












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
PermalinkIntroducción
El tomate (Solanum lycopersicum L.) es la hortaliza más cultivada bajo condiciones de invernadero en México (SIAP, 2023). la Universidad Autónoma Chapingo se ha desarrollado un sistema hidropónico de producción de tomate bajo invernadero (Sánchez-Del Castillo et al., 2021) en el cual se optimiza el uso del espacio y tiempo para incrementar la productividad anual. Se despuntan las plantas apenas por encima de la tercera inflorescencia para cosecharles solamente tres racimos a cada una y se manejan densidades de población de seis hasta 10 plantas m-². El ciclo de cultivo se acorta a menos de 120 d desde el trasplante hasta el fin de la cosecha, lo que hace posible obtener hasta tres ciclos anuales, cada uno con rendimientos de 160 t ha-¹, utilizando sistemas hidropónicos con sustratos de relativamente bajo costo como la arena de tezontle rojo, métodos de riego por goteo sencillos de implementar y manejar e invernaderos sólo con equipamiento básico de control de temperatura y humedad relativa (Mejía-Betancourt et al., 2023; Moreno-Pérez et al., 2021; Sánchez y Moreno, 2017; Sánchez-Del Castillo et al., 2012).
Con el sistema de producción arriba descrito, el rendimiento anual supera al sistema convencional de producción de jitomate en invernadero a niveles de mediana tecnología (Moreno-Pérez et al., 2021; Sánchez-Del Castillo et al., 2012); sin embargo, aún se puede incrementar si en cada racimo cuajaran más flores y frutos de los que produce cada racimo sin afectar el peso medio de fruto, ya que con sólo un fruto más que se lograra desarrollar en cada racimo, con una densidad de seis plantas m-2 y tres racimos por planta se podrían obtener 18 frutos más por m2 por cada ciclo de cultivo (54 frutos m-2 año-1); suponiendo un peso medio de 120 g por fruto, equivaldría a 64.8 t ha-1 año-1 adicionales.
La producción de una planta está influenciada por su genética (Heuvelink y Okello, 2018), pero factores ambientales como la radiación solar, la fertilización carbónica, la nutrición mineral y la regulación de las temperaturas diurnas y nocturnas tiene un impacto directo en el desarrollo de los primordios florales y en el porcentaje de los que llegan a flor (Heuvelink y Okello, 2018; Mejía-Betancourt et al., 2024; Yang et al., 2020). Estos factores afectan las relaciones fuente-demanda, la producción de fotoasimilados y el equilibrio entre crecimiento vegetativo y reproductivo (Lanoue et al., 2018; Mamatha et al., 2014; Rangaswamy et al., 2021). Por ello, autores como Heuvelink et al. (2020) y Wien (2020) sugieren que, para incrementar el número de flores por planta, es crucial estudiar los factores que aumentan la cantidad total de asimilados y aquellos que disminuyen la demanda diaria en otros órganos favoreciendo un mayor porcentaje de distribución hacia las inflorescencias en formación.
Por lo tanto, mejorar la velocidad y eficiencia de la fotosíntesis es una estrategia clave para aumentar la producción de biomasa y el rendimiento de los cultivos agrícolas (Ort et al., 2015), donde la luz es uno de los factores ambientales más influyentes en este proceso (Li et al., 2020). La extensión del periodo de iluminación con luz suplementaria en plántulas de tomate, a una densidad de flujo fotónico fotosintético de por lo menos 150 μmol de fotones m-2 s-1 durante la noche y por periodos cortos de tiempo (15-18 d) permitió un incremento de la integral diaria de radiación fotosintéticamente activa incidente (Moreno-Pérez et al., 2022). Esta práctica no solamente mejora la fotosíntesis, sino que también permite un mejor equilibrio entre el crecimiento vegetativo y el reproductivo, contribuyendo así a un aumento en el número de flores y frutos por planta (Lanoue et al., 2018; Wigge y Jaege, 2020).
La temperatura diurna y nocturna es otro factor ambiental clave en el crecimiento y floración de las plantas de tomate, pues influye en procesos fisiológicos esenciales como la fotosíntesis, la respiración y la translocación de fotoasimilados (Rodrigues et al., 2019; Sánchez y Moreno, 2017; Taiz et al., 2018; Yang et al., 2020). Se ha observado que temperaturas más bajas en las raíces de plántulas promueven un mayor número de flores en las primeras inflorescencias, lo que subraya la importancia de regular tanto la temperatura del aire como la del ambiente de la raíz (Heuvelink y Okello, 2018). El periodo en que la temperatura influye sobre la floración en tomate es durante la iniciación floral, que para la primera inflorescencia ocurre aproximadamente 8 d después de la expansión de los cotiledones (entre 15 y 20 d después de la siembra) y se extiende por una o dos semanas más para influir sobre la expresión de la segunda y tercera inflorescencia (Contreras-Magaña et al., 2013; Heuvelink y Okello, 2018).
También se ha reportado que la baja temperatura nocturna aumenta la acumulación de carbohidratos no estructurales que activan enzimas como la sacarosa sintasa, lo que favorece la formación de más flores por inflorescencia (Qi et al., 2011), así como una mayor ramificación floral (Heuvelink y Okello, 2018), posiblemente por una reducción en la tasa de respiración celular (Frantz et al., 2004), que ralentiza el crecimiento vegetativo y hace que más azúcares se destinen hacia los primordios florales en desarrollo, lo que reduce la tasa de aborto de éstos (He et al., 2019; Heuvelink et al., 2018; Yang et al., 2020).
La temperatura nocturna óptima para jitomate es entre 18 y 20 °C (Sánchez y Moreno, 2017); sin embargo, es posible bajarla aún más de manera temporal sin que se afecte el crecimiento y rendimiento, ya que esta especie puede tolerar desviaciones de 3 a 6 °C durante periodos cortos (Kubota et al., 2018; Papadopoulos y Hao, 2000), lo que permite manejar una temperatura media para el cultivo en lugar de mantener intervalos estrictos para los periodos diurnos y nocturnos, permitiendo un manejo flexible de la temperatura sin efectos adversos a la planta (Frantz et al., 2004; Kubota et al., 2018).
A pesar de su importancia, la investigación reciente sobre la aplicación de luz suplementaria para favorecer la tasa fotosintética o inducir efectos morfogénicos en plántula, la información sobre su efecto y sobre la acción conjunta con la temperatura nocturna incidente durante la diferenciación floral en las primeras inflorescencias es limitada; por ello, se llevó a cabo este estudio con el objetivo de determinar si la aplicación de luz suplementaria durante la noche y el manejo de las temperaturas nocturnas bajas durante la diferenciación floral de las primeras tres inflorescencias favorecen el proceso de fotosíntesis en las plántulas y modifican el número de flores y frutos de plantas de tomate despuntadas por encima del tercer racimo.
Materiales y métodos
Localización y condiciones del experimento
Durante la fase de semillero, que duró 34 días después de la siembra (dds), las plántulas crecieron en un invernadero de 300 m², ubicado en el campo experimental de la Universidad Autónoma Chapingo. El invernadero estuvo cubierto en la parte superior con polietileno térmico de alta dispersión lumínica, y en los frentes y costados por cortinas laterales móviles y mallas antiáfidos. Para controlar la temperatura y humedad relativa, se contaba con un sistema de enfriamiento evaporativo a base de muro húmedo de fibra de celulosa y extractores de aire, así como con un sistema de calefacción a base de gas LP. A los 35 dds se hizo el trasplante usando un segundo invernadero de 1000 m² con características similares al del semillero, donde las plantas permanecieron hasta finalizar su ciclo.
Material vegetal
Como material vegetal se utilizó tomate variedad Ramses, de la empresa Harris Moran, con frutos tipo saladette y hábito de crecimiento indeterminado.
Siembra y manejo de las plántulas
Para la siembra, se usaron charolas de poliestireno expandido con 128 cavidades, cada una con volumen de 46 cm3, previamente lavadas con agua y desinfectadas con una solución de sales cuaternarias de amonio (Anibac® Plus 45 %) a una concentración de 5 ml L-1 de agua, las cuales fueron rellenadas con una mezcla de peat moss y perlita en proporción 1:1 (v:v), colocando una semilla por cavidad.
A partir de la emergencia (4 dds) se inició la aplicación de riegos con solución nutritiva. En los primeros 15 dds la solución nutritiva contenía los siguientes elementos y concentraciónes (mg L-1): N = 100, P = 20, K = 125, Ca = 125, S = 75, Mg = 25, Fe = 1, Mn = 0.5, Cu = 0.05, Mn = 0.5, B = 0.25 y Zn = 0.05 (50 % de concentración), aplicando dos riegos por día y un gasto diario medio de 500 cm³ por charola. A partir de los 16 dds y hasta finalizar la cosecha, las concentraciones de elementos de la solución nutritiva se duplicaron.
Trasplante y manejo del cultivo
El trasplante se efectuó en camas de cultivo de 0.90 m de ancho por 20 m de largo, rellenas de tezontle rojo con partículas de 1 a 3 mm de diámetro. La densidad de población establecida fue 6 plantas m-² de invernadero. Cada planta fue tutorada con hilo de rafia y anillos plásticos, los brotes laterales fueron eliminados conforme aparecían para dejar un solo tallo por planta. A los 65 dds, con la tercera inflorescencia ya formada, se eliminó la yema terminal dejando dos hojas por encima del tercer racimo. Se implementó un sistema de riego por goteo, programado para proporcionar nueve riegos diarios entre las 9:00 y 17:00 h, con un gasto diario entre 0.9 y 1.0 L de solución nutritiva por planta.
Diseño experimental y tratamientos
Se utilizó un diseño experimental de bloques completos al azar con arreglo factorial, que incluyó dos condiciones de luz suplementaria y tres niveles de temperatura, dando lugar a un total de seis tratamientos con cuatro repeticiones; la unidad experimental constó de un grupo de 12 plantas.
os tratamientos fueron: Tratamiento 1) plántulas de tomate dentro del invernadero expuestas a temperatura nocturna de 18 ± 2 oC durante 18 d, sin luz suplementaria; Tratamiento 2) plántulas de tomate dentro del invernadero expuestas a temperatura nocturna de 18 ± 2 oC y con luz suplementaria nocturna durante 18 d; Tratamiento 3) plántulas de tomate expuestas a temperatura nocturna de 11 ± 1 oC durante 18 d, colocadas dentro de una cámara frigorífica en un horario de 18:00 a 6:00 h del siguiente día y sin luz suplementaria, durante el periodo diurno las plántulas eran colocadas dentro del invernadero usado como semillero; Tratamiento 4) plántulas de tomate expuestas a temperatura nocturna de 11 ± 1 oC y con luz suplementaria nocturna durante 18 d colocadas dentro de una cámara frigorífica en un horario de 18:00 a 6:00 h del siguiente día, durante el periodo diurno las plántulas eran colocadas dentro del invernadero usado como semillero; Tratamiento 5) plántulas de tomate expuestas a temperatura nocturna de 15 ± 1 oC durante 18 d colocadas dentro de una cámara frigorífica en un horario de 18:00 a 6:00 h del siguiente día y sin luz suplementaria, durante el periodo diurno las plántulas eran colocadas dentro del invernadero usado como semillero; Tratamiento 6) plántulas de tomate expuestas a temperatura nocturna de 15 ± 1 oC y luz suplementaria nocturna durante 18 d, colocadas dentro de una cámara frigorífica, en un horario de 18:00 a 6:00 h del siguiente día, con luz suplementaria nocturna. Todos los tratamientos fueron aplicados del día 17 al 34 después de la siembra.
La luz suplementaria fue proporcionada con lámparas led (IGOTO Swiss, modelo LS01, Yueqing, China) de 120 cm de largo por 7 cm de ancho, que emitían luz blanca a una densidad de flujo fotónico fotosintético de 175 μmol de fotones m-2 s-1, por 8 h colocadas 15 cm por arriba del ápice de las plántulas.
Para separar las plántulas bajo tratamiento con luz suplementaria de los tratamientos sin luz, tanto dentro de las cámaras como en el invernadero, se colocó entre ambos tratamientos de temperatura, una lámina de cartón prensado, revestida con papel vinil adhesivo mate de color negro.
Variables evaluadas
El día del trasplante se hicieron mediciones de altura de plántula (cm) con un flexómetro; diámetro del tallo (mm), entre la segunda y tercera hoja utilizando un vernier electrónico (Keatronic, modelo A150, County Mayo, Irlanda); área foliar (cm2), determinada con un integrador de área foliar (LI-COR® LI-3000A, Lincoln, Nebraska, EUA); volumen de raíz (cm3) utilizando el método de desplazamiento de agua que consistió en sumergir el total de raíces previamente lavadas en un volumen conocido de agua contenida en una probeta; peso seco total por plántula (g), para ello se secaron las plantas en estufa a 70 ºC por 72 h y después se pesaron utilizando una balanza analítica (Radwag® AS 220.R2 Plus, Radom, Polonia); largo y ancho de las hojas en cm, utilizando un flexómetro; grosor de la lámina foliar en mm utilizando un vernier electrónico (Keatronic, modelo A150), y el contenido de clorofila total, expresado en µg g-1, siguiendo la técnica descrita por Lichtenthaler (1987) en una muestra de 10 g de hoja por unidad experimental. Posteriormente, se cuantificó el número de flores por racimo y por planta, el número de frutos cosechados por planta, el peso promedio de los frutos (g) y el rendimiento (kg/planta).
Análisis estadístico
Los datos fueron sometidos a análisis de varianza y comparaciones de medias (Tukey, P≤ 0.05) utilizando el programa estadístico Statistical Analysis System 9.1 (SAS Institute, 2002).
Resultados y discusión
Los análisis de varianza en plántulas (datos no mostrados), mostraron diferencias significativas entre las condiciones de luz suplementaria para área foliar y peso seco, y entre las diferentes temperaturas nocturnas para todas las variables evaluadas.
Las comparaciones de medias entre tratamientos de luz suplementaria muestran que la luz adicional no modificó la altura de planta, diámetro de tallo, ni volumen de raíz, pero incrementó en 59.5 cm-2 el área foliar (plántulas con área foliar de 212.8 cm²), lo que condujo a plántulas con mayor peso seco total (1.11 g) (Cuadro 1). Según Taiz et al. (2018), la luz adicional estimula la fotosíntesis, lo que favorece el crecimiento, reflejado en un aumento en el peso seco, como lo observado en el presente estudio. Aunque autores como Moreno et al. (2022) han reportado de manera consistente disminuciones en la altura de plántula por efecto de luz led suplementaria, en este estudio no se observó tal modificación.
Cuadro 1 Comparación de medias de variables de crecimiento en plántulas de tomate de 35 días de edad manejadas bajo dos condiciones de luz suplementaria y tres temperaturas nocturnas.
| Tratamiento | Altura (cm) | Diámetro de tallo (mm) | Área foliar (cm2) | Volumen de raíz (cm3) | Peso seco total (g) |
| Condición de la luz suplementaria | |||||
| Con luz | 19.2 a | 3.80 a | 212.8 a | 1.28 a | 1.11 a |
| Sin luz | 19.5 a | 3.74 a | 153.3 b | 1.13 a | 0.78 b |
| DSH (0.05) | 1.47 | 0.19 | 19.51 | 0.21 | 0.14 |
| Temperatura nocturna | |||||
| 11 ± 1 °C | 15.5 c | 3.84 a | 158.9 b | 1.35 a | 0.84 b |
| 15 ± 1 °C | 18.9 b | 3.94 a | 186.5 ab | 0.97 b | 0.92 ab |
| 18 ± 2 °C | 23.6 a | 3.54 b | 203.8 a | 1.30 a | 1.07 a |
| DSH (0.05) | 2.18 | 0.28 | 29.03 | 0.31 | 0.20 |
Valores con la misma letra dentro de cada columna y factor de variación son estadísticamente iguales (Tukey, P ≤ 0.05). DSH: diferencia significativa honesta.
En cuanto al efecto de la temperatura nocturna (Cuadro 1), se encontró que las plántulas expuestas a temperaturas más altas (18 °C) mostraron el mayor desarrollo en términos de altura, área foliar y peso seco, con diferencias respecto al tratamiento de menor temperatura (11 °C), el cual afectó negativamente el crecimiento. Al respecto, Van-der-Ploeg et al. (2007) y Pereira y Sampaio (2023) indicaron que la exposición de las plántulas a bajas temperaturas provoca una inhibición del crecimiento debido a una reducción en la actividad enzimática y, en consecuencia, en la actividad respiratoria.
Frantz et al. (2004) señalaron que la respiración de las plantas de tomate aumenta de 2 a 7 % por cada grado centígrado de incremento en la temperatura nocturna, o disminuye en la misma proporción cuando la temperatura baja, y que las plantas responden a cambios de temperatura en periodos de menos de 20 d, ya que después se aclimatan, por lo que el impacto en la actividad respiratoria y en el crecimiento, deja de ser significativo debido a pérdidas de sensibilidad a esos cambios, fenómeno que quizá ocurrió en la respuesta diferenciada de las plántulas al ser expuestas a temperaturas bajas durante 17 d.
Aunque con temperatura de 18 °C las plántulas presentaron mayor altura, éstas tuvieron menor diámetro de tallo, lo que reduce calidad al trasplante. En este sentido, las plántulas que crecieron a 15 °C presentaron mejores características para el trasplante, ya que fueron más compactas y con mayor diámetro de tallo. Estos resultados son consistentes con Wien (2020), quien mencionó que la tasa de elongación del tallo generalmente aumenta con la temperatura, pues hay mayor síntesis de giberelinas que estimulan el alargamiento celular; además, cuando hay una diferencia importante entre la temperatura diurna y nocturna ocurre una reducción en el crecimiento en longitud del tallo (Heuvelink et al., 2020; Taiz et al., 2018), como ocurrió con el tratamiento de menor temperatura.
En volumen de raíz, Pereira et al. (2021) reportaron que las altas temperaturas promueven una mayor ramificación, lo que incrementa su volumen; en contraste, a temperaturas bajas como 14 °C, las raíces tienden a ser más gruesas y menos ramificadas, aunque hay otros factores como la nutrición mineral y el tipo de sustrato que también influyen en el crecimiento radical.
Las comparaciones de medias de variables medidas en la hoja (Cuadro 2), mostraron que con luz suplementaria las plántulas tuvieron mayor largo y ancho de hoja, y mayor grosor de la lámina foliar, comparadas con las plántulas sin luz adicional, lo que indica que la luz led adicional, además de promover crecimiento foliar, indujo efectos morfogenéticos para formar hojas con mayor densidad, como también lo señalaron Taiz et al. (2018). El mayor largo y ancho de la hoja con luz suplementaria contribuyó a formar plántulas de mayor área foliar (Cuadro 1). También se encontró que con temperaturas de 11 ± 1°C las plántulas redujeron significativamente el largo y ancho de hoja e incrementaron su grosor respecto al tratamiento de 18 ± 2°C, mientras que con 15 y 18 °C las dimensiones en hoja fueron similares (Cuadro 2).
Cuadro 2 Comparación de medias de variables en hojas de plántulas de tomate de 35 días de edad manejadas bajo dos condiciones de luz suplementaria y tres temperaturas nocturnas.
| Tratamiento | Largo de hoja (cm) | Ancho de hoja (cm) | Clorofila total (µg g-1) | Grosor de lámina foliar (mm) |
| Condición de la luz suplementaria | ||||
| Con luz | 9.71 a | 10.67 a | 17.17 a | 0.27 a |
| Sin luz | 9.03 b | 9.77 b | 16.02 a | 0.22 b |
| DSH (0.05) | 0.46 | 0.54 | 1.17 | 0.04 |
| Temperatura nocturna | ||||
| 11 ± 1 °C | 8.61 b | 9.39 b | 16.10 a | 0.27 a |
| 15 ± 1 °C | 9.4 a | 10.31 a | 17.27 a | 0.24 ab |
| 18 ± 2 °C | 10.08 a | 10.95 a | 16.42 a | 0.22 b |
| DSH (0.05) | 0.69 | 0.8 | 1.76 | 0.05 |
Valores con la misma letra dentro de cada columna y factor de variación son estadísticamente iguales (Tukey, P ≤ 0.05). DSH: diferencia significativa honesta.
El mayor grosor de la lámina foliar con 11 ± 1 °C (0.27 mm) puede ser atribuido a un estrés térmico por baja temperatura, pues bajo estas condiciones se reducen las actividades enzimáticas relacionadas con la síntesis de hormonas de crecimiento como giberelinas y auxinas (Wien, 2020), que compactan el tejido foliar (células más pequeñas), dando lugar a la formación de hojas más pequeñas, pero más gruesas, ésto como un mecanismo para conservar calor y resistir el estrés ocasionado por bajas temperaturas (Taiz et al., 2018). Heuvelink et al. (2018) también mencionaron que la tasa de crecimiento en plántulas de tomate se reduce con temperaturas nocturnas bajas, disminuye el área foliar, y las temperaturas subóptimas producen hojas más gruesas debido a células más grandes que acumulan almidón.
En síntesis, la presencia de luz suplementaria nocturna y de temperaturas nocturnas de 15 a 18 ºC en el periodo de iniciación floral de las tres primeras inflorescencias (17 a 34 dds), influyeron en la producción temporal de más área foliar y más peso seco por planta, indicando la producción de más fotoasimilados por plántula por día en ese periodo. Estas condiciones pueden hacer que más primordios de flor en cada inflorescencia dispongan de suficientes fotoasimilados para no abortar y desarrollarse hasta flores en antesis (Heuvelink et al., 2018). El menor grosor de tallo, volumen de raíz y área foliar en las plántulas con el tratamiento de 15 ºC, respecto a las plantas a 18 ºC, sugiere que pudo haber menos gasto de fotoasimilados en crecimiento vegetativo.
En cuanto al contenido de clorofila total, no hubo cambio alguno por efecto de la luz suplementaria y tampoco fue modificado por la temperatura nocturna aplicada en las plántulas (Cuadro 2). El análisis de varianza indicó efectos significativos para la interacción luz × temperatura nocturna respecto a altura de plántula y clorofila total (Figura 1). La interacción para altura de plántula se explica porque con luz suplementaria y mayor temperatura, las plántulas incrementan su altura en mayor proporción que las plántulas sin luz a la misma temperatura, probablemente por síntesis de más auxinas o giberelinas que intervienen en la elongación celular (Wien, 2020), o porque la combinación de luz suplementaria y temperatura nocturna más alta provocaron un metabolismo más acelerado reflejado en una tasa de producción y uso de asimilados más rápida que favoreció un mayor crecimiento (Taiz et al., 2018). La interacción significativa para la concentración de clorofila total se debe a que cuando las plántulas fueron manejadas a 15 °C durante la noche, con luz suplementaria se incrementó la concentración de clorofila total respecto al tratamiento con luz a 11 ºC; sin embargo, al aumentar la temperatura a 18 °C con luz suplementaria, se redujo la concentración de clorofila, mientras que sin luz led suplementaria la concentración fue casi constante bajo cualquier nivel de temperatura evaluado.
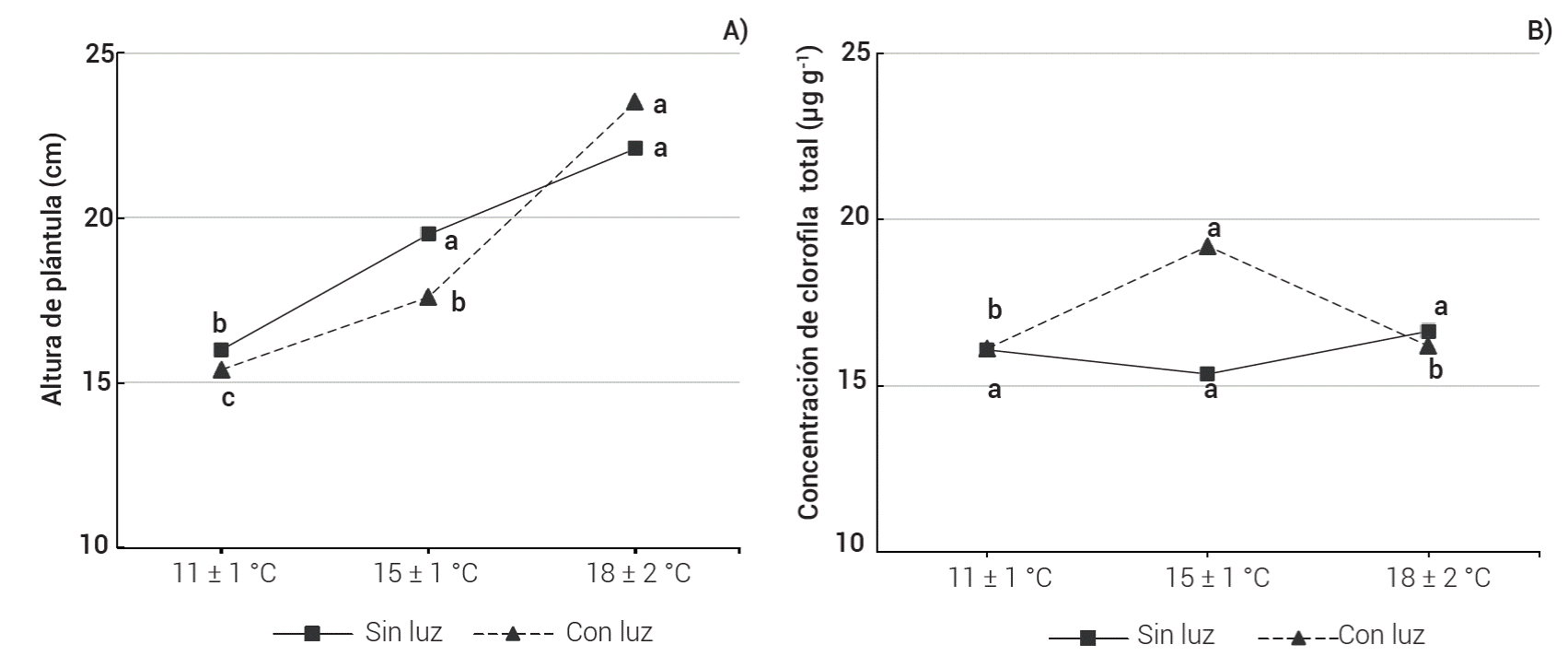
Figura 1 Interacción entre condición de luz suplementaria y temperatura nocturna para altura de plántula (A) y concentración de clorofila total (B) en plántulas de tomate, 35 días después de la siembra.
Cuando se elevó la temperatura de 11 a 15 ºC y se aportó luz suplementaria, la concentración de clorofila aumentó (Figura 1B), probablemente debido a una mayor síntesis de clorofila provocada por un incremento en la integral diaria de luz respecto al tratamiento sin luz suplementaria (Taiz et al., 2018) combinado con el aporte de 4 ºC más de temperatura nocturna promedio en relación con el tratamiento de 11 ºC, lo que seguramente aceleró la actividad enzimática y el proceso de biosíntesis en general. Al elevar la temperatura de 18 a 20 ºC se formó numéricamente más área foliar que a 15 ºC (Cuadro 1) ocasionando un efecto de dilución que redujo la concentración de clorofila otra vez a niveles similares a los del tratamiento de 11 ºC.
Con respecto a las variables de rendimiento y sus componentes, el análisis de varianza (datos no mostrados) y comparaciones de medias indicaron que la luz nocturna suplementaria no afectó ninguna de las variables evaluadas, pero sí se tuvieron efectos de temperatura nocturna, tampoco hubo efectos significativos de interacción entre condiciones de luz suplementaria y temperatura nocturna aplicadas de los 17 a los 34 dds.
Se encontró que las plántulas expuestas a 15 ± 1 °C dieron lugar a plantas con mayor número de flores en el primer y tercer racimo (Cuadro 3); en consecuencia, más flores por planta (tres flores más por planta). En número de frutos por planta, también hubo un incremento estadísticamente significativo a favor del tratamiento de 15 ± 1 °C, respecto al tratamiento de 11 °C. Aunque el peso de fruto fue igual en todos los tratamientos, la diferencia en el número de frutos formados por planta no fue suficiente para que se tuvieran cambios significativos en el rendimiento. Estos resultados permiten deducir que el régimen de temperatura nocturna durante el periodo de iniciación de las inflorescencias afecta el porcentaje de aborto de los primordios florales, lo cual concuerda con Heuvelink y Okello (2018), quienes mencionaron que el número de flores no solamente está determinado por el cultivar, sino también por las condiciones ambientales en que las plantas crecen. Por su parte, Qi et al. (2011) reportaron que temperaturas nocturnas relativamente bajas favorecen la acumulación de carbohidratos no estructurales que ayudan a prevenir la aborción de primordios florales, aumentando así el número de flores y frutos en plantas de tomate, como ocurrió en este estudio.
Cuadro 3 Comparación de medias entre tratamientos de variables del rendimiento y sus componentes evaluadas en planta de tomate variedad Ramses.
| Tratamiento | NFLR1 | NFLR2 | NFL3 | NFLPL | NFRPP | PMFR (g) | RENPP (kg/planta) |
| Condición de la luz suplementaria | |||||||
| Con luz | 9.3 a | 10.0 a | 10.8 a | 30.1 a | 25.3 a | 111 a | 2.81 a |
| Sin luz | 9.1 a | 9.5 a | 10.4 a | 28.9 a | 24.9 a | 119 a | 2.95 a |
| DSH (0.05) | 0.45 | 0.70 | 0.73 | 1.14 | 1.79 | 11.20 | 0.27 |
| Temperatura nocturna | |||||||
| 11 ± 1 °C | 8.7 b | 9.4 a | 10.2 b | 28.2 b | 23.9 b | 118 a | 2.83 a |
| 15 ± 1 °C | 9.7 a | 10.2 a | 11.3 a | 31.3 a | 26.6 a | 110 a | 2.94 a |
| 18 ± 2 °C | 9.1 ab | 9.7 a | 10.2 b | 29.0 b | 24.6 ab | 117 a | 2.87 a |
| DSH (0.05) | 0.68 | 1.04 | 1.08 | 1.69 | 2.66 | 16.67 | 0.40 |
Valores con la misma letra dentro de cada columna y factor de variación son estadísticamente iguales (Tukey, P ≤ 0.05). DSH: diferencia significativa honesta, NFLR1: número de flores en el racimo uno, NFLR2: número de flores en el racimo dos, NFLR3: número de flores en el racimo tres, NFRPP: número de frutos por planta, PMFR: peso medio de fruto, RENPP: rendimiento por planta.
Finalmente, el rendimiento de 2.94 kg por planta alcanzado a una temperatura de 15 °C en etapa de plántula y con la densidad de 6 plantas m-2 de invernadero establecida (17.64 kg m-²) es alto para un ciclo de cultivo de trasplante a cosecha de solamente 110 d.
Conclusiones
Los resultados obtenidos permiten concluir que la luz led suplementaria nocturna a una densidad de flujo fotónico fotosintético de 175 μmol de fotones m-2 s-1 aplicada en etapa de plántula en tomate, incrementa el área foliar y el peso seco de las plántulas al momento del trasplante, pero no tiene ningún efecto sobre el número de flores y frutos formados en las tres primeras inflorescencias de las plantas. Cuando las plántulas de tomate fueron expuestas a temperatura nocturnas de 15 ± 1 °C durante el periodo sensitivo de la diferenciación floral en las tres primeras inflorescencias, además de obtenerse plántulas con calidad al trasplante, se promovió un mayor número de flores y frutos por planta respecto a cuando estuvieron expuestas a 18 ± 2 ºC. En ese mismo periodo, con temperatura nocturna de 11 ± 1 ºC, el crecimiento de las plántulas se redujo y posteriormente expresaron menor número de flores y frutos por planta.

