yacimientos arqueológicos del sitio San Pedro. El conjunto arqueofaunístico recuperado en el Sitio San Pedro de la Depresión Momposina en el año 2014 está compuesto por tres clases de vertebrados: Actinopterygii (45,7%), Mammalia (22,2%) y Reptilia (19,9%). Estos porcentajes se basaron en el NISP de cada clase taxonómica. De la especie H. isthmius se identificaron 314 restos (MNE) que corresponden al 16,4% de la arqueofauna en general, y al 74,9% de los restos de mamíferos. En la Plataforma 1 se recuperaron 162 fragmentos de H. isthmius (51,5%), de los cuales el 80,1% corresponde a elementos craneales que incluyen incisivos (4,3%), molares y premolares (66,6%), maxilares (3,7%), mandíbulas (2,4%) y nasal (0,6%), al igual que un cráneo completo sin mandíbula (0,6%) compactado sobre una concreción sedimentaria. El 18,55% corresponde a huesos de las extremidades anterior y posterior que incluyen fragmentos de húmeros (2,4%), ulnas (0,6%), fémures (4,9%), tibias (2,4%), metapodiales (3%), falanges proximales (2,4%), falanges mediales (12%) falanges distales (1,2%), y huesos cortos como el calcáneo (1,2%) (Cuadro 2 y Fig. 4). En los depósitos I y II de la U.R. 100, fechados en 1390-1300 años A.P., anexos a la U.R. 26, se identificaron restos de tres individuos (MNI: 3) representados por dos fémures derechos de dos adultos (fusionados) (MNE: 2) y un fémur izquierdo de un juvenil (no fusionado) (MNE: 1) (Cuadro 3).
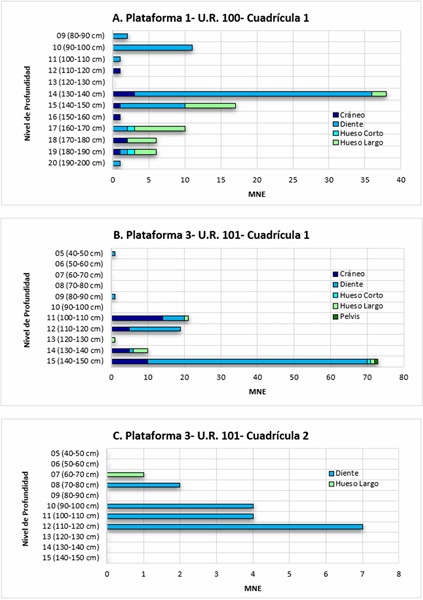
Figura 4 Abundancia relativa de las regiones anatómicas de H. isthmius en los niveles de profundidad de las excavaciones arqueológicas en el sitio San Pedro de la Depresión Momposina, Colombia. A) U.R. 100, Cuadrícula 1; B) U.R. 101, Cuadrícula 1; C) U.R. 101, Cuadrícula 2.
Cuadro 2 Cuantificación del Mínimo Número de Elementos Anatómicos (MNE) de H. isthmius hallados en las Unidades de Exploración (U.E.) del sitio arqueológico San Pedro de la Depresión Momposina, Colombia, incluyendo todos los pozos de sondeo y cortes de excavación (U.R.).
| Cuantificación Anatómica de los Restos Óseos de H. isthmius en el sitio arqueológico San Pedro | ||||||||||||
| Tipología y | U.E. 1 | U.E. 3 | MNE TOTAL | |||||||||
| Elemento | Bilateral | Derecho | Indet | Izquierdo | MNE | MNE% | Derecho | Indet | Izquierdo | MNE | MNE% | |
| Anatómico | ||||||||||||
| Cráneo | 2 | 7 | 2 | 4 | 15 | 9,2% | 14 | 3 | 17 | 34 | 22,3% | 49 |
| Completo | 1 | 0 | 0 | 0 | 1 | 0,6% | 0 | 0 | 0 | 0 | 0% | 1 |
| Mandíbula | 0 | 3 | 0 | 1 | 4 | 2,4% | 12 | 0 | 14 | 26 | 17,1% | 30 |
| Maxilar | 0 | 3 | 0 | 3 | 6 | 3,7% | 2 | 0 | 3 | 5 | 3,2% | 11 |
| Nasal | 1 | 0 | 0 | 0 | 1 | 0,6% | 0 | 0 | 0 | 0 | 0% | 1 |
| Temporal | 0 | 0 | 0 | 0 | 0 | 0% | 0 | 1 | 0 | 1 | 0,6% | 1 |
| Indeterminado | 0 | 1 | 2 | 0 | 3 | 1,8% | 0 | 2 | 0 | 2 | 1,3% | 5 |
| Dientes | 0 | 52 | 23 | 40 | 115 | 70,9% | 44 | 28 | 36 | 108 | 71% | 223 |
| Incisivos | 0 | 0 | 7 | 0 | 7 | 4,3% | 0 | 2 | 0 | 2 | 1,3% | 9 |
| Molares y premolares | 0 | 52 | 16 | 40 | 108 | 66,6% | 44 | 26 | 36 | 106 | 69,7% | 214 |
| Huesos Cortos | 0 | 1 | 1 | 0 | 2 | 1,2% | 0 | 0 | 1 | 1 | 0,6% | 3 |
| Calcáneo | 0 | 1 | 1 | 0 | 2 | 1,2% | 0 | 0 | 1 | 1 | 0,6% | 3 |
| Huesos Largos | 0 | 9 | 16 | 5 | 30 | 18,5% | 4 | 1 | 3 | 8 | 5,2% | 38 |
| Húmero | 0 | 2 | 0 | 2 | 4 | 2,4% | 0 | 0 | 0 | 0 | 0% | 4 |
| Ulna | 0 | 0 | 1 | 0 | 1 | 0,6 | 0 | 0 | 0 | 0 | 0% | 1 |
| Radio | 0 | 0 | 0 | 0 | 0 | 0% | 1 | 0 | 0 | 1 | 0,6% | 1 |
| Fémur | 0 | 4 | 3 | 1 | 8 | 4,9 | 3 | 0 | 3 | 6 | 3,9% | 14 |
| Tibia | 0 | 3 | 0 | 1 | 4 | 2,4 | 0 | 0 | 0 | 0 | 0% | 4 |
| Metapodial | 0 | 0 | 5 | 0 | 5 | 3% | 0 | 0 | 0 | 0 | 0% | 5 |
| Falange proximal | 0 | 0 | 4 | 0 | 4 | 2,4% | 0 | 0 | 0 | 0 | 0% | 4 |
| Falange medial | 0 | 0 | 2 | 0 | 2 | 1,2% | 0 | 1 | 0 | 1 | 0,6% | 3 |
| Falange distal | 0 | 0 | 1 | 1 | 2 | 1,2% | 0 | 0 | 0 | 0 | 0% | 2 |
| Pelvis | 0 | 0 | 0 | 0 | 0 | 0% | 0 | 0 | 1 | 1 | 0,6% | 1 |
| TOTAL | 2 | 69 | 41 | 49 | 162 | 100% | 62 | 32 | 58 | 152 | 100% | 314 |
Cuadro 3 Abundancia relativa de restos óseos y dentales de H. isthmius en las Unidades de Recuperación (U.R.) 100 (U.E. 1) y 101 (U.E. 3) del sitio arqueológico San Pedro de la Depresión Momposina, Colombia.
| Cuantificación de Restos Óseos y Dentales de H. isthmius por Unidad de Recuperación | |||||||||||||||||
| Nivel | Profundidad | U.E. 1 U.R. 100 | U.E 3 U.R. 101 | MNE | |||||||||||||
| General | |||||||||||||||||
| Cuadrícula 1 | Cuadrícula 1 | Cuadrícula 2 | |||||||||||||||
| Estrato | Depósito | MNE | MNE % | MNI | Estrato | Depósito | MNE | MNE % | MNI | Estrato | Depósito | MNE | MNE % | ||||
| 5 | 40-50 cm | 0 | 0% | 0 | 2 | III | 1 | 0,79% | 1 | 0 | 0% | 1 | |||||
| 6 | 50-60 cm | 0 | 0% | 0 | 0% | 0 | 0 | 0% | 0 | ||||||||
| 7 | 60-70 cm | 0 | 0% | 0 | 0% | 3 | II | 1 | 5,55% | 1 | |||||||
| 8 | 70-80 cm | 0 | 0% | 0 | 0% | 2 | 11,11% | 2 | |||||||||
| 9 | 80-90 cm | 8 | II | 2 | 2,12% | 3 | 3 | II | 1 | 0,79% | 1 | 4 | 0 | 0% | 3 | ||
| 10 | 90-100 cm | 11 | 11,7% | 4 | 0 | 0% | 0 | 5 | I | 4 | 22,22% | 15 | |||||
| 11 | 100-110 cm | 1 | 1,06% | 5 | I | 21 | 16,66% | 3 | 4 | 22,22% | 26 | ||||||
| 12 | 110-120 cm | 1 | 1,06% | 19 | 15,07% | 7 | 38,88% | 27 | |||||||||
| 13 | 120-130 cm | 0 | 0% | 1 | 0,79% | 0 | 0% | 1 | |||||||||
| 14 | 130-140 cm | 38 | 40,42% | 10 | 7,93% | 0 | 0% | 48 | |||||||||
| 15 | 140-150 cm | 17 | 18,08% | 73 | 57,93% | 0 | 0% | 90 | |||||||||
| 16 | 150-160 cm | 1 | 1,06% | 0 | 0% | 0 | 0 | 0% | 1 | ||||||||
| 17 | 160-170 cm | 9 | I | 10 | 10,63% | 0 | 0% | 0 | 0% | 10 | |||||||
| 18 | 170-180 cm | 6 | 6,38% | 0 | 0% | 0 | 0% | 6 | |||||||||
| 19 | 180-190 cm | 6 | 6,38% | 0 | 0% | 0 | 0% | 6 | |||||||||
| 20 | 190-200 cm | 1 | 1,06% | 0 | 0% | 0 | 0% | 1 | |||||||||
| Total general | - | - | 94 | 100% | 3 | - | - | 126 | 100% | 5 | - | - | 18 | 100% | 238 | ||
| MNE% general | - | 39,49% | - | - | - | - | 52,94% | - | - | - | - | 7,56% | 100% | ||||
Entre los restos identificados en la U.R. 100 se encontró un neurocráneo completo asociado a una tibia, ambos con excepcional integridad, que indican un descarte conjunto de ambas regiones anatómicas. El neurocráneo presenta en su vista dorsal: ancho del rostro de 62,18 mm, ancho zigomático en la raíz posterior de 76,67 mm, ancho parietal de 67,77 mm y ancho occipital de 72,06 mm. En la vista ventral: ancho de los forámenes incisivos de 6,65 mm, ancho del paladar entre los primeros premolares de 6,61 mm, ancho del paladar entre los últimos molares de 32,17 mm, longitud total de la serie dental de 83,5 mm, longitud basiesfenoide de 37,14 mm, ancho anterior del basiesfenoide de 14,19 mm, ancho posterior del basiesfenoide de 24,98 mm y amplitud de las bullas timpánicas de 56,48 mm. En su vista caudal presenta: diámetro horizontal del foramen magnum de 20,84 mm y vertical de 30,67 mm. La tibia presenta ancho máximo de la cara articular proximal de 37,21 mm, ancho de la fosa intercondilar proximal de 5,91 mm, profundidad metafisiaria proximal de 29,44 mm y ancho central de la diáfisis de 15,79 mm (Fig. 6).
En la Plataforma 3, se identificaron 152 fragmentos de H. isthmius (48,4%) de los cuales el 93,3% corresponde a elementos craneales que incluyen restos de incisivos (1,3%), molares y premolares (69,7%), maxilares (3,2%), mandíbulas (17,1%) y temporal (0,6%). A su vez, el 5,2% corresponde a huesos largos de las extremidades anterior y posterior que incluyen radio (0,6%), fémures (3,9%) y falange medial (0,6%), así como pelvis (0,6%) y calcáneo (0,6%) (Cuadro 2). En el depósito I de la U.R. 101, se identificaron como mínimo tres individuos (MNI: 3) representados por tres fémures izquierdos (MNE: 3), pertenecientes a un juvenil (no fusionado), un subadulto (parcialmente fusionado) y un adulto (fusionado). En el depósito II (101-2-7), fechado en 1350-1290 años A.P., se halló un fémur derecho fusionado de un adulto (MNI: 1) (Cuadro 3).
Fracturas en hueso fresco. A partir del análisis de las modificaciones antrópicas en las muestras se identificaron fracturas de tipo oblicuo-espiral en hueso fresco en el 25% de los restos óseos de la Plataforma 1 (U.R. 100) y en el 15.9% de la Plataforma 3 (U.R. 101). Se identificaron ocho fragmentos de mandíbula de las plataformas 1 y 3 con fracturas múltiples en hueso fresco, generalmente sobre el cuerpo mandibular cerca de los últimos molares y en la rama ascendente, comprometiendo la superficie articular con el neurocráneo. El desmembramiento de las extremidades consistió en la fracturación intencional de las superficies articulares de los huesos largos, de los cuales se hallaron fragmentos de húmero (MNE: 1) y cúbito (MNE: 1) fracturados cerca de la epífisis proximal, indicando la desarticulación de la cintura escapular y la extremidad anterior, así como 4 fragmentos de fémures (MNE: 4) con presencia de fracturas en espiral de paño oblicuo y longitudinal y de borde homogéneo, tanto en el cuello femoral como en la diáfisis, indicando desarticulación de la cintura pélvica y descuartizamiento por secciones. A su vez, se hallaron fracturas en hueso fresco en fragmentos de tibia (MNE: 1) y metatarso (MNE: 1) en la región distal (Fig. 5).
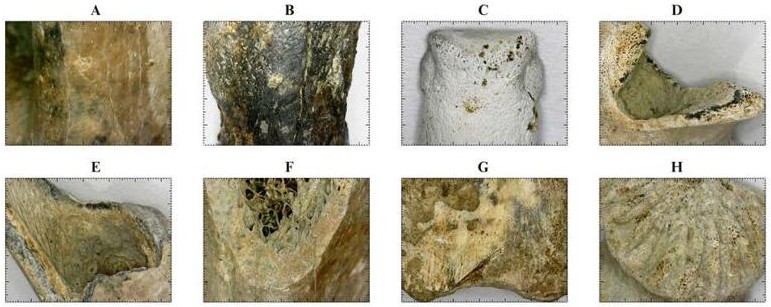
Figura 5 Modificaciones tafonómicas de origen antrópico (A-G) e indicadores del estado de desarrollo (H) en restos óseos de H. isthmius bajo Estereomicroscopio Brand NY (Escala: 1mm). A) Grietas longitudinales asociadas a termoalteración entre los 250 y 300°C en trocánter mayor de Fémur derecho [U.R. 26-20]; B) Falange medial parcialmente carbonizada entre los 300 y 350°C con grieta longitudinal en vista anterior [U.R. 59-13]; C) Falange medial calcinada a más de 650°C en vista anterior de la epífisis distal [U.R. 101-1-11]; D) Fractura en hueso fresco sobre rama ascendente de Mandíbula izquierda en vista caudal [U.R. 101-1-14]; E) Fractura en espiral sobre la diáfisis de Fémur izquierdo en vista craneal; F) Fractura en hueso fresco sobre Ilion de Pelvis derecha [U.R. 101-1-17]; G) Marca de corte diagonal sobre la epífisis proximal de Falange proximal en vista ventral [U.R. 100-1-17]; H) Estrías de fusión epifisiaria en cabeza de Fémur izquierdo de un individuo juvenil indicadoras del estado de desarrollo del espécimen capturado por los seres humanos [U.R. 101-1-1].
Marcas de corte en los restos óseos. Se identificaron marcas de corte sobre el hueso en el 8,5% de los restos óseos de la Plataforma 1 (U.R.100), encontrando cortes repetitivos y concentrados en el fémur sobre el trocánter mayor, en la falange proximal cerca de la epífisis proximal, y en fragmentos de mandíbula cerca de la rama ascendente y cortes repetitivos sobre la superficie ventral del cuerpo mandibular (Fig. 5).
Alteraciones térmicas. A partir del análisis de los restos óseos se identificó un patrón de exposición al fuego de todas las regiones anatómicas del esqueleto a temperaturas moderadas entre los 200 y 250ºC que se registra en el 80,8% de los restos óseos en la Plataforma 1 (U.R. 100) y en el 56,8% en la Plataforma 3 (U.R. 101), donde además se encontró que el 27,2% evidencia exposición entre los 250 y 300ºC. Esto indica que la preparación de la carne de ponche consistió en el asado directo de las presas sobre el fuego generando focos de alteración térmica en el tejido cortical del hueso (Fig. 5). En la Plataforma 3 (U.R. 101) se hallaron restos óseos con evidencia de sometimiento a altas temperaturas, en menor proporción, de los cuales el 2,2% presenta evidencias de carbonización entre los 300 y 350ºC, el 2,2% indica calcinación parcial entre los 550 y 600ºC, y el 2,2% calcinación total a temperaturas superiores a los 650ºC. En la Plataforma 1 (U.R. 100) se halló un 2,1% con señales de carbonización entre los 300 y 350ºC. Estas alteraciones probablemente se generaron en momentos posteriores al consumo, en contacto con fogones u hornos, o en quemas de desechos (Fig. 5).
Análisis de la diagénesis ósea en restos de cráneo y tibia asociados contextualmente. En general, los restos craneales presentan un alto grado de fragmentación, predominantemente entre el 75% y 100% en relación a procesos de quema y depositación. En la U.R. 100, nivel 18 (170-180 cm), se halló un neurocráneo completo compactado sobre una concreción sedimentaria, que presenta múltiples líneas de fragmentación postdeposicional sobre los ejes longitudinal y transversal y se encontró asociado contextualmente a una tibia izquierda que presenta grietas transversales a lo largo de la diáfisis (Fig. 6).

Figura 6 Neurocráneo completo y Tibia izquierda de H. isthmius [U.R. 100-1-18]. A) Neurocráneo en vista dorsal; B) Neurocráneo en vista lateral izquierda; C) Neurocráneo en vista ventral; D) Fragmentos craneales; E) Tibia izquierda en vista anterior o craneal.
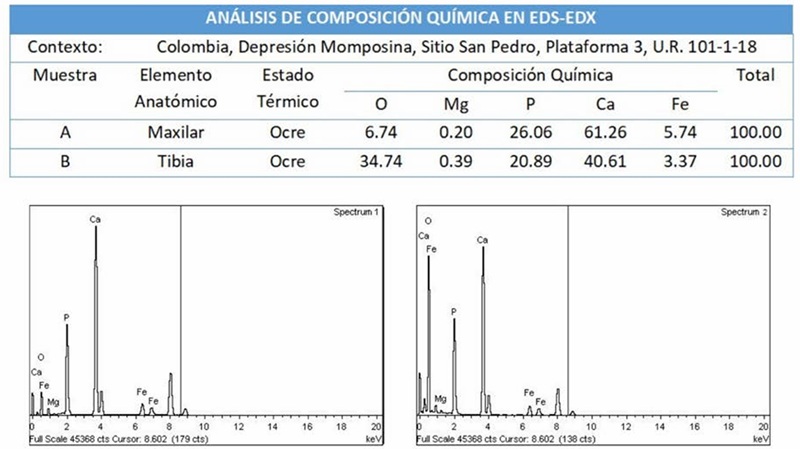
A partir del análisis histológico del tejido óseo del maxilar (Fig. 7 F-G) y la tibia (Fig. 7 H-I) en MEB se determinó un Estado B de alteración histológica (Prowse et al., 2004), con osteonas claramente visibles y pequeños focos de degradación. A su vez, se estimó un estado 3 según el índice de alteración histológica de Oxford con presencia de lagunas osteocíticas preservadas (Hedges et al., 1995; Millard, 2001). En la microestructura del maxilar se encontró un patrón de agrietamiento concéntrico alrededor de los sistemas osteónicos y sobre los ejes estructurales de las láminas intersticiales y de cemento celular (Fig. 7G). En la tibia se evidencia la incorporación de partículas sedimentarias, recristalización alrededor de los canales de Havers y ataque bacteriano alrededor de las osteonas (Fig. 7I). En el tejido odontológico se encontró una adecuada preservación de la pulpa, con capas bien diferenciadas y grietas radiales sobre el eje transversal de la dentina (Fig. 7 A-B-C), así como grietas transversales sobre el tejido trabecular del hueso alveolar (Fig. 7A-D-E). En el Cuadro 1 se presentan las convenciones empleadas en la identificación de las estructuras histológicas. El análisis en MET indica una orientación paralela de las fibras de colágeno con evidencias de alteración nanoestructural (Fig. 7J-Ñ). Los análisis en EDS-EDX del tejido óseo del maxilar y la tibia arrojaron una composición química de elementos como el Calcio (Ca) predominantemente, Fósforo (P), Oxígeno (O), Hierro (Fe) y Magnesio (Mg) (Fig. 8).
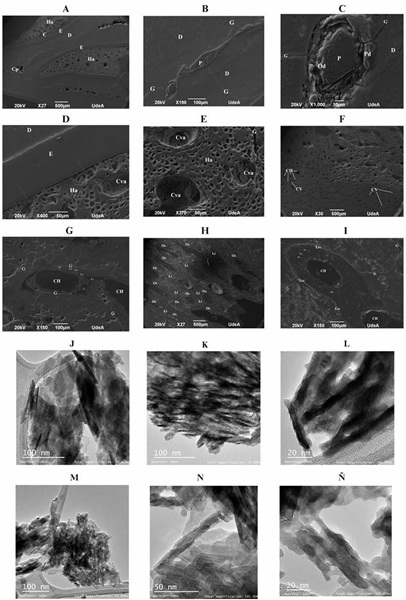
Figura 7 Magnificaciones histológicas de restos óseos y dentales de H. isthmius en Microscopía Electrónica de Barrido: (A-E) Premolar insertado en alvéolo [U.R. 101-2-7], (F-G) Maxilar [U.R. 101-1-15] y (H-I) Tibia izquierda [U.R. 100-1-18]. Alteración térmica de la nanoestructura de las fibras de colágeno visualizadas en Microscopía Electrónica de Transmisión: (M-Ñ) Maxilar [U.R. 100-1-18] y (J-L) Tibia izquierda [U.R. 100-1-18].
Discusión
El análisis de los restos óseos y dentales de H. isthmius recuperados por PIDMO-UdeA en el sitio arqueológico San Pedro de la Depresión Momposina, junto con las características contextuales del yacimiento y la presencia de evidencias tafonómicas del procesamiento antrópico, indican el aprovechamiento de esta especie por la población humana que habitó este antiguo poblado durante el siglo VII d.C. La gran variedad de ecosistemas hídricos asociados con la llanura de inundación del caño Rabón, constituyen el hábitat óptimo de este roedor acuático como señalan distintos estudios ecológicos en el Norte de Colombia, indicando una cacería local (Jorgenson, 1986; Ruiz, 1989; Ballesteros, 2001; Carrascal et al., 2007; González & Naranjo, 2007; Ballesteros & Jorgenson, 2009; Carrascal et al., 2011; Chacón et al., 2013).
La representación de unidades anatómicas y la abundancia relativa de H. isthmius en las secuencias estratigráficas de la U.R. 100 y la U.R. 101, señalan la preservación de todas las regiones esqueléticas en los yacimientos que se asocia con el transporte de los individuos completos desde las estaciones de caza hasta el asentamiento humano. La presencia de individuos juveniles y adultos está relacionada con una cacería aleatoria. Debido a la temprana madurez sexual y prolificidad característica de la especie, esta práctica pudo tener influencia en las tasas reproductivas de las poblaciones antiguas (Trapido, 1949; Nogueira, 1997; Moreira et al., 2013), según la densidad demográfica de las poblaciones humanas en distintos períodos de tiempo (Plazas et al., 1993). A su vez, la formación de los basureros arqueológicos del sitio San Pedro en la época seca (Flórez-Correa et al., 2024) se relaciona con el aprovechamiento estacional de la especie en una época de mayor vulnerabilidad ambiental (Ballesteros & Jorgenson, 2009). Esta información podría aportar claves para la conservación actual de H. isthmius en el Norte de Colombia al establecer una relación entre la estacionalidad hidroclimática y la cacería histórica de sus poblaciones. En ese sentido, las estrategias de conservación deben intensificarse en la época seca a partir de la divulgación científica de la ecología de esta especie con las comunidades locales y otros actores sociales relacionados con la conservación de los ponches.
Las evidencias tafonómicas del procesamiento antrópico están relacionadas con el descuartizamiento del cuerpo en secciones para su distribución y consumo entre los miembros del grupo humano, lo cual se infiere a partir de la presencia de fracturas en el esqueleto craneal y postcraneal, las cuales pudieron generarse a partir del impacto de cortes en el hueso y su posterior desprendimiento manual, produciendo la resistencia del tejido óseo fresco. De acuerdo con Blasco (1992), en la fracturación de los huesos frescos la distribución de la fuerza del impacto se da en el contenido orgánico mediante la absorción del golpe, haciendo que los límites de la resistencia del tejido cedan, permitiendo que el hueso recobre su estado original, dándose la ruptura a partir de una microfractura que se propaga a través de las ondas de percusión desde la zona del impacto hacia el exterior, siguiendo las líneas de debilidad del elemento anatómico. Este patrón fue observado en los restos óseos de ponche cerca de las zonas articulares, donde se identificaron fracturas de tipo oblicuo-espiral con paños longitudinales indicando su origen perimortem, que también podrían asociarse con la extracción de la médula ósea interna para su consumo.
Las marcas de corte superficiales identificadas en huesos largos y fragmentos mandibulares, caracterizadas por finas secciones en V, se asocian con el desprendimiento de los tejidos musculares empleando instrumentos de filo cortante. Según Jones (1980) los objetos líticos bifaciales son más útiles en las actividades de carnicería en contraste con los instrumentos retocados o sin retocar, mientras que Walker (1978) reconoce una mayor utilidad de los líticos retocados para cortar los tendones de las articulaciones. Sin embargo, los hallazgos de este tipo de artefactos en el sitio San Pedro son escasos, lo que impide hacer una asociación directa entre las marcas de corte y las herramientas que las generaron.
En un estudio etnozooarqueológico con cazadores de la Ciénaga de Ayapel, Baquero-Soto (2022) encontró que la forma de preparación de la carne de H. isthmius consiste en el hervido en agua y su posterior ahumado para garantizar su preservación. La presencia de restos óseos en el sitio San Pedro con alteraciones térmicas predominantemente entre los 200 y 300ºC en estado fresco, indican la exposición directa de las presas sobre el fuego. Las escasas evidencias de carbonización y calcinación a altas temperaturas son un indicativo de las formas de piro reducción posterior al consumo ya sea en el fuego de cocción o en pilas de desechos, tal como fue registrado por Flórez-Correa (2018b) en San Basilio de Palenque (Mahates, Bolívar) como una práctica habitual de quema de los restos óseos de vertebrados posterior a su aprovechamiento, que se asocia con el saneamiento de los desechos en descomposición.
La carbonización sucede tras la carbonatación de los componentes orgánicos como el colágeno y está asociada con la desaparición de carbonatos produciendo la coloración negra del tejido. A altas temperaturas, el dióxido de carbono (CO2) que se forma por la combustión de la materia orgánica con el oxígeno, genera la calcinación y la fusión de las sales del hueso (Correia, 1997). La exposición térmica genera cambios que comienzan con la deshidratación modificando la estructura ósea y generando un aumento en la microdureza que produce la disminución en la capacidad de absorción de energía del hueso, resultando en un material frágil de aspecto vidrioso susceptible al agrietamiento (Evans, 1973; Rootare & Craig, 1977) como se observó en los restos craneales quemados y carbonizados del sitio San Pedro que presentan una fragmentación cercana al 100%. De acuerdo con Baquero-Soto (2022), el desposte de H. isthmius se da a partir de un corte inicial entre el cráneo y las vértebras cervicales para la desarticulación de la cabeza, que posteriormente es desechada al igual que las zonas distales de las extremidades debido a su bajo contenido muscular. Esta práctica puede explicar el hallazgo de un neurocráneo completo asociado a una tibia en la U.R. 100, que es consecuente con la acción de arrojar estas unidades anatómicas tras su desarticulación en los basureros.
A partir del análisis histológico de estos elementos se encontró una adecuada preservación de las estructuras celulares (canales de Havers, conductos de Volckman, lagunas osteocíticas, lamelas concéntricas, líneas de cemento celular y láminas intersticiales) con focos puntuales de degradación. La preservación histológica depende del grado de descomposición y alteración de los compuestos orgánicos e inorgánicos del tejido óseo por procesos de bioerosión asociada a la actividad de microorganismos como hongos y bacterias en ambientes terrestres, al igual que la composición de los suelos y los regímenes hidrológicos que afectan el ambiente de depositación.
Tradicionalmente se ha asociado la bioerosión ósea a la acción de hongos y bacterias (Hackett 1981; Jans et al. 2004; Marchiafava et al. 1974) y se ha planteado que en los restos de fauna predomina el ataque fúngico sobre el bacteriano cuando estos se incorporan desarticulados y desprovistos de otros tejidos a la matriz sedimentaria (Jans et al., 2004). Sin embargo, no se han encontrado evidencias concluyentes de que los hongos realmente tengan la capacidad de destruir la microestructura ósea mediante el uso de sus componentes como fuente de nutrientes (Kendall et al., 2018).
La alteración encontrada en las células óseas de la tibia hallada en el sitio San Pedro indica una mayor actividad bacteriana que puede estar asociada a la descomposición de los tejidos blandos residuales posterior al descarte. Las bacterias capaces de producir bioerosión en el hueso son predominantemente aeróbicas procedentes del suelo (Kendall et al. 2018; Schotsmans et al. 2024; Turner-Walker, 2019; Turner-Walker et al. 2023). A su vez, la bioerosión está asociada a la pérdida de colágeno (Hedges, 2002), la incorporación de ADN bacteriano y/o fúngico (Hollund et al., 2012) y un aumento de la porosidad del hueso que conduce a una descomposición acelerada del tejido óseo (Gilbert et al., 2005). Estas condiciones tafonómicas de bioerosión bacteriana son consistentes con la formación de los depósitos arqueológicos en el Sitio San Pedro, generados a partir de la acumulación y descomposición de los desechos orgánicos producidos por la población humana.
Los datos de composición química obtenidos con EDS-EDX, arrojan valores esperables de O, Mg, Ca y P que son consistentes con los componentes de la matriz ósea, sin embargo, la presencia de Hierro (Fe) está asociada a su incorporación posterior a la depositación de los restos óseos en la matriz sedimentaria rica en óxidos de Hierro, junto con otras partículas sedimentarias visibles en las células óseas de la Tibia en MEB. De manera similar, López et al., (2011) encontraron Hierro mediante la técnica de EDS-EDX en restos óseos arqueofaunísticos de un sitio sumergido del Pleistoceno final de la costa central de Chile, e infirieron su incorporación en procesos diagenéticos, representando un ejemplo de interpretación que coincide con lo encontrado en los restos de tibia y maxilar de Ponche del sitio San Pedro.
Conclusiones
A partir del análisis de las evidencias óseas y dentales de H. isthmius halladas en el contexto arqueológico del sitio San Pedro, en la Depresión Momposina, Colombia, se puede concluir que la población humana que habitó este antiguo poblado, incorporó a la fauna vertebrada como una parte fundamental de su paleodieta, al interior de la cual, el ponche fue una especie importante dentro de su gastronomía como sucede actualmente en la región.
La cacería del ponche se dio en ecosistemas aledaños al sitio como canales artificiales, zapales, caños y madres viejas, principalmente en la época seca, a partir de su persecución y apresamiento. Los individuos capturados fueron transportados completos al asentamiento donde se dio su procesamiento por parte de los seres humanos, que consistió en la desarticulación del cuerpo y el descuartizamiento de los elementos anatómicos para su distribución y consumo, así como la extracción del tuétano, dando lugar a la fracturación intencional en hueso fresco cerca de las zonas articulares y la producción de marcas de corte sobre el hueso. Algunas regiones anatómicas de bajo contenido muscular como el cráneo y las regiones distales de las extremidades fueron descartadas, iniciando su proceso de descomposición bacteriana que generó la bioerosión de las estructuras celulares de las osteonas.
La preparación de los tejidos blandos del ponche se dio a partir del asado directo exponiendo las presas cerca del fuego, lo que generó alteraciones térmicas producidas por temperaturas de alrededor de 250°C con focos de carbonización parcial en las zonas más expuestas del hueso.
Posterior al consumo, se dio el desecho de los restos óseos y eventual quema, encontrando evidencias de carbonización y calcinación en algunos elementos óseos. Incluso los elementos descartados como el cráneo y la tibia presentan degradación térmica de la nanoestructura molecular del colágeno óseo. Más adelante, los restos orgánicos derivados de la alimentación humana fueron acumulados en pilas de desechos junto con piezas cerámicas fragmentadas, madera y semillas carbonizadas, que se compactaron y sedimentaron formando estratos arqueológicos caracterizados como basureros antrópicos, que registraron distintas ocupaciones humanas del sitio a través del tiempo, durante el Holoceno Tardío.

