












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
La vertiginosa pérdida de la diversidad ha llevado a que uno de los objetivos de la Agenda 2030 considere análisis detallados del estado actual de la diversidad de seres vivos y los factores que la afectan. La mayoría de estos estudios ha priorizado el conocimiento de plantas y vertebrados, sin considerar a los insectos (Carignan y Villard, 2002), pese a que representan la mayor proporción de la riqueza global de especies (Fernández, 2022). Dentro de estos, adquieren relevancia los parasitoides del orden Hymenoptera, ya que en términos de diversidad, son importantes por su elevado número de especies (Forbes et al., 2018), con 1 107 487 de especies estimadas a nivel mundial, y con poco más de 33,640 especies para el Neotropico (Fernández, 2022).
Además de su importancia en términos de riqueza, los parasitoides son importantes como enemigos naturales de insectos, ya que se desarrollan en su etapa larval dentro o sobre otros artrópodos, frecuentemente fitófagos, a los cuales, matan al final del proceso; por lo que también son importantes económicamente, al poder utilizarse en el control biológico de plagas (Godfray, 1994). Pese a la importancia del grupo, se desconocen los patrones de diversidad, ya que la mayoría de las investigaciones se enfocan a la interacción hospedero-parasitoide o al estudio de diversidad de alguna familia específica, por ejemplo, Braconidae, Chalcididae, Ichneumonidae, entre otras (González-Maldonado, 2020; Ramírez-Ahuja et al., 2020; Serrano-Domínguez et al., 2021; Jacobo-Macías et al., 2022; Villa-Ayala et al., 2022). Particularmente para la Península de Yucatán, se han desarrollado estudios sobre sistemática y diversidad de parasitoides (ej. González-Moreno et al., 2012; 2015a; 2015b; 2018; González-Moreno y Bordera, 2011; 2012; Chan-Canché et al., 2020; Cutis et al., 2022; Flota et al., 2024).
Algunos factores que pueden afectar la diversidad de insectos son la estructura y edad de la vegetación; se ha demostrado la relación inversa entre el estado sucesional del bosque y la diversidad de insectos, principalmente de organismos fitófagos (Díaz-Álvarez et al., 2023). En contraste, otros autores han demostrado que la diversidad de fitófagos es mayor en estados de sucesión avanzada (Byriel et al., 2023; Thomas, 2023). Además de estos resultados contrastantes, hay pocos estudios de cómo la sucesión de la vegetación afecta grandes grupos de insectos, como Diptera e Hymenoptera (Thomas, 2023).
Recientemente, se demostró que la edad de la vegetación, en niveles intermedios de sucesión, favorece una mayor diversidad de especies de parasitoides de la familia de Ichneumonidae, en comparación con selvas de sucesión temprana o con más de 60 años (González-Moreno et al., 2023); no obstante, se desconoce si estos patrones de diversidad que se reflejan a nivel de especie, también se presentan a nivel de familia. Por lo que en este trabajo se analizó la diversidad de familias de parasitoides del Orden Hymenoptera, en tres estados sucesionales de la selva mediana subcaducifolia. Se partió de la hipótesis que a nivel familia, los patrones de riqueza y diversidad asociados a estados sucesionales, se comportarán de manera diferente que a nivel de especie; ya que al analizar altos niveles taxonómicos (ej. familias), la riqueza y diversidad será mayor en la vegetación más vieja, debido a su estructura más compleja, lo que favorece mayor riqueza de fitófagos (Byriel et al., 2023; Thomas, 2023) y por consiguiente de parasitoides; así mismo, se espera que la composición de comunidades sea diferente para cada estado sucesional, principalmente por la familia que domine cada edad de vegetación. Esta información será relevante ya que genera no sólo conocimiento sobre la diversidad de parasitoides del bosque subcaducifolio, sino también, cuál es la composición de familias en cada estado sucesional de los ecosistemas.
Materiales y métodos
Área de estudio
El presente trabajo se realizó en la reserva ecológica biocultural Kaxil-Kiuic, localizada en el municipio de Oxkutzcab, Yucatán, entre los paralelos 20 º 21 ’ y 19 º 58 ’ N y 89 º 22 ’ y 89 º 46 ’ W a 33 msnm. El clima es cálido-subhúmedo, con lluvias en verano y un período de secas de noviembre a abril, con temperatura media anual de 26.3 °C y una precipitación pluvial media anual con rangos entre 1000 y 1100 mm. La humedad relativa en el mes de marzo es del 66 % y en diciembre de 89 %. Los vientos dominantes proceden del sureste. La flora se clasifica como selva mediana subcaducifolia con vegetación secundaria (Hernández-Stefanoni et. al., 2014).
Metodología
En la selva mediana subcaducifolia, se seleccionaron tres parches de vegetación con diferente estado sucesional: 5 a 9 años, 10 a 25 años, más de 60 años; los sitios fueron seleccionados considerando la caracterización estructural y de edad realizada por estudios previos (Hernández-Stefanoni et al., 2014). En cada sitio se colocaron, siguiendo un diseño completamente al azar, 10 trampas Malaise ampliamente utilizadas en programas de monitoreo de parasitoides (Chan-Canché et al., 2020). Las trampas funcionaron ininterrumpidamente durante la temporada de lluvias de agosto a diciembre de 2016, ya que son los meses en los que se han registrado picos de abundancia de parasitoides en la región (González-Moreno y Bordera, 2012; González-Moreno et al., 2015a; González-Moreno et al., 2018), con reposición de los botes colectores cada quince días.
Las muestras recolectadas en alcohol etílico al 70 % fueron llevadas al Laboratorio del Instituto Tecnológico de Conkal, donde fueron procesadas según las técnicas curatoriales convencionales. El material colectado fue identificado y clasificado a nivel de familia utilizando claves taxonómicas de Goulet y Huber (1993) y Gibson et al. (1997). Los ejemplares se depositaron en la colección entomológica del Instituto Tecnológico de Conkal.
Análisis de datos
Se realizaron curvas de acumulación, utilizando los estimadores no paramétricos de riqueza de especies el ACE y Chao 1, que consideran la abundancia de las muestras, para determinar el esfuerzo de muestreo (Magurran, 2004). La riqueza de familias de parasitoides entre las diferentes edades de la vegetación fue comparada con un análisis de rarefacción, con el software EcoSim700, con límites de confianza al 95 %. La diferencia en la abundancia de parasitoides entre los sitios se analizó con un análisis de varianza del número de individuos de las familias más abundantes, posteriormente se realizó una prueba de Tukey para determinar diferencias entre las medias de los grupos. De igual forma se analizaron las comunidades de parasitoides entre los diferentes sitios, construyendo curvas de Whittaker. Para calcular la diversidad alfa se utilizó el índice de Simpson, que se basa en el número de taxones dominantes. Para calcular la diversidad beta se hizo un análisis de similitud y se usaron diagramas de Venn para mostrar las especies compartidas entre sitios (Magurran, 2004).
Resultados y discusión
Se recolectaron 23 435 himenópteros parasitoides, representados por 9 superfamilias: Chalcidoidea, Chrysidoidea, Cynipoidea, Ceraphronoidea, Ichneumonoidea, Proctotrupoidea, Platygastroidea, Vespoidea, y Evanioidea. Las superfamilias más abundantes fueron Ichneumonoidea y Chalcidoidea con 7 422 (31.7 %) y 5 015 (21.4 %) ejemplares, respectivamente. Estos resultados son una sustancial contribución al conocimiento de la diversidad y composición de himenópteros parasitoides presentes en diferentes edades de vegetación de la selva mediada subcaducifolia. La superfamilia Ichneumonoidea, ha sido reportada comúnmente como abundante en otros estudios (Klopfstein et al., 2019; Chan-Canche et al., 2020; Polaszek y Vilhemsen, 2023), esto se debe a su gran número de especies, además de estar presente en diversos hábitats, por la variedad de estrategias de parasitoidismo y plasticidad que poseen (Quicke, 2015).
Riqueza de familias
La riqueza de familias fue de 33, coincidiendo con el valor estimado de la riqueza, basado en los estimadores no paramétricos Chao 1 (33) y ACE (33), lo que significa que se recolectó el 100 % de la riqueza total estimada para el área. Estos valores se alcanzaron a partir de la colecta seis, que equivale a tres meses de muestreo (Figura 1). El haber alcanzado el 100 % de las familias estimadas para el sitio de estudio, se debe a que con un muestreo extensivo como éste, con 30 trampas Malaise, se recolecte la totalidad de familias del sitio, sin llegar a la totalidad de especies.
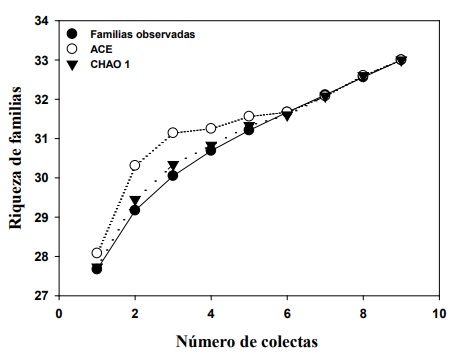
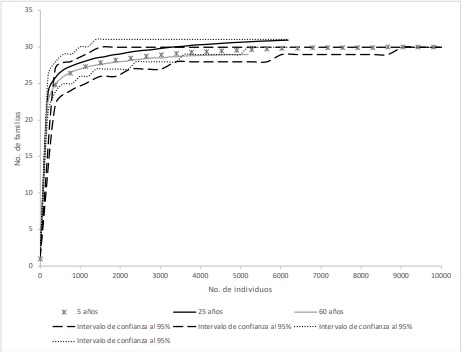
Con relación a la riqueza de familias se observó que no existen diferencias en el número de familias de parasitoides de los fragmentos de selva de diferente edad sucesional, de acuerdo con el análisis de rarefacción (Figura 2). El no haber diferencias probablemente se deba a que es altamente probable recolectar a ejemplares representantes de las familias, al ser éste un nivel taxonómico alto. Sin embargo, se recolectaron dos familias más, que las 31 familias registradas para la región (Chan-Canche et al., 2020).
Abundancia de familias
Las familias más abundantes fueron Braconidae, Scelionidae, Bethylidae y Diapridae (Figura 3), que son especialmente ricas en especies (Fernández, 2022), lo que aumenta la probabilidad de que sus especies puedan colonizar diferentes tipos de vegetación. Braconidae que destacó por su riqueza y abundancia, es la segunda familia del orden Hymenoptera con mayor riqueza y alberga géneros más grandes, principalmente con distribución cerca del ecuador (Quicke, 2015; Sharkey et al., 2023).
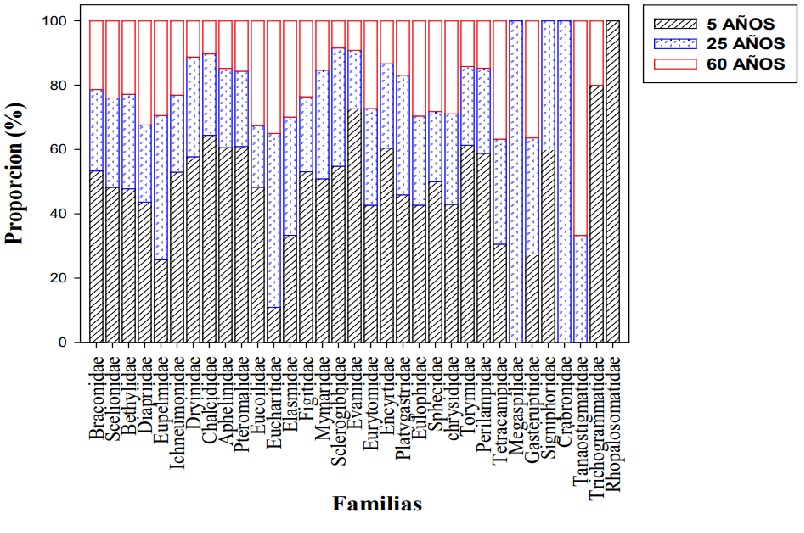
Con respecto a las otras familias más abundantes, también se caracterizan por el alto número de especies conocidas en el Neotrópico, con 3 000, 2 200 y 2 300 especies, respectivamente. Así mismo, Bethylidae es una familia gregaria (Fernández y Sharkey, 2006), por lo que es probable capturar altos números de individuos. Cabe resaltar que el 33 % de los Diapriidae estuvieron en la vegetación de 60 años, probablemente porque se encuentran conmunmente en habitats húmedos y áreas sombrías (Fernández y Sharkey, 2006), por lo que es probable que una selva con estructura de la vegetación más cerrada sea más atractiva para estos parasitoides.
La mayoría de las familias encontradas fueron más abundantes en la vegetación de 5 años, aunque sólo las familias Chalcididae, Braconidae y Pteromalidae fueron significativamente más abundantes en la vegetación de 5 años que en las otras edades de acuerdo a la comparación de medias (Tabla 1). Las familias Eucharitidae y Eupelmidae fueron más abundantes en un 54 % y 45 %, respectivamente, en la vegetación de 25 años; y en la vegetación de 60 años, la familia, Tanaostigmatidae, fue la única que tuvo el mayor número de individuos (Figura 3). También se registra por primera vez para Yucatán la familia Tetracampidae.
Tabla 1 Media de la abundancia de las familias con más de 500 individuos recolectados con trampa Malaise en diferentes edades de una selva mediana subcaducifolia, Yucatán, México.
Table 1: Mean abundance of families with 500 or more individuals collected with Malaise traps in dry semi-deciduous tropical forest fragments of different ages, Yucatan, Mexico.
| Familias Hymenoptera | 5-9 años | 10-25 años | > 60 años |
| Media | |||
| Braconidae | 315.20±0.25a | 14.80±0.10 b | 126.00±0.11b |
| Diapridae | 123.40 ±0.60 a | 67.80± 0.51b | 91.50±0.58 ab |
| Scelionidae | 132.00± 0.80a | 76.20± 0.71ab | 65.50±0.50 b |
| Bethylidae | 115.20±0.29a | 70.90± 0.22ab | 55.00±0.19 b |
| Eupelmidae | 27.50± 0.33b | 48.10± 0.61a | 31.50± 0.55ab |
| Aphelinidae | 47.20±0.57a | 19.20± 0.33ab | 11.50± 0.28b |
| Chalcididae | 49.60± 0.50a | 19.50± 0.32b | 7.90± 0.25b |
| Eucolidae | 32.40±0.95 a | 19.20±0.44 b | 21.90± 0.35ab |
| Pteromalidae | 39.00±0.87 a | 15.00±0.83 b | 10.00± 0.73b |
| Drynididae | 36.50±0.79a | 19.60± 0.61ab | 7.10± 0.55b |
Medias ± error estándar, letras diferentes en la misma fila indican diferencias significativas (Tukey P ≤ 0.05). Mean + standard error, different literals in the same row indicate significant differences (Tukey P < 0.05).
En este trabajo, la vegetación de 5 años fue la que presentó la mayor abundancia de varias familias de parasitoides, con relación a la vegetación de mayor edad. Esto coincide con lo registrado en la literatura, selvas en estados tempranos de sucesión, como los remanentes de 5 años de nuestros sitios, tendrán comunidades de parasitoides con altos valores de abundancia sólo para algunas familias, lo que resulta en comunidades poco diversas (Díaz-Álvarez et al., 2023; González-Moreno et al., 2023). Otro factor que pudo influir fue el tipo de trampa utilizada, ya que la trampa Malaise tiene una altura de captura que va desde el nivel de suelo hasta los 2 metros, por lo que es probable, que selvas con menor edad, tengan arboles más jóvenes, menos altos, y la altura en la que vuelan los parasitoides buscando sus hospederos, esté dentro del rango de captura de las Malaise, lo cual disminuye su eficiencia de captura en vegetación con mayor sucesión, con una estructura más cerrada y árboles más grandes, donde es probable que los parasitoides vuelen a mayor altura y queden fuera del rango de acción de la trampa. Además de que se ha comprobado que la altura en la que se coloca la Malaise, influye en la captura de algunas familias de parasitoides, tal es el caso de que las trampas colocadas a nivel del suelo favorecen la captura de Bethylidae, Diapriidae e Ichneumonidae (Chan-Canché et al., 2020), que fueron las familias más abundantes en este trabajo.
Estructura de comunidades
La estructura de comunidades de parasitoides fue similar en las tres edades sucesionales, sin embargo, considerando la pendiente de la curva, la vegetación de 25 años tiene mayor equidad que las otras edades de la vegetación; además cabe resaltar que tanto para la selva de 5 y 25 años, domina la familia, Braconidae; por el contrario en la edad sucecional de 60 años, además de Braconidae, Diapridae, Scelionidae y Bethylidae tienen rangos altos de abundancia (Figura 4).
Figura 4 Curvas de rango-abundancia de familias de parasitoides colectadas en tres estados sucecionales de la selva mediana subcaducifolia. Abundancia de las familias fue trazada sobre una escala logarítmica contra el rango de familias ordenadas por la abundancia, de mayor a menor. Los códigos de las familias más abundantes por estado sucesional: A. Braconidae, B. Diapridae, C. Scelionidae y D. Bethylidae.
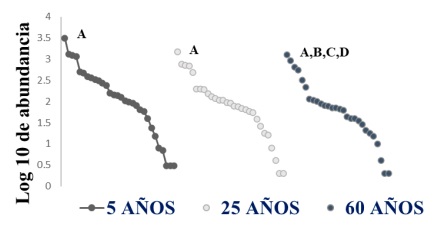
Figure 4: Rank-abundance plots of families of parasitoids collected from the three vegetation succesional stages in a dry semideciduos forest. A logarithmic scale of abundance was plotted against the species-rank ordered by families, from those with the most abundant individuals to those with the fewest. The codes of families most abundant by suceccional stage: A. Braconidae, B. Diapridae, C. Scelionidae y D. Bethylidae.
Diversidad alfa y beta
La diversidad alfa de familias de parasitoides fue mayor en la vegetación de 25 años (D = 9.31; 8.12, 8.60), con diferencias significativas, considerando los intervalos de confianza al 95 %, con la diversidad de la de vegetación sucesional de 5 años (D = 8.35; 8.12, 8.60) y 60 años (D = 8.17; 7.88, 8.47). Estas diferencias encontradas en las diferentes edades de la vegetación, coincide con varios autores que han demostrado que la edad de la vegetación puede afectar de forma positiva las poblaciones de herbívoros (Byriel et al., 2023; Díaz-Álvarez et al., 2023; Thomas, 2023) y dado que los parasitoides son densodependientes de sus hospederos, es de esperarse que también presenten el mismo patrón de afectación.
Con relación a la estructura de comunidades, la pendiente de la curva en la vegetación de 25 años sugiere una mayor equidad dentro de estas comunidades de parasitoides. Estos resultados son reforzados con los índices de diversidad alfa, que fueron más altos en esta vegetación con un estado de sucesión intermedia, comparada con la vegetación de edad temprana (5 años) o de estados sucesionales más avanzados (60 años). Estos resultados pueden explicarse con la Hipótesis de la perturbación media, que explica que la diversidad puede ser mayor en sitios donde la perturbación no es muy frecuente ni muy intensa, comparada con sitios no perturbados o con perturbación intensa (Speight et al., 2008). Se ha observado que los ecosistemas heterogéneos albergan más poblaciones de parasitoides reguladores de poblaciones de insectos fitófagos, pues existe una correlación positiva entre la diversidad de parasitoides y la diversidad vegetal (Wan et al., 2020; Guo et al., 2021; González-Moreno et al., 2023); y la arquitectónica o tipo de inflorescencias de las plantas (Fagundes et al., 2020; Zu et al., 2020).
La diversidad beta entre las diferentes edades fue baja, los valores de complementariedad alcanzan únicamente el 18% en las tres edades sucesionales, con 27 familias presentes en las tres edades de vegetación. Sin embargo, existieron algunas familias que se colectaron en dos edades de vegetación como fue el caso de Signiphoridae, colectada en 5 y 25 años, Tanaostigmatidae en 25 y 60 años y Trichogrammatidae 60 y 5 años. De la misma manera, se observó que algunas de las familias se colectaron sólo en una edad de vegetación, como es el caso de la familia Rhopalosomatidae que sólo se colectó en la vegetación de 5 años, y las familias Megaspilidae y Crabronidae que se colectaron únicamente en la vegetación de 25 años (Figura 5).
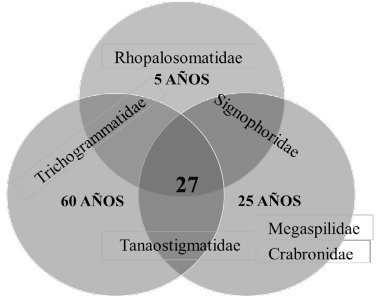
En cuanto a la diversidad beta, los altos números de familias compartidas puede deberse a que la mayor parte de las familias está bien representada en los tres sitios y al ser el taxón de familia un nivel taxonómico alto, no se pueden observar grandes diferencias en los ensambles; por lo que, si queremos ver diferencias en la composición de las comunidades es importante identificar los ejemplares a niveles taxonómicos más específicos como géneros y de ser posible a nivel de especie.
Las familias que fueron exclusivas para una edad de la vegetación, probablemente esté reflejando la presencia de sus hospederos, por ejemplo, Rhopalosomatidae encontrada en los fragmentos de 5 años, son ectoparasitoides de ninfas de Gryllidae (Orthoptera). Las familias Megaspilidae y Crabronidae colectadas en la vegetación de 25 años, son familias con un gran número de especies descritas y con amplios rangos hospederos, incluyendo no sólo insectos, sino también arañas (para el caso de Crabronidae), en general atacan diferentes especies de Coccinelidae, neurópteros, pupas de dípteros e hiperparasitoides de bracónidos, que fue una de las familias más abundantes. La familia Tanaostigmatidae fue la única que tuvo el mayor número de individuos en la vegetación de 60 años, sin encontrarse en la vegetación de 5 años, esto puede deberse a que varias especies de esta familia, no son parasitoides, sino fitófagos formadores de agallas (Goulet y Huber, 1993).
Conclusiones
En conclusión, la riqueza de himenópteros parasitoides en la selva mediana subcaducifolia está integrada por 33 familias, donde la familia Braconidae fue la más abundante. La edad de la vegetación es importante no sólo en términos de abundancia, sino también de diversidad, siendo la vegetación de 5 años, donde se encontró el mayor número de ejemplares de parasitoides y en la vegetación de 25 años se encontraron comunidades más equitativas y diversas de parasitoides. Estos resultados ponen de relieve la importancia biológica de las selvas medianas subcaducifolias de Yucatán como zona de refugio y preservación de parasitoides al albergar un alto número de familias conformando comunidades diversas.

