











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El cáncer de mama es el tumor maligno más frecuente entre las mujeres; en 2020 afectó a 2.2 millones de mujeres y causó la muerte de 684 996 pacientes en el mundo y de 70 931 mujeres en México.1
Los anticuerpos terapéuticos son tratamientos eficaces con múltiples mecanismos de acción para eliminar células tumorales, entre los cuales se encuentra la citotoxicidad dependiente de complemento (CDC) que lleva a lisis celular.2,3 La sobreexpresión de las proteínas de membrana reguladoras del complemento (mCRP, membrane complement regulatory proteins), como CD46, CD55 y CD59, restringe la CDC en tumores y disminuye la eficacia de los anticuerpos terapéuticos.4-11
Generalidades del cáncer de mama
El cáncer de mama es una enfermedad heterogénea originada en las células epiteliales de la glándula mamaria. La clasificación histológica se basa en la anatomía de la glándula mamaria (carcinoma ductal y lobular) y se subdivide de acuerdo con la progresión (Figuras 1A y 1B). En el cáncer de mama se distinguen tres grados histológicos y cinco estadios clínico-patológicos que brindan información sobre el pronóstico de una paciente (Figuras 1A y 1B).12 El cáncer de mama suele clasificarse en cuatro subtipos caracterizados por la expresión de receptores: luminales A y B, HER2+ y triple negativo (Figura 1B). El tratamiento más adecuado se selecciona en función de la expresión de estos receptores, el grado y estadio tumoral.12
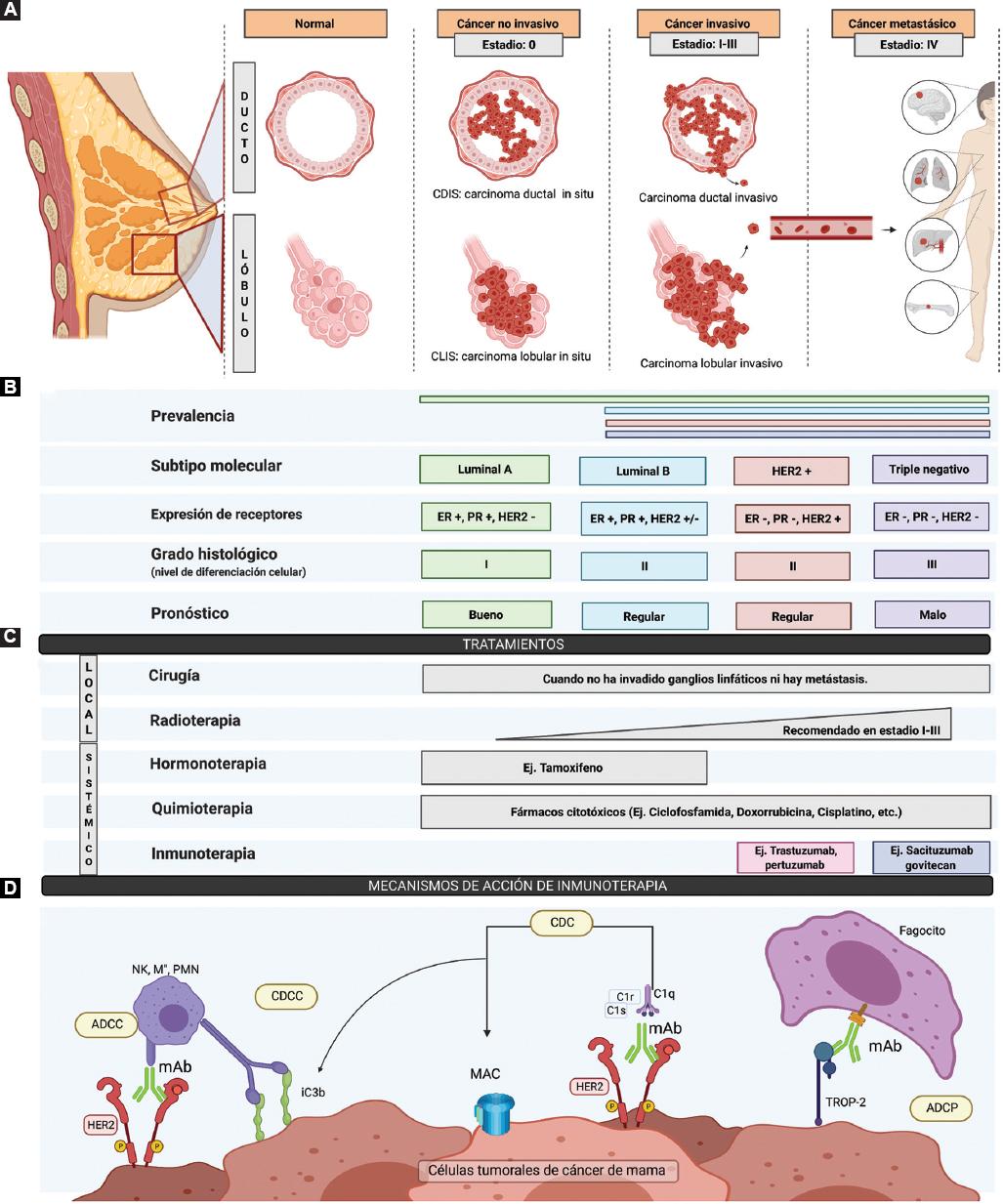
Figura 1 Patogénesis del cáncer de mama, histología, clasificación y tratamiento. A: la carcinogénesis se puede originar en las unidades ductolobulillares de las glándulas mamarias, dando lugar al carcinoma ductal o lobular. B: el carcinoma no invasivo comprende 25 % de los casos de cáncer de mama, de los cuales el subtipo más frecuente es el carcinoma ductal in situ (CDIS), con una incidencia de 80 %, seguido por el carcinoma lobular in situ (CLIS), cuya incidencia es de 20 %. El cáncer invasivo, que corresponde a 75 % de los casos, comprende de 5 a 15 % de los carcinomas lobulares invasivos y de 50 a 75 % de los carcinomas ductales invasivos. El cáncer de mama se clasifica de acuerdo con la presencia o ausencia de los receptores: los tipos luminal A (40 % de las pacientes) o B (20 % de las pacientes) expresan el RE y el RP; el tipo HER2 + (20 a 30 % pacientes) expresa el receptor 2 del factor de crecimiento epidérmico humano; y el tipo triple negativo (15 a 20 % de las pacientes) carece de la expresión de estos tres receptores. C: tanto los estadios como el grado de diferenciación proporcionan información para determinar el pronóstico y tratamiento de un paciente (local y sistémico). El cáncer de mama metastásico se trata según el subtipo, con el objetivo de prolongar la supervivencia global y paliar los síntomas (aproximadamente un año en el subtipo triple negativo y cinco años en los otros dos subtipos). D: mecanismos de acción de los anticuerpos que llevan a la eliminación de las células tumorales por complemento y células del sistema inmunológico. ER: receptor de estrógenos; PR: receptor de progesterona; HER2: receptor 2 del factor de crecimiento epidérmico humano; CDC: citotoxicidad dependiente de complemento; CDCC: citotoxicidad celular dependiente de complemento; ADCC: citotoxicidad celular dependiente de anticuerpos; ADCP: fagocitosis celular dependiente de anticuerpos. Creada con Biorender.com.
Tratamiento
El manejo terapéutico se basa en tratamientos locales: cirugía y radioterapia; y sistémicos: quimioterapia, hormonoterapia e inmunoterapia (Figura 1C).
Entre 20 y 30 % de los casos de cáncer de mama invasivo sobreexpresa HER2, una glucoproteína de transmembrana que regula el crecimiento, la supervivencia, la migración y la diferenciación celular.4,8,12 Trastuzumab y pertuzumab son tratamientos para pacientes HER+ (Figuras 1C y 1D); estos anticuerpos monoclonales (AcM) fueron aprobados desde 1998 y 2012, respectivamente. Ambos AcM (IgG1) reconocen distintos dominios extracelulares de HER2 y entre sus mecanismos de acción provocan la eliminación de células tumorales por diferentes mecanismos inmunológicos, incluyendo la CDC (Figura 1D).2-4 En comparación con la quimioterapia convencional, los AcM mejoran significativamente la supervivencia libre de enfermedad y la supervivencia global de las pacientes.13
Para el cáncer de mama subtipo triple negativo, en 2020 se aprobó un AcM conjugado con irinotecan, sacituzumab-govitecan. Este AcM humanizado (IgG1) reconoce una glucoproteína, conocida como TROP2 (antígeno 2 de superficie de trofoblastos) que promueve la migración y proliferación celular. La eficacia de este AcM depende de la citotoxicidad de irinotecan y de mecanismos de citotoxicidad celular dependiente de anticuerpos y CDC.14
La resistencia terapéutica a trastuzumab se ha observado en 40 a 60 % de las pacientes que sobreexpresan HER2;8 para pertuzumab se dispone de evidencia en modelos in vitro y murinos15,16 y para sacituzumab-govitecan aún no hay información debido a su reciente aprobación. Aun cuando no se conocen todos los mecanismos de resistencia a estos AcM, se ha propuesto que en parte se deben a la inhibición de la CDC por las mCRP.3,8,17
El sistema de complemento y las mCRP
El sistema del complemento es un mecanismo de la inmunidad innata que se activa en cascada para eliminar tumores. Funciona mediante tres vías convergentes: clásica, lectinas y alternativa.2-4 Trastuzumab, pertuzumab y sacituzumab-govitecan forman complejos inmunes de antígeno-anticuerpo que activan la vía clásica del sistema del complemento e inducen la lisis de células tumorales por CDC (Figura 2A). Sin embargo, la sobreexpresión de al menos una de las mCRP genera resistencia a la CDC y promueve la progresión tumoral.2,3,8,9
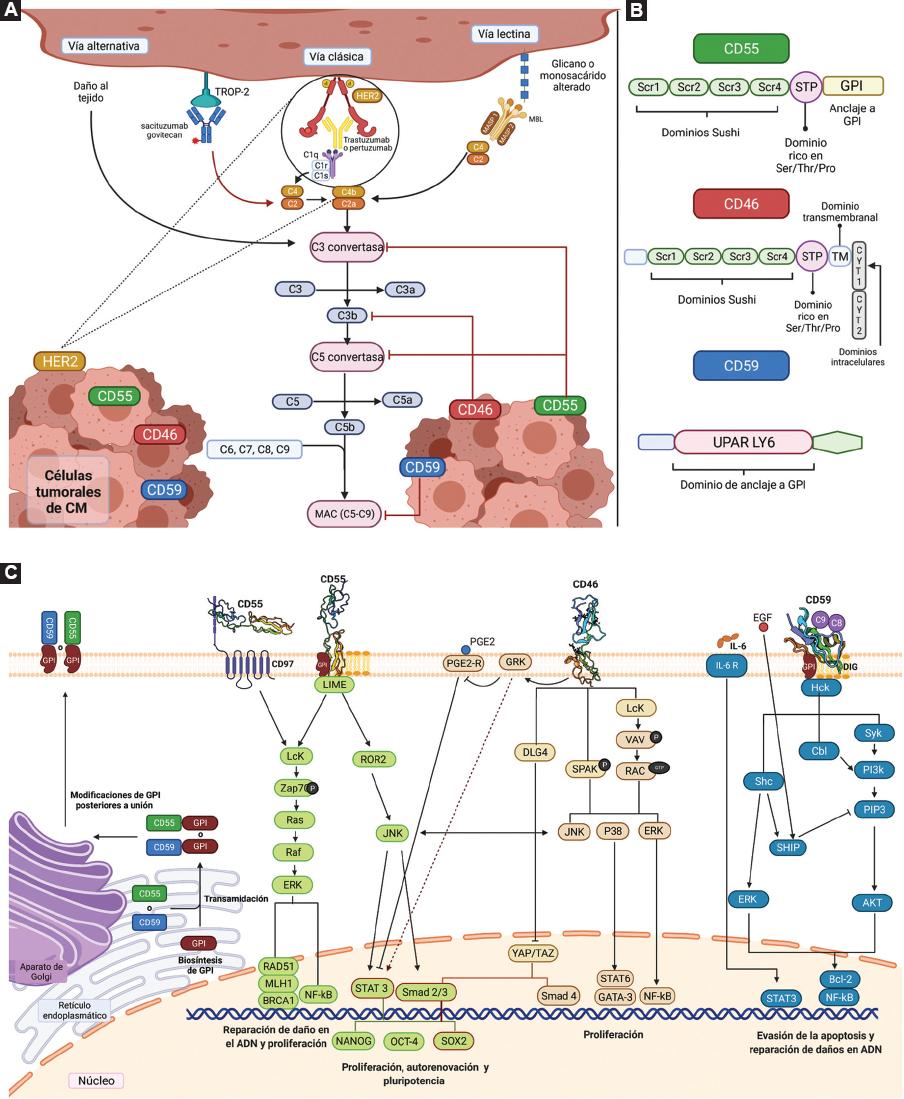
Figura 2 La expresión de mCRP en las células de cáncer de mama permite la supervivencia tumoral mediante la inhibición del sistema de complemento y la señalización intracelular. A: inhibición del sistema de complemento por las mCRP. Las células de cáncer de mama que expresan mCRP regulan negativamente la cascada del complemento inducida por la vía clásica, alternativa y de lectinas. CD55 evita la formación de las convertasas C3 y C5. CD46 favorece la degradación proteolítica de C3b y C4b y evita la formación de la C5 convertasa. CD59 actúa en la fase final de la cascada al unirse a C8 y C9 y evita la formación del complejo de ataque a la membrana (MAC). B: dominios estructurales de CD55, CD46 y CD59. Las proteínas CD55 y CD46 comparten una estructura similar al ser parte de la familia de reguladores del complemento (RCA), tienen cuatro dominios SCR, un dominio rico en serina, treonina y prolina (STP) y difieren en sus últimos dominios. CD55 tiene un dominio de anclaje a glicosilfosfatidilinositol (GPI) y CD46 tiene un dominio transmembranal y dos dominios CYT con propiedades de cinasas y proteasas. CD59 tiene un dominio funcional de la familia UPAR-LY6. C: modelo de las cascadas de señalización activadas por las mCRP. El anclaje a la membrana y la unión con los ligandos del complemento de las proteínas CD55, CD46 y CD59 inducen la activación de vías de señalización como AKT/PIP3 y MAPK, las cuales favorecen la reparación por daño en el ADN, la evasión de la apoptosis, la proliferación, la autorrenovación y la pluripotencia en células de cáncer de mama. CD55 y CD59 llegan a la membrana y se anclan a balsas lipídicas y DGI, mediante una unión con GPI inducida por una transamidación posterior a su biosíntesis en el retículo endoplasmático. La familia de proteínas p24 transfiere los complejos GPI-CD55 o GPI-CD59 al aparato de Golgi para modificaciones postranscripcionales de maduración que confieren la señal para exportarse a la membrana. STAT3 promueve la expresión de CD46. Cuando PGE2 se une a su receptor en la membrana de células tumorales, se disminuye la fosforilación de STAT3 y, consecuentemente, la expresión de CD46; para contrarrestar lo anterior, CD46 activa a las cinasas GRK, las cuales son capaces de mediar la activación de STAT3 al desensibilizar al receptor de PGE2. Proteínas cinasas: DLG4, Grk, Hck, JNK, Lck, ROR2, SPAK y Syk. Factores de transcripción: Bcl-2, BRCA1, GATA-3, MLH1, NANOG, NF-kB, OCT4, RAD51, Smad2, Smad3, Smad4, SOX2, STAT3 y STAT6. Vías activadas: AKT/PIP3, MAPK/ERK, MAPK/JNK y MAPK/P38. Creada con Biorender.com.
CD46
Es una glucoproteína transmembranal que inhibe la formación de la C5 convertasa del sistema del complemento, promoviendo la degradación de C3b y C4b mediante escisión proteolítica (Figura 2A).18 La transcripción de CD46 depende de la unión del factor de transcripción STAT3 a su promotor. Su estructura se basa en un dominio de repetición de consenso corto (SCR, short consensus repeats), una región de anclaje, un dominio transmembranal y una cola citoplásmica dividida en dos dominios: CYT1 y CYT2 (Figura 2B). Los estudios de mapeo bioquímico sugieren que los dominios SCR interaccionan con las proteínas del sistema del complemento para su inhibición.18,19
CD55
Es una proteína que acelera la disociación y desintegración de la convertasa C3 y previene la formación de la convertasa C5, evitando el daño celular (Figura 2A). Su expresión se regula a través del factor de transcripción Sp1.20 Estructuralmente, CD55 tiene un dominio extracelular que le permite interaccionar con las convertasas del sistema del complemento, una región espaciadora para determinar las posiciones de los dominios SCR y una región de anclaje con glicosilfosfatidilinositol que la une a la membrana celular (Figura 2B).21,22
CD59
Es una glucoproteína que inhibe la lisis celular y se une a la cadena α de C8 y al dominio β de C9 para evitar la formación del complejo de ataque a la membrana (Figura 2A).22 Pertenece a una superfamilia de proteínas con dominios Ly-6/uPAR que unen al sistema del complemento (Figura 2B).23 La expresión de CD59 se lleva a cabo bajo diferentes contextos:
– Regulada constitutivamente por el factor de transcripción Sp1.
– Inducida bajo condiciones inflamatorias por andamios entre las proteínas NFnnnnnnnnB y CREB unidas a CBP/p300.5
– Regulada condicionalmente por el factor de transcripción Smad3 inducido por TGF-nnnnnnnn durante la transición epitelial-mesenquimal.24
– Expresada selectivamente por el factor de transcripción SOX2 en poblaciones de células troncales tumorales.24
Señalización intracelular de las mCRP
La mayoría de las funciones reportadas para las mCRP están relacionadas con la inhibición del sistema del complemento (Figura 2A). Sin embargo, en diversas líneas celulares se ha identificado que la unión de las mCRP con sus respectivos ligandos (C4b, C3b, C8 o C9) y en coordinación con otras moléculas (TNF-α, IL-1β, IL-6, IFN-γ, E2 o EGF), también activan vías de señalización para regular procesos celulares (Figura 2C).3,6,23-25
Algunos estudios sugieren que la invasividad de las células de cáncer de mama depende de una cascada de señalización mediada por glicosilfosfatidilinositol unido a CD55 y CD97 mediante los dominios SCR.3,21 Tras la activación de CD55 por su ligando, las cinasas Lck y ROR2 se activan a través de un adaptador transmembranal llamado LIME.6 Lck regula una cascada río abajo por la activación de la vía MAPK/ERK, permitiendo la transcripción de genes involucrados en la reparación del ADN (MLH1, BRCA1 y RAD51) para evadir el proceso de apoptosis.6 Asimismo, por la vía de ERK se activa al factor de transcripción NFκB para favorecer la proliferación tumoral y la regulación positiva de las mCRP. Por otra parte, la activación de ROR2 conduce a la fosforilación de la cinasa JNK, favoreciendo la expresión de oncogenes característicos de poblaciones de células troncales tumorales responsables de la autorrenovación y pluripotencia (NANOG, SOX2 y OCT4)6 mediante las vías de STAT326 y Smad2/3 (Figura 2C).
En el cáncer de mama, CD59 activa la vía de ERK para regular su expresión a la alza,5,24 mientras que esta regulación es mediada por la vía IL-6R/STAT3 en las células de cáncer de páncreas (Figura 2C).26 La co-inmunoprecipitación de glicosilfosfatidilinositol en células THP1 y U937 ha permitido identificar que la activación de la tirosina cinasa Hck conduce a la fosforilación de la cinasa Syk y de las proteínas adaptadoras de señalización Cbl y Shc.23 Dado que CD59 se une a la membrana a través de la interacción con glicosilfosfatidilinositol y DGI,22,27 es probable la activación de estas proteínas en células tumorales. Las proteínas Syk y Cbl activan la transcripción de NFκB mediante la vía AKT/PI3k, mientras que la proteína de señalización Shc en sus formas p46 y p52 activa la vía MAPK/ERK para la activación de NFκB y Bcl-2, factores de transcripción que permiten la evasión de la apoptosis.23,27 Además, la activación de Shc promueve la fosforilación de la proteína SHIP, la cual es favorecida gracias a factores de crecimiento como EGF, con el fin de regular la activación de la vía AKT al inhibir a PIP3.23
CD46 induce una señalización intracelular mediante sus dominios CYT1 y CYT2, los cuales interaccionan con las cinasas DGL4, SPAK y Lck. Estas cinasas son capaces de activar las vías MAPK/ERK, MAPK/JNK y MAPK/p38. La cascada de fosforilación originada por estas cinasas permite la expresión de factores de transcripción (STAT6, GATA3 y NFκB), que favorecen la proliferación tumoral; asimismo, activa vías relacionadas con la adquisición del fenotipo troncal en el cáncer de mama.5 En contraste, la cinasa DGL4 lleva a una regulación negativa de las vías activadas por CD46 y CD55 al inhibir por fosforilación a los coactivadores transcripcionales YAP/TAZ, necesarios para la activación de los factores transcripcionales Smad4 y Smad2/3, que regulan la transcripción del oncogén SOX2 (Figura 2C). La vía CD46-GRK-PGE2 representa un bucle de regulación por realimentación que favorece la expresión de CD46 en las células tumorales (Figura 2C).28
Expresión de las mCRP y resistencia terapéutica en cáncer de mama
Se analizó la expresión génica y proteica de las mCRP en pacientes con cáncer de mama y fue comparada con la de tejido normal, para lo cual se emplearon las bases de datos TCGA (The Cancer Genome Atlas) y CPTAC (Clinical Proteomic Tumor Analysis Consortium) de UALCAN (The University of ALabama at Birmingham CANcer data analysis Portal). A nivel génico y proteico, CD59 y CD55 están subexpresadas en todos los subtipos histológicos. CD46 se expresa de forma diferencial entre los subtipos histológicos; se sobreexpresa de forma significativa en el subtipo luminal, mientras que se subexpresa en los subtipos HER2 y triple negativo. El nivel proteico de CD46 tiende a aumentar en todos los subtipos histológicos y es significativo en el subtipo luminal (Tabla 1). Diversos autores han reportado la sobreexpresión de las mCRP en muestras de pacientes y líneas celulares de los diferentes subtipos de cáncer de mama, que han sido usadas como modelos para estudiar la resistencia terapéutica a trastuzumab y pertuzumab.5,8,13,17
Tabla 1 Expresión génica y proteica de las mCRP e impacto en la supervivencia de pacientes con cáncer de mama
| CD55 | CD46 | CD59 | |||||
|---|---|---|---|---|---|---|---|
| Expresión génica (transcritos por millón)a | Luminal | ↓* (p = 0.012) | ↑* (p < 1 × 10−12) | ↓* (p < 1 × 10−12) | |||
| HER2+ | ↓ (p = 0.495) | ↓ (p = 0.903) | ↓* (p = 1.91 × 10−5) | ||||
| Triple negativo | ↓* (p = 0.002) | ↓* (p = 0.0005) | ↓* (p = 0.0003) | ||||
| Expresión proteica (valor Z)b | Luminal | ↓* (p = 0.032) | ↑* (p = 6.9 × 10−7) | ↓* (p = 0.0001) | |||
| HER2+ | ↓ (p = 0.620) | ↑ (p = 0.061) | ↓* (p = 0.021) | ||||
| Triple negativo | ↓* (p = 0.038) | ↑ (p = 0.268) | ↓* (p = 0.0023) | ||||
| Expresión de CD55 | Expresión de CD46 | Expresión de CD59 | |||||
| Baja | Alta | Baja | Alta | Baja | Alta | ||
| Supervivencia de pacientes con cáncer de mama (meses)c | Luminal | 148 | 81.48 | 108 | 91.93 | 90 | 111 |
| HR = 1.39, p= 0.054 | HR = 0.96, p= 0.805 | HR = 0.82, p= 0.245 | |||||
| HER2+ | 171.43 | 58.15 | 62.88 | 171.43 | 24 | 25.6 | |
| HR = 1.53, p= 0.124 | HR = 0.7, p= 0.196 | HR = 0.98, p= 0.933 | |||||
| Triple negativo | 35.42 | 27 | 30.42 | 28.09 | 30 | 29 | |
| HR = 1.34, p= 0.057 | HR = 1.07, p= 0.67 | HR = 0.97, p= 0.833 | |||||
aAnálisis de la expresión del gen de la base de datos TCGA (The Cancer Genome Atlas) en la plataforma UALCAN.
bAnálisis de la expresión proteica de la base de datos CPTAC (Clinical Proteomic Tumor Analysis Consortium) en la plataforma UALCAN.
cProbabilidad de supervivencia entre pacientes con diferentes subtipos de cáncer de mama y expresión de CD55, CD46 o CD59 (análisis log rank. Bases de datos: GEO, EGA y TCGA. Plataforma: Kaplan-Meier plotter).
*Significación estadística (p < 0.05) versus muestra de tejido normal (t de Student). ↑ aumento en la expresión; ↓ disminución en la expresión. HR: cociente de riesgo.
Con base en la plataforma Kaplan-Meier plotter y las bases de datos GEO, EGA y TCGA se identificó que no existe una relación significativa entre la expresión de las mCRP y la supervivencia de las pacientes, independientemente del subtipo de cáncer de mama (Tabla 1).
Un estudio retrospectivo mostró que aproximadamente 46 % de las pacientes con cáncer de mama HER2+ sobreexpresa de forma significativa las mCRP; sin embargo, solo la sobreexpresión de CD55 resultó ser un factor de riesgo y provocó una tasa de recaída más alta y una supervivencia libre de enfermedad más corta.13 De forma similar, en muestras de pacientes con neoplasias de mama negativas para CD46 se identificó un mayor tiempo de supervivencia global y supervivencia libre de enfermedad en comparación con pacientes con tumores positivos para CD46.9 Otro estudio reveló que la sobreexpresión de CD59 en pacientes con cáncer de mama se asocia a metástasis a pulmón y a una menor supervivencia libre de enfermedad.5
Estrategias para inhibir la expresión y función de las mCRP
Por su relevancia en la inmunoterapia y la progresión tumoral, se han propuesto y estudiado a las mCRP como dianas terapéuticas mediante diversos modelos y estrategias: siRNA,16,17,29 inhibidores farmacológicos,30-33 enzimas-péptidos8,10,34 y diversos AcM monoespecíficos y biespecíficos8,11,35-37 (Tabla 2).
Table 2 Enfoques dirigidos en el desarrollo de fármacos para regular la función de las mCRP y favorecer la eliminación de tumores
| Estrategia/tratamiento | Blanco | Efecto | Ventajas | Desventajas |
|---|---|---|---|---|
| siRNA anti-mCRP + inmunoterapia (trastuzumab)29 | CD55, CD46 y CD59 + HER2 | La inhibición de CD55 y CD59 causó la sensibilización de células de carcinoma de útero HER2+ a CDC y ADCC | Aumento en la eficacia de trastuzumab | La entrega del siRNA es inespecífica entre células normales y tumorales |
| siRNA anti-mCRP encapsulados en lipoplexos acoplados a transferrina16 | CD55, CD46 y CD59 | El silenciamiento de las tres mCRP condujo a un aumento significativo de la CDC y la opsonización | – La administración dirigida de siRNA anti-mCRP permite
la muerte de células tumorales sin afectar células sanas. – Eficiencia hasta de 90 % |
La entrega de los lipoplexos dirigida con transferrina depende de la presencia de CD71 en células tumorales |
| Atuplex™, sistema de administración de siRNA basado en lípidos catiónicos17 | CD55, CD46 y CD59 | Aumento de CDC, opsonización y apoptosis por caspasas en células tumorales de mama, ovario y pulmón HER2+ | Mejora de la actividad antitumoral del tratamiento con trastuzumab y pertuzumab | No descritas aún |
| Pequeña proteína recombinante Ad35K++10 | CD46 | La eliminación transitoria de CD46 sensibiliza a las células tumorales a CDC y ADCC | – Mejora la eficacia de trastuzumab, alemtuzumab y
rituximab. – Se comprobó seguridad y eficacia en ratones y macacos |
Existe una mínima posibilidad de que se produzcan anticuerpos contra la proteína Ad35K++ |
| Anticuerpo biespecífico anti-EpCAM* anti-CD5535 | CD55 y EpCAM | Aumento de 200 % en la deposición de C3 en células de carcinoma de cuello uterino que favorece la opsonización, la CDC y la ADCC in vitro | Aumento de la ADCC y CDC en células tumorales negativas para HLA de clase I | La efectividad del anticuerpo depende de una expresión elevada de los antígenos EpCAM y CD55 |
| Anticuerpo biespecífico anti-CD20,* anti-CD55 o anti-CD5911 | CD20*CD55 o CD20*CD59 | Aumento de la CDC de 4-25 veces mayor que con un anticuerpo dirigido únicamente a CD20 | Prevención tumoral completa en ratones | No descritas aún |
| AcM anti-idiotipo ONYVAX-10536 | CD55 | – Activación de células T y producción de citocinas.
– Menor recaída en pacientes respondedores. – Se favorece la lisis de células de cáncer colorrectal por CDC |
Se iniciaron los ensayos clínicos de fase II en mayo de 2000 | La fase clínica fue suspendida en 2007 y se inició el desarrollo de una segunda generación para entrar a fases clínicas |
| Conjugado de anticuerpo anti-CD46 con inhibidor de MMAF37 | CD46 y microtúbulos | – Aumento de la CDC e inhibición de proliferación de
células tumorales. – Inhibición del crecimiento tumoral en modelos de xenoinjerto ortometastásico |
Potencial para la enfermedad de alto riesgo y que tiene amplificación genómica en el locus del gen CD46 en pacientes con mieloma múltiple | No descritas aún |
| MFM-43 (AcM de bloqueo contra CD59)8 | CD59 | Aumento de la lisis celular de células SKBR3 y BT474 después del tratamiento con trastuzumab | Mejora de la eficacia de trastuzumab | No descritas aún |
| Fosfolipasa C específica de fosfatidilinositol8 | Fosfatidilinositol | – Provoca disminución de CD55 y CD59 en la membrana de
células tumorales de cáncer de mama (SK-BR-3 y BT474). – Aumenta la CDC inducida por trastuzumab. |
– Disminuye la resistencia de las células de cáncer de
mama a trastuzumab. – Proporciona una estrategia de tratamiento personalizada para pacientes con cáncer de mama HER2+ |
La entrega de la fosfolipasa es inespecífica entre células normales y tumorales |
| Metaloproteinasa ADAM1034 | CD46 | – La escisión y liberación de CD46 activa la apoptosis
de células tumorales a través de caspasas. – Se favorece la activación del sistema de complemento |
Estrategia para favorecer la destrucción de células tumorales por activación del complemento y apoptosis | El papel de la forma soluble de CD46 no ha sido estudiado a profundidad |
| Saracatinib.30,31 | Lck Src |
La inhibición de la reparación de daños en ADN provoca un aumento en la apoptosis de células tumorales de melanoma metastásico y cáncer de ovario. | – Finalizaron los ensayos clínicos de fase II con pocos
eventos adversos de grados 3 y 4. – Expresión río abajo de la vía de CD55 y CD46. – Aumenta la eficacia de los inhibidores de proteínas PARP |
Los eventos adversos incluyeron principalmente fatiga, hiperglucemia, anemia, linfopenia, síntomas gastrointestinales y aumento de las aminotransferasas |
ADAM10: desintegrina y metaloproteasa de matriz extracelular 10; C3: componente 3 del complemento; MMAF: inhibidor microtubular monometil auristatina F; PARP: poli-ADP-ribosa polimerasa; Lck: proteína tirosina cinasa específica de linfocitos; Src: protooncogén tirosina cinasa Src.
Mamidi et al. revelaron un aumento de CDC en líneas celulares de cáncer de mama y pulmón al usar un tratamiento combinado con trastuzumab, pertuzumab y lipoplexos catiónicos para la entrega de siRNA específicos para las mCRP.17 En otro estudio se usaron tres estrategias para aumentar la lisis celular mediada por trastuzumab en células de cáncer de mama: la primera estrategia consistió en el bloqueo de la función de CD55 y CD59 mediante el uso de anticuerpos monoclonales específicos; en la segunda estrategia se usó una fosfolipasa C específica de fosfatidilinositol (PI-PLC) que disminuyó la expresión de CD55 y CD59 en la membrana; en la tercera opción se usaron shCD59 y shCD55 para inhibir la expresión de dichas moléculas.8
Algunos inhibidores farmacológicos de las vías de señalización río abajo de las mCRP y de HER2 también se han evaluado como estrategias eficaces en el tratamiento del cáncer de mama. Saracatinib es un fármaco selectivo para las cinasas Src y Lck que ha mostrado un efecto sinérgico con un anticuerpo anti-ErbB2 en células resistentes a trastuzumab; dicho efecto se ha asociado a la inhibición de estas cinasas que funcionan como nodos de las vías de MAPK y PI3K.31
Lapatinib (inhibidor de la cinasa de HER2) y buparlisib (inhibidor de la cinasa PI3k) han demostrado eficacia en el tratamiento del cáncer de mama; dado que sus blancos terapéuticos comparten vías de señalización con las mCRP,32,33 cabe la posibilidad de aumentar la eficacia de los disponibles.
Discusión y conclusiones
Los AcM terapéuticos son una inmunoterapia eficaz contra el cáncer que cuentan con múltiples mecanismos de acción que incluyen la activación del sistema del complemento promoviendo la CDC.2,4,38 La expresión de las mCRP ha sido estudiada de forma extensa en cáncer, pero aún no se comprenden por completo los mecanismos que obstaculizan la eficacia de los AcM.3,4,8,13 Dado que los AcM anti-HER2 han estado más tiempo en uso que sacituzumab-govitecan, se ha identificado que la expresión de las mCRP permite a las células tumorales escapar de la actividad del sistema del complemento;5,8,13,15,17 sin embargo, se requieren más estudios para aclarar su función en la activación de cascadas de señalización.
El análisis de datos transcriptómicos y proteicos de muestras de pacientes con diferentes subtipos de cáncer de mama y su relación con la supervivencia de los pacientes, sugiere que no existe una relación directa entre la expresión de cada mCRP y la supervivencia. En contraste, algunos reportes indican que la sobreexpresión de las mCRP en pacientes con cáncer de mama se asocia a un peor pronóstico, resultado de una menor CDC inducida por anticuerpos monoclonales o lectinas.5,8,9,22 Estas diferencias se pueden explicar parcialmente, si consideramos que los niveles de las mCRP y su función podrían depender del estadio tumoral, tratamientos adyuvantes o alteraciones génicas y estructurales de estas proteínas. Por ejemplo, el tratamiento adyuvante con tamoxifeno en pacientes con cáncer de mama HER+ disminuye la expresión de CD55, con lo que mejora la lisis mediada por trastuzumab y resulta en un mejor pronóstico.13
Otro estudio identificó que la sobreexpresión de CD46 de tejido tumoral de pacientes con cáncer de mama tratadas con quimioterapia y hormonoterapia representa un factor de riesgo para la supervivencia general y libre de enfermedad.9 Se ha observado que en tumores de cáncer de mama, la sobreexpresión de CD46 correlaciona con el grado tumoral, tipo histológico y recidiva.39
La evidencia de que CD55 y CD59 en células troncales tumorales favorecen la tumorigénesis, proliferación, autorrenovación y resistencia a la muerte celular, sugiere que la función de estas moléculas no se limita a disminuir la eficacia terapéutica por inhibición del sistema del complemento, sino que pueden ocasionar resistencia terapéutica al regular subpoblaciones intrínsecamente resistentes, como las células troncales tumorales.6,24
Aquí resaltamos a las mCRP y a las moléculas involucradas en su señalización como blancos terapéuticos relevantes. No obstante, aún es necesario realizar más investigaciones para evaluar la seguridad y eficacia de las estrategias terapéuticas descritas (Tabla 2), así como evaluar su posible uso como tratamientos adyuvantes en pacientes con cáncer de mama.
Entender el papel de las mCRP en la resistencia terapéutica como un mecanismo que depende de la inhibición del sistema del complemento y de la transducción de señales en las células tumorales, proporciona una nueva perspectiva para identificar y desarrollar nuevas estrategias que aumenten la eficacia clínica de la inmunoterapia.

