












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
Los ecosistemas forestales son uno de los principales sumideros de carbono terrestre (Ameray et al., 2021), además, son proveedores de bienes y servicios a escala local, regional y global (Von Döhren & Haase, 2015). Sin embargo, estos ecosistemas están constantemente bajo presiones antropogénicas y naturales que modifican su composición, estructura del dosel, en procesos y funciones que se desarrollan dentro de los bosques (Grantham et al., 2020; Patacca et al., 2023). Se han producido rápidas modificaciones en la estructura y composición de los bosques pasando de maduros a jóvenes debido a la sobrexplotación forestal y a incendios, sequias, plagas y enfermedades, etc... (Allen et al., 2010; O’Hara et al., 2017 Hammond et al., 2022). Asimismo, la distribución y la superficie de las áreas forestales ha disminuido por el aumento en la frecuencia e intensidad de las sequías provocadas por el cambio climático, lo que ha intensificado la mortalidad de las especies forestales en el mundo (Abbass et al., 2022). Por ende, la comunidad científica se ha centrado en entender la dinámica de los ecosistemas forestales y la transformación de su estructura bajo los efectos del cambio climático permite anticipar su capacidad de adaptación y resistencia (Muller-Landau et al., 2021; Hartmann et al., 2022; McDowell et al., 2022). Es importante para desarrollar estrategias que aseguren la sostenibilidad de los bosques y mitiguen los efectos del cambio climático, como la pérdida de carbono y la alteración de ciclos ecológicos.
Actividades como la productividad forestal está asociada a las limitaciones del crecimiento del arbolado, un factor clave en el almacenamiento de carbono que ayuda a mitigar los efectos del cambio climático (p.e. aumento de la frecuencia e intensidad de las sequías) (Girona et al., 2023). Unos de los temas que más ha captado la atención de los ecólogos es la relación entre el clima, la edad, especie, manejo y disturbios en el crecimiento radial de los árboles (Wilmking et al., 2020; Rydval et al., 2024), así como también la respuesta del crecimiento radial a la sequía en un contexto de cambio climático (Anderegg et al., 2019). Aun cuando el crecimiento forestal depende de múltiples factores ambientales y locales, la comunidad científica concuerda en que los anillos de los árboles son un proxy confiable para establecer estas relaciones clima-crecimiento (Babst et al., 2018; Wilmking et al., 2020). En este contexto, la resiliencia se define como la capacidad de un ecosistema para recuperar su estructura y función tras un evento de perturbación (Lloret et al., 2011). Por lo tanto, es fundamental profundizar en este tipo de estudios para comprender el proceso de adaptación de las especies forestales ante escenarios adversos (Kannenberg et al., 2020).
Las condiciones hidrológicas locales desempeñan un papel crucial a la hora de determinar la resiliencia al estrés causado por el calor y la sequía en los ecosistemas forestales (Song et al., 2022; B. Wang et al., 2022). La resiliencia en los árboles se refiere a la capacidad para mantener y recuperar una tasa de crecimiento estable después de una perturbación o un evento extremo (p.e. sequías, incendios o plagas) (Lloret et al., 2011; Marcotti et al., 2021). La resiliencia está influenciada por diversos factores, como las condiciones hidrológicas locales, sequias, la variabilidad genética, la biogeografía, el tamaño de los árboles y la competencia por los recursos (Castagneri et al., 2022; Locosselli et al., 2024). Es por lo que analizar los efectos de estos factores y su temporalidad es valioso para cuantificar puntos de crecimiento o decremento y determinar la persistencia de los bosques (Wang et al., 2022). También, los árboles que integran las diferentes comunidades tienden a crecer de forma independiente en respuesta al clima, lo que muestra un patrón del crecimiento que contribuye a la estabilidad y resiliencia generales de los bosques (Castruita-Esparza et al., 2019; Bernal et al., 2023; Correa-Díaz et al., 2023).
Existen evaluaciones que abordan la sensibilidad de la relación entre la respuesta al clima y el crecimiento (Acosta-Hernández et al., 2017; Carlón Allende et al., 2021). Sin embargo, nuestro conocimiento sobre como las especies forestales mexicanas responden a las variaciones y las condiciones climáticas locales, así como fenómenos a gran escala aún es limitado y requiere más investigación. Principalmente, las sequías extremas y los efectos acumulativos o legados afectan negativamente a los ecosistemas forestales, incidiendo más aún sobre aquellos ecosistemas especialmente vulnerables. Entre ellos, los Bosques de Alto Valor de Conservación (BAVC) son ricos en biodiversidad, constituyen el hábitat de especies en peligro, son refugios de fauna silvestre, y proporcionan servicios hidrológicos y de importancia social y cultural (Mikusiński et al., 2021; García-Trujillo et al., 2023).
Durango es la primera reserva forestal de México, donde destacan sus bosques de coníferas por ser ricos en biodiversidad con presencia de los géneros de Pinus, Quercus, Juniperus y Arbutus¸ entre otras hojosas (Martínez-Rivas et al., 2023); destacando las especies forestales de coníferas con distribución limitada como Pseudotsuga, Picea y Abies. La especie Abies durangensis Martínez tiene su distribución potencial centrada en grupos discontinuos en los estados de Chihuahua, Sinaloa, Jalisco y Durango (González-Elizondo et al., 2012). Es una conífera endémica de México, que forma parte de comunidades con Alto Valor de Conservación, puede llegar a medir hasta 40 metros de altura, tiene conos erectos con semillas en ramas laterales sobre un corto pedúnculo y las hojas son de color verde brillante de 20-35 mm de longitud por 1-1.5 mm de ancho (CONABIO, 2016).
A pesar de que en la actualidad en México se están abordando investigaciones relacionadas con la resiliencia a la sequía de las especies forestales (Cabral-Alemán et al., 2022; Correa-Díaz et al., 2023), aún es necesario un mayor esfuerzo para comprender la dinámica de la resiliencia de los bosques y especies arbóreas mexicanas ante eventos climáticos extremos. En particular, responder la pregunta: ¿Cómo ha respondido el crecimiento radial de A. durangensis a las sequías en el suroeste de la Sierra Madre Occidental?, y ¿Cuál es su capacidad de resiliencia ante estos eventos? Por tanto, el objetivo de este estudio fue evaluar la respuesta de los anillos de crecimiento radial a las condiciones climáticas y la resiliencia a los eventos de las sequias de A. durangensis en el suroeste de la Sierra Madre Occidental mediante proxies dendroecológicos. Se espera que A. durangensis presente una reducción significativa en el crecimiento radial durante los años de sequía, pero una alta resiliencia en las décadas más recientes debido a posibles adaptaciones a condiciones climáticas más extremas.
Materiales y métodos
Área de estudio
El área de estudio se ubica al suroeste del estado de Durango, en el macizo montañoso de la Sierra Madre Occidental (SMO) a 2800 m.s.n.m. (23.6° N, -105.3ºO; Figura 1).
Estos tipos de bosques se localizan principalmente en cañadas y laderas con exposición norte, cuya donde la orientación geográfica los protege de los fuertes vientos y la insolación intensa, formando un microclima húmedo y con temperaturas moderadas que favorece su adecuado crecimiento (González-Elizondo et al., 2012). En el sitio de estudio convergen especies de los géneros Abies, Pseudotsuga, Pinus, Juniperus y Quercus, los árboles pueden alcanzar alturas de hasta 50 m y supera los 70 cm de diámetro normal.
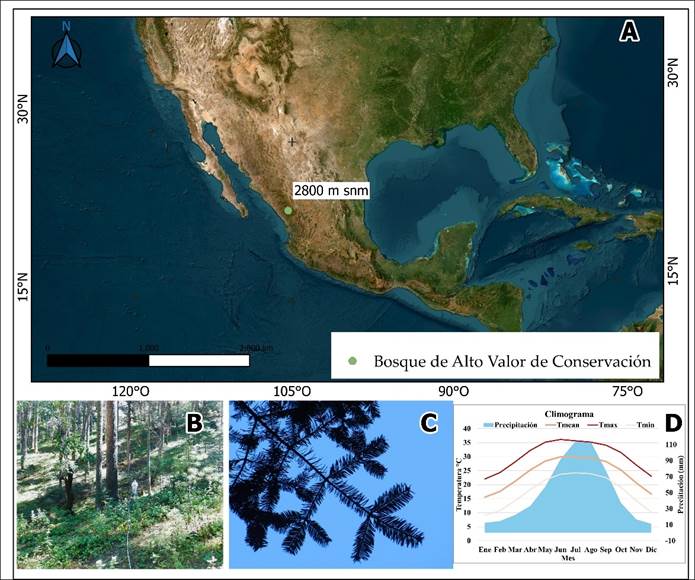
Figura 1 Ubicación geográfica del área de estudio (A), Vista general de área de estudio (B), vista del follaje de Abies durangensis Martínez (C) y Climograma (D).
El clima del área de estudio tiene temperaturas máximas de 22.03 °C en enero a 36.11 °C en junio, mientras que las mínimas oscilan de 9.05 °C y 24.25 °C. La precipitación sigue un patrón estacional, siendo julio el mes más húmedo con 114.25 mm, mientras que enero y diciembre son los más secos, con 12.38 mm y 11.17 mm, respectivamente (Figura 1). El tipio de suelo es Cambisol dístrico, y se caracteriza por un horizonte superficial rico en materia orgánica y buena fertilidad, con textura media (INEGI, 2017).
Colecta y procesamiento de datos dendrocronológicos
Fueron seleccionados 21 árboles de A. durangensis dominantes de edad desigual, contiguos a un árbol central elegido aleatoriamente, luego seleccionamos los 20 árboles adicionales ubicados más cerca del árbol centro (23.6°N, -105.3°O). Los árboles muestreados alrededor del árbol central incluyeron individuos adultos con diámetros mayores a 30 cm a una altura de 1.30 m. De cada individuo se colectaron dos núcleos de madera a una altura de 1.30 m utilizando una barrena de incremento de Pressler (5 mm de diámetro y 50 cm de longitud). Las muestras fueron montadas sobre bastidores de madera y secados a temperatura ambiente. Después se pulieron con papel lija de granulometría de gruesa a fina (100, 300, 600, 800 y 1000) hasta que fueran visibles los anillos de crecimiento (Stokes & Smiley, 1968).
Las series de crecimiento radial fueron datadas y sincronizadas mediante la identificación de años de crecimiento característicos, que son aquellos en los que los árboles muestran un crecimiento radial inusualmente rápido (anillo ancho) o reducido (anillos estrechos), generalmente en respuesta a condiciones climáticas extremas. El ancho total de anillo o TRW (Tree Ring Width, por sus siglas en ingles), fue medido sobre cada muestra utilizando el sistema de VELMEX el cual ofrece una alta precisión (0,001 micras) (VELMEX Inc. 2016, Bloomfield, NY, EE. UU.).
Luego del fechado visual y la medición de las series, se validó estadísticamente la datación cruzada mediante el programa estadístico COFECHA (Holmes, 1983). Los efectos biológicos y las tendencias geométricas de los árboles fueron eliminados mediante un modelo exponencial negativo como medio de estandarización. Este proceso se llevó a cabo utilizando la biblioteca dplR (Bunn, 2008) del programa estadístico R studio (R. Core Team, 2018), lo que asegura que los datos reflejan adecuadamente las variaciones en el crecimiento radial sin la influencia de estos factores. Las cronologías son consideradas registros que muestran cómo varía el crecimiento anual de los árboles con el tiempo, se generaron posterior a la estandarización a partir de las mediciones de los anillos de crecimiento (madera temprana, madera tardía y ancho total de anillo) para finalmente obtener los índices de ancho de anillo (RWI), índice que se caracteriza por su adimensionalidad, media cercana a 1,0 y varianza homogénea. Así mismo, fue calculada la señal poblacional expresada (EPS; Wigley et al., 1984) para el periodo 1871-2021 para determinar la idoneidad de las cronologías en términos de la señal poblacional (EPS > 0.85), las ventanas utilizadas en el análisis fueron 50 observaciones con un paso de 25. Esto implica que se evaluaron segmentos de datos de 50 años, avanzando 25 años en cada iteración.
Relaciones clima-crecimiento
Con el fin de identificar la influencia del clima sobre el crecimiento radial anual, se calcularon correlaciones de Pearson mediante la paquetería treeclim entre las variables climáticas mensuales y cronología del crecimiento radial. Se utilizó la plataforma Climate Explorer (https://climexp.knmi.nl/start.cgi, acceso el 20 de mayo del 2024; Trouet & Van Oldenborgh, 2013), a una resolución de 0.5° de la base de datos CRU TS v. 4.04 (Harris et al., 2020) para acceder a una serie temporal consistente y de calidad que abarca desde 1900-2022, lo que permite un análisis detallado de las tendencias climáticas. La información mensual descargada correspondió a las variables temperatura mínima (TMIN), temperatura máxima (TMAX) y precipitación (PP) dentro del periodo común de 1900 a 2022.
La influencia de la intensidad y duración de la sequía sobre el crecimiento radial fue evaluada mediante correlaciones de Pearson calculadas entre la corología y el Índice Estandarizado de Precipitación-Evapotranspiración (SPEI) (Vicente-Serrano et al., 2010; Beguería et al., 2014) fue el indicador empleado para representar la sequía. Este indicé basado en el balance hídrico y considera datos de precipitación y temperatura a diferentes escalas de tiempo. Los datos del SPEI fueron extraídos de la página oficial (https://spei.csic.es/, acceso el 20 de mayo del 2024), con una resolución espacial de 0.5° y una escala temporal de 1-48 meses para el periodo de crecimiento de 1954-2021. El SPEI es utilizado para monitorear sequías, basado en la relación entre la precipitación y la evapotranspiración, una herramienta útil para evaluar la disponibilidad hídrica en diferentes escalas temporales (Lee et al., 2024).
Índices de resiliencia
Se utilizó el paquete estadístico pointRes (van der Maaten-Theunissen et al., 2015) para analizar años característicos y los diferentes índices de resiliencia de las cronologías de crecimiento en R studio (R Core Team, 2018). Para identificar años característicos, identificados como crecimiento anormalmente ancho o estrecho, se procedió mediante el método de normalización de ventana móvil de Neuwirth (Neuwirth et al., 2007), el cual ajusta los datos de crecimiento a través de un análisis de ventanas móviles para detectar anomalías en el ancho de los anillos en comparación con las expectativas normales de crecimiento. Para conocer la capacidad de recuperar el crecimiento anterior a un disturbio, se utilizó el cálculo de los índices de resistencia, recuperación, resiliencia y periodo de recuperación de A. durangensis a la sequía (Lloret et al., 2011).
La resistencia mide la capacidad de la vegetación para mantener su funcionamiento (por ejemplo, crecimiento, producción de biomasa) durante una sequía. Se calcula como la proporción del estado del crecimiento durante la sequía en comparación con su estado antes de la sequía.
Donde:
La recuperación mide la capacidad del ecosistema para volver a su estado original después de que la sequía ha terminado. Se calcula como la proporción del estado del crecimiento después de la sequía en comparación con su estado durante la sequía.
Donde:
La resiliencia combina la resistencia y la recuperación, proporcionando una medida global de la capacidad del ecosistema para soportar y recuperarse de la sequía. Se calcula como la proporción del estado del crecimiento después de la sequía en comparación con su estado antes de la sequía.
Donde:
El periodo de recuperación representa el tiempo necesario para que el crecimiento alcance los niveles previos a la perturbación, lo cual es crucial para entender la dinámica de la respuesta del ecosistema a eventos estresantes como la sequía.
Resultados
Crecimiento de radial
Las mediciones de ancho total de anillo para A. durangensis mostraron un crecimiento promedio de 0.961 ± 0.251 mm/año con una amplitud o extensión de 151 años correspondientes al periodo 1870-2022, la intercorrelación entre series fue de 0.655, con una sensibilidad media de 0.274 mostró una autocorrelación de primer orden de 0.802 y se registró un EPS de 0.981, Estos resultados sugieren que A. durangensis presenta patrones de crecimiento particulares, los cuales se reflejan en años indicadores comunes (Figura 2). La variabilidad interanual del crecimiento puede ser evaluada analíticamente, considerando que los anillos de crecimiento pueden ser clasificados como anillos estrechos, asociados a condiciones de sequía, o anillos anchos relacionados con años húmedos. Por ejemplo, los años 2011, 2006, 1998, 1991, 1983, 1962, 1950, 1927, 1902, 1890 y 1885 son identificados como anillos estrechos, mientras que 2010, 2005, 2002, 1996, 1990, 1986, 1983, 1961, 1948, 1945 y 1941 corresponden a anillos anchos.
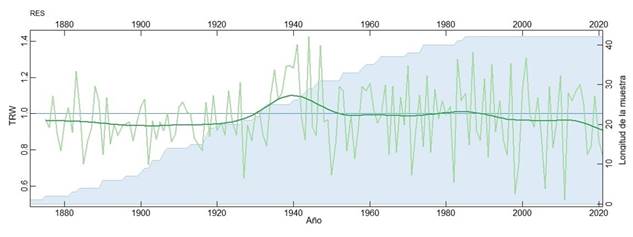
Figura 2 Cronología residual de índice de ancho de anillo (verde olivo) de A. durangensis. La línea verde oscura representa el spliline, mientras que el área azul indica el número de radios medidos (profundidad de la muestra) para cada año (eje y derecho).
Sin embargo, el índice de ancho de anillo de A. durangensis también revela que, en algunos periodos, como de 1940 a 2022, presenta una correlación más clara con los aumentos en la temperatura máxima (TMAX), sugiriendo que no solo la precipitación, sino también la temperatura, tiene un rol clave en la variabilidad del crecimiento radial. (Figura 2). Las variaciones de temperatura máxima y mínima también comienzan a tener un incremento a partir de 1940, mientras que la precipitación no mostró una tendencia temporal por lo que la disminución del índice de TRW tiene correspondencia con el incremento de temperatura (Figura 3).
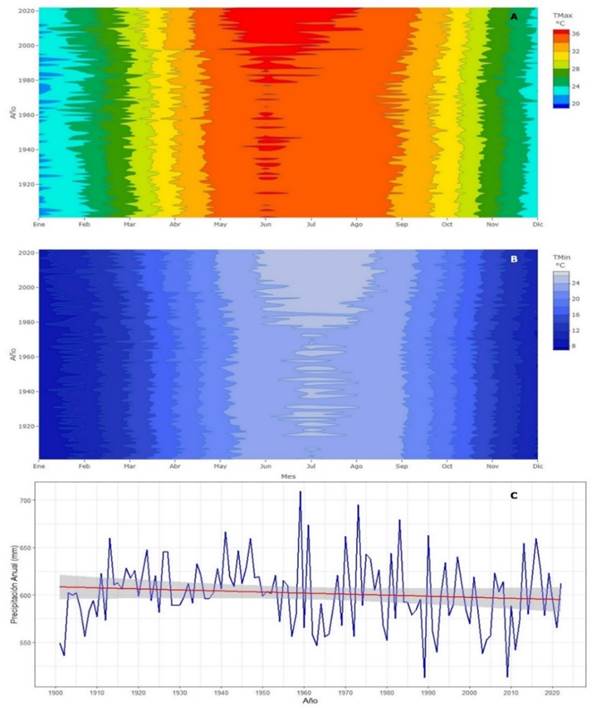
Correlación con clima
De acuerdo con los datos obtenidos de Climate Explorer, se puede observar que, las temperaturas máximas y mínimas promedio de cada mes, han comenzado a incrementarse a partir de 1980 (Figura 3). Caso contrario con lo que ocurre con la precipitación a partir de ese año, que no muestra una tendencia significativa en este periodo, sin embargo, ha comenzado a disminuir (Figura 3).
El análisis de correlación realizado mediante el paquete estadístico treeclim para el periodo 1900-2021 revelo la relación entre las variables climáticas y el crecimiento de los anillos de A. durangensis. Estos coeficientes indican la magnitud y el signo de la correlación entre las variables climáticas y el crecimiento de los anillos (Figura 4).
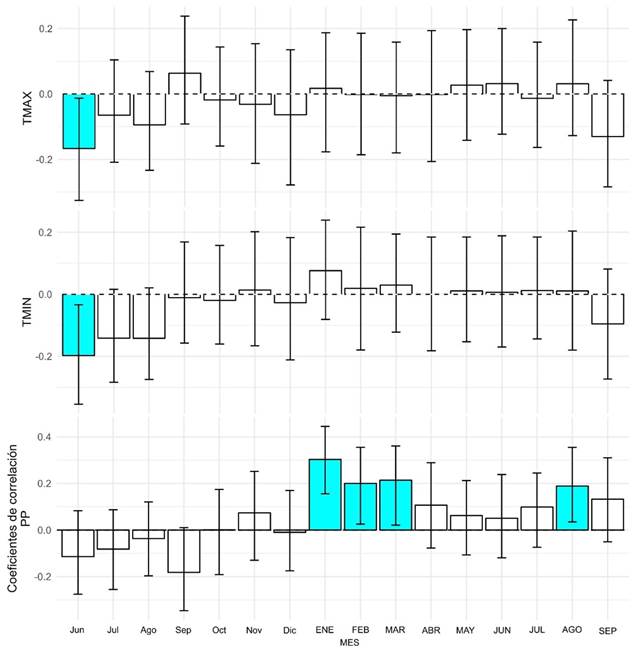
Figura 4 Correlaciones de la cronología del ancho de anillo con la temperatura máxima, la temperatura mínima y la precipitación (TMAX, TMIN y PP, respectivamente). Los meses en minúsculas corresponden al periodo de crecimiento anterior, mientras que los que están en mayúsculas representan el año actual. Las correlaciones que son estadísticamente significativas se destacan en cyan y se obtuvieron a un nivel de confianza del 95%.
Para la TMAX, el coeficiente de correlación para junio del año anterior es -0.172, lo que sugiere una correlación negativa significativa (p < 0.05) con el crecimiento del ancho de anillo. Esto indica que temperaturas máximas diarias mayores en junio del año anterior están asociadas con anillos de crecimiento más estrechos en el año siguiente.
En cuanto a la TMIN, en junio (-0.202) las temperaturas mínimas nocturnas más elevadas están asociadas con anillos de crecimiento más estrechos en el año siguiente (p < 0.05). Estos resultados sugieren que los patrones de temperatura mínima durante estos meses pueden tener un impacto significativo en el crecimiento de los anillos de A. durangensis. Por otro lado, para la PP se muestran correlaciones positivas significativas con el crecimiento de los anillos en enero, febrero, marzo y agosto del año siguiente con incrementos porcentuales del 30.6%, 20.1%, 20.9% y 19.1%, respectivamente. Esto sugiere que una mayor precipitación durante estos meses está asociada con anillos de crecimiento más anchos en el mismo año (Figura 4).
SPEI y TRW
El análisis de las correlaciones entre los meses del año del SPEI y TRW, revelan una fuerte relación entre la variabilidad climática y el crecimiento radial de los árboles. Las correlaciones de 0.39-0.60 indican que los patrones de crecimiento radial están estrechamente vinculados con el SPEI en diferentes periodos del año. Durante los meses de marzo, abril, mayo, junio, julio, agosto, septiembre, octubre, noviembre, y diciembre y la sequía acumulada desde 2 a 20 meses se observan correlaciones de 0.39 (Figura 5). Esto sugiere que el SPEI tiene un impacto significativo en el crecimiento radial de los árboles. En términos ecológicos, esto significa que la disponibilidad de agua es un factor crítico que influye en la capacidad de los árboles para crecer y desarrollarse. Un balance hídrico positivo, donde la precipitación es suficiente para satisfacer las demandas de evapotranspiración, se traduce en un aumento en el crecimiento radial.
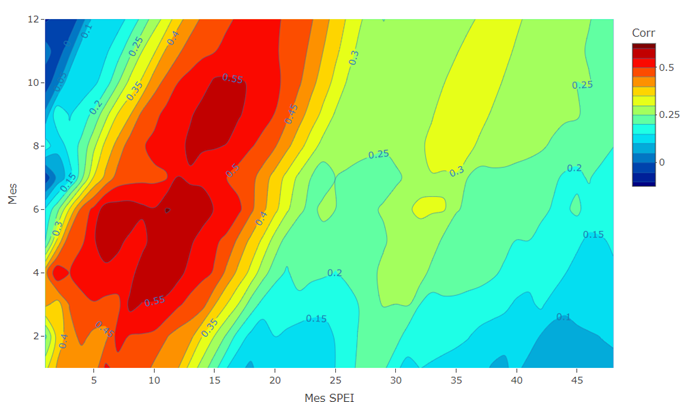
Figura 5 Correlaciones de los meses de crecimiento (enero-diciembre [1-12], eje y) en relación con la sequía y el crecimiento de A. durangenisis relacionando con TRW y el índice de sequía SPEI acumulado, considerando escalas de hasta 48 meses (eje x). Las correlaciones se calcularon para el período común y mejor replicado 1900-2021. Los coeficientes de correlación por encima de los valores de 0.30- 0.60 son significativos en P <0.05 y P <0.01, respectivamente.
Índices de Resiliencia
Los años característicos indica que los años extremos corresponden a eventos climáticos extremos, ya sean adversos o favorables y tienen un impacto significativo en el crecimiento de los anillos A. durangensis, ya sea de manera negativa o positiva. Los años característicos negativos extremos, como 1999, 1982, 1916, 1880 y 1879, sugieren sequías u otros factores ambientales negativos que resultaron en anillos más estrechos. En contraste, los años característicos positivos extremos registrados fueron 2015, 2001, 1987, 1970 y 1921. Los años característicos negativos fuertes fueron 2011, 2006, 1998, 1974, 1956, y 1987, mientras que solo 1881 fue positivo fuerte.
Aunque es probable que las sequías hayan sido un factor clave, es necesario considerar otros posibles factores ambientales negativos, como la competencia del suelo o perturbaciones biológicas, que podrían contribuir al desarrollo de anillos más estrechos (Figura 6).
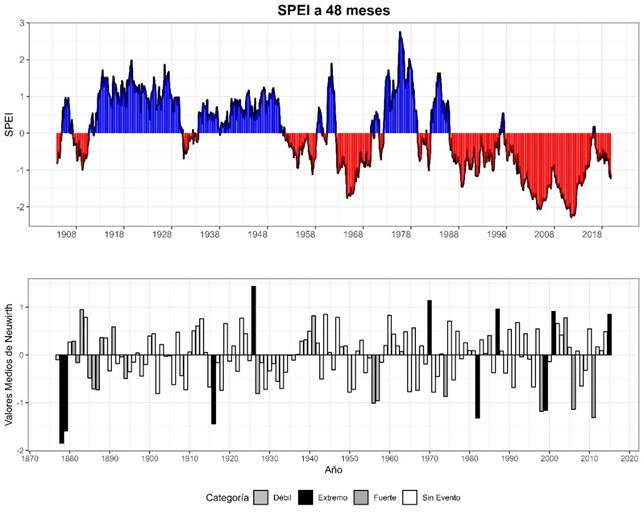
Figura 6 SPEI a 48 meses (superior) para el periodo 1900-2020 y valores medios de Neuwirth para el periodo 1878-2020 (inferior). Años característicos positivos y negativos mostrando aumentos y reducciones en el crecimiento radial (TRW, Tree-Ring Width), respectivamente.
En la Figura 7 se muestra la variación anual de los índices de resistencia, recuperación y resiliencia de TRW de A. durangensis para la extensión de la cronología de 1870-2021. A partir de 1990, los índices de recuperación, resistencia y resiliencia tienen una mayor variación que en los años anteriores, coincidiendo con una reducción del crecimiento medio de A. durangensis. Sin embargo, para los años identificados como característicos extremos negativos 1999, 1982 y los años negativos fuertes 1956, 1957, 1974, 1998, 2006 y 2011, éstos presentan registros más elevados de estos índices en comparación con los años no característicos. La resistencia registro los menores valores en los años 1982, 1998, 1999, 2006 y 2011. En cuanto a la recuperación, registró los mayores valores para esos mismos años. La resiliencia mostró variaciones a lo largo del periodo. En 1998, 1999, 2011 y 2012 se observaron valores de resiliencia con mayor frecuencia partir del fin del siglo XX. Se muestra que, las reducciones en el crecimiento son más irregulares e intensas, en donde se ve reflejado en la resistencia, la recuperación y resiliencia (Figura 7).
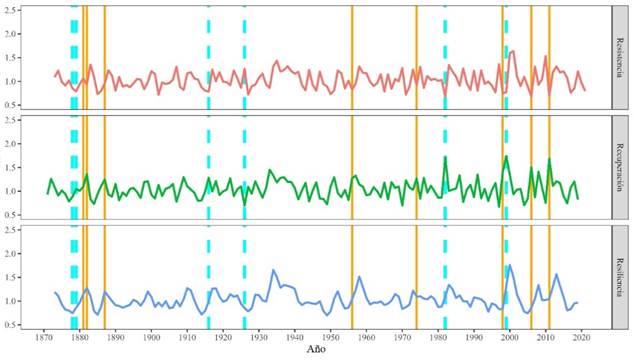
Figura 7 Índices de resistencia, recuperación y resiliencia para el periodo de 1870-2021. Años característicos negativos extremos: línea cyan y años característicos fuertes: líneas naranja.
El análisis del periodo de recuperación ha revelado una disminución significativa del tiempo de recuperación a lo largo del tiempo (pendiente = -0.0045, p = 0.0381), lo que indica que, en promedio, la especie está tardando menos en recuperar a los niveles de crecimiento pre-sequía (Figura 8). Este hallazgo parece contradictorio al considerar que la resistencia ha disminuido. Sin embargo, una posible explicación es que las condiciones post-evento, como la disponibilidad de agua y nutrientes, han mejorado en algunos casos, permitiendo que los árboles se recuperen más rápidamente. Alternativamente, podría haber un efecto compensatorio, donde la especie invierte más recursos en su recuperación a expensas de otros procesos, como el crecimiento a largo plazo.
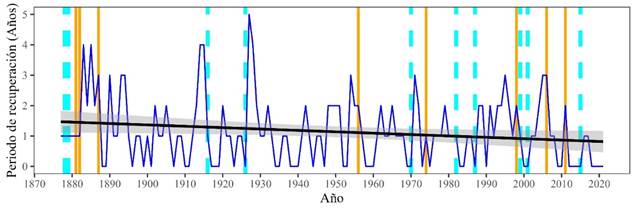
Figura 8 Tendencia del Periodo de recuperación de Abies durangensis de 1880-2021 (línea azul). Años característicos negativos extremos: línea cyan, años característicos negativos fuertes: líneas naranja, y línea de tendencia: línea negra.
A pesar de la disminución en el tiempo de recuperación, la estabilidad en los índices de resiliencia sugiere que A. durangensis mantiene una capacidad para recuperarse, pero sin lograr una adaptación superior en el tiempo. Esto resalta la complejidad de la dinámica de crecimiento post-sequía, en donde factores como el cambio climático, la disponibilidad de recursos y las perturbaciones forestales juegan roles interrelacionados. Aunque la especie puede recuperarse más rápidamente, su resistencia y resiliencia no han mejorado significativamente, lo que podría tener implicaciones importantes en la capacidad de A. durangensis para enfrentar futuros eventos extremos. Es crucial profundizar en el análisis de estos patrones para entender si el cambio en el tiempo de recuperación está vinculado a una mayor vulnerabilidad general del ecosistema o si responde a cambios en las dinámicas forestales a nivel local.
Discusión
La cronología evaluada muestra robustez, con un valor de señal poblacional expresada (EPS) de 0.981, similar a las reportadas en otros estudios (Villanueva-Díaz et al., 2016), indicando una alta fiabilidad en la serie cronológica. Con 42 núcleos y un total de 861 anillos, la correlación media (r-bar) fue de 0.546, lo que sugiere una adecuada consistencia entre las muestras.
Estudios anteriores han abordado la respuesta de crecimiento de los bosques a las condiciones climáticas, mostrando discrepancias entre los autores respecto al impacto de las señales climáticas por la edad. Mientras que algunos sugieren que la edad influye en dichas señales (Hadad et al., 2015; Jiao et al., 2017; Belokopytova et al., 2022), otros indican que las respuestas de crecimiento ante el clima son independientes de la edad (Linares et al., 2013). Esta divergencia sugiere que factores adicionales podrían estar modulando las respuestas del crecimiento de las especies arbóreas frente a las condiciones climáticas. Aunque no se evaluó directamente en este estudio, se reconoce que la edad puede ser un factor relevante en futuras investigaciones más específicos sobre el papel de la edad y otros factores en diferentes especies y regiones. Estos métodos no destructivos son una opción para evitar la remoción del arbolado, y necesarios para nuestra comprensión de la influencia de estos procesos naturales en el crecimiento de los árboles a condiciones extremas cada vez más recurrentes. Además, estos cálculos son vitales para sustentar propuestas de mitigación al cambio climático (Cabral-Alemán et al., 2022).
Esta investigación contribuye al posicionamiento de México en la agenda internacional sobre el tema de resiliencia de especies ante el cambio climático (Gazol et al., 2017; Schwarz et al., 2020; Song et al., 2022). Los resultados de esta investigación comprenden 151 años de resiliencia a la sequía en A. durangensis al suroeste de la SMO de Durango. En términos de vulnerabilidad climática, encontramos que A. durangensis es sensible al estrés por sequía similar a lo reportado anteriormente (Pompa-García et al., 2017), lo que pone en evidencia que será más vulnerable a las condiciones cada vez más cálidas y secas durante este siglo (Wang & Wang, 2024).
La sensibilidad de las especies forestales a las variaciones climáticas es contrastante, especialmente en términos de condiciones cálidas (Camarero et al., 2021). Por ejemplo, A. durangensis se correlacionó negativamente con las TMAX y TMIN en el mes de junio (Figura 4). En concordancia, las reducciones del crecimiento radial de TRW (Figura 2) decrecieron cuando comenzaron a incrementar las TMAX y TMIN a partir de 1970 (Figura 2 y 3) este mismo comportamiento también fue reportado por Pompa-García et al. (2017). Un coeficiente de correlación bajo no necesariamente indica que no haya una relación significativa; podría reflejar la complejidad de los factores ambientales y biológicos que interactúan en el proceso de crecimiento. A pesar de la existencia de una relación, otros factores influyentes en el crecimiento de los anillos no están siendo considerados en este análisis y, por lo tanto, podrían ser objeto de investigaciones adicionales para comprender mejor la variabilidad observada.
La falta de correlación entre la precipitación y el crecimiento radial sugiere que la respuesta del crecimiento a la humedad del suelo puede ser retardada y dependiente de otros factores como la temperatura y la luz solar. Además, el estrés hídrico acumulado puede limitar el crecimiento, resaltando la complejidad de la relación entre estas variables (Lloret et al., 2011). Además, se ha documentado que A. durangensisi responde significativamente al clima en su crecimiento radial, los anchos de los anillos son más sensibles a las condiciones de humedad, mientras que la densidad de la madera se ve más afectada por la temperatura. (González-Cásares, Pompa-García & Venegas-González, 2019).
Los meses de primavera y verano (marzo a agosto) muestran correlaciones altas. Estos meses coinciden con la temporada de crecimiento activo de los árboles. En otoño (septiembre a noviembre), las correlaciones altas sugieren que la disponibilidad de agua sigue siendo importante para el crecimiento radial, posiblemente debido a la acumulación de reservas y la preparación para el invierno. Incluso en diciembre, las correlaciones positivas indican que el balance hídrico puede estar reflejando los efectos acumulativos o residuales de las condiciones climáticas a lo largo del año. Las correlaciones mayores a 0.39 entre el SPEI y los meses del año de crecimiento destacan la importancia del balance hídrico en el crecimiento radial de los árboles. Estas relaciones subrayan la sensibilidad de A. durangensis a las variaciones climáticas, especialmente en términos de disponibilidad de agua.
Nuestros resultados sugieren que desde 1990 a 2022 han ocurrido 4 eventos de reducciones del crecimiento (1998, 1999, 2006 y 2011). Estos eventos han sido identificados en otras investigaciones como sequías en el norte del país (Pompa-García et al., 2017; Castruita-Esparza et al., 2019; Cabral-Alemán et al., 2022). Además, los años 1956, 1974 y 1982 también fueron registrados como reducciones de crecimiento por sequías en otras investigaciones del norte el país (Carlón Allende et al., 2021; Cabral-Alemán et al., 2022; Correa-Díaz et al., 2023). En los años 1878 y 1879 los estados del norte de México sufrieron una sequía que provocó gran escasez y carencia de alimentos (Cerano Paredes et al., 2011; Domínguez, 2016). Estos eventos también fueron registrados en esta investigación como eventos extremos de reducción del crecimiento radial.
En total fueron registradas 9 reducciones del crecimiento radial a lo largo de 1910-2020 años que corresponden a sequías históricas que azotaron el norte de México. Nuestros resultados sugieren que A. durangensis ha estado sometido a mayores presiones climáticas, lo que se refleja en una disminución de su resistencia y un aumento en la capacidad de recuperación. Esto ha llevado a una mayor resiliencia frente a la creciente recurrencia e intensidad de los periodos secos en México durante las últimas décadas (Stahle et al., 2016; Pompa-García et al., 2017; Correa-Díaz et al., 2023). Sin embargo, esta interpretación podría deberse a otros factores no climáticos, como la competencia intraespecífica, las condiciones edáficas o perturbaciones bióticas, que pudieron haber influido en el crecimiento de los anillos. Para mejorar la precisión, sería conveniente complementar el análisis con datos ambientales, como humedad del suelo, plagas y enfermedades que ayuden a determinar el origen multifactorial de las observaciones.
Los valores de resistencia, recuperación y resiliencia tienen concordancia con los registrados para otras especies mexicanas. Por ejemplo, Correa-Dìaz et al. (2023) encontraron que Abies concolor y Pinus jeffreyi presentaron una baja resistencia a la sequía, pero mostraron una alta capacidad de recuperación. En contraste con Pinus lumholtzi tuvo un valor de resistencia más alto, aunque su recuperación fue menor. Por otro lado, Pinus lambertana mostro una resistencia intermedia y una buena capacidad de recuperación. Además, también mostraron concordancia con especies como Pinus ponderosa Douglas en E.E.U.U y Araucaria araucana (Molina) en Argentina (Lloret et al., 2011; Piraino et al., 2022).
Una limitante de nuestra investigación fue la evaluación del comportamiento dendroclimático de un solo sitio para la especie, lo que no representa un gradiente geográfico amplio que podría revelar las limitaciones de la respuesta a los factores climáticos bajo condiciones diferentes. Además, otra limitante son los datos climáticos, pues al no tener datos climáticos detallados a nivel local podría subestimar la correlación precisa entre las variables climáticas con los anillos de crecimiento, lo que podría afectar en la exactitud de las conclusiones sobre la relación clima-crecimiento. Por tanto, se sugiere para futuros estudios complementar datos climáticos de mayor resolución u otras fuentes de datos climáticos.
Conclusiones
El crecimiento radial de A. durangensis durante el periodo 1910-2020 mostró una correlación positiva con las precipitaciones y negativas con la temperatura máxima y mínima. Este crecimiento fue especialmente sensible a los periodos de sequía acumulada, mientras que años con abundantes precipitaciones y temperaturas moderadas favorecieron el crecimiento radial.
A. durangensis muestra una notable capacidad para recuperarse de eventos climáticos extremos, aprovechando eficientemente los recursos hídricos y restaurando su tasa de crecimiento anterior. Aunque mantiene su resistencia durante períodos de sequía, la recuperación puede verse afectada. A pesar de un aumento en la resiliencia para recuperar tasas de crecimiento previas a estos eventos, la creciente frecuencia e intensidad de las sequías asociadas al cambio climático podría comprometer su capacidad de recuperación.
La protección de los BAVC es esencial para mantener servicios ecosistémicos críticos. Este estudio resalta la importancia de priorizar la conservación de estos bosques, enfocándose en estrategias prácticas que promuevan su resiliencia ante el cambio climático. Se recomienda adaptar las prácticas de manejo forestal para aumentar su resistencia a condiciones climáticas extremas, así como fomentar la reforestación garantizando la preservación de la biodiversidad y los servicios ecosistémicos.
Los estudios dendroecológicos no destructivos constituyen una evaluación del pasado que permite realizar una proyección del comportamiento futuro de nuestros bosques. Analizar los anillos de crecimiento mediante técnicas no destructivas ayuda comprender la vida y el comportamiento de las especies forestales, y permiten pronosticar como influyen los cambios ambientales en el crecimiento de especies forestales.

