












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
PermalinkIntroducción
Taxus globosa es una especie que pertenece a la familia Taxaceae, se caracteriza por ser un árbol perennifolio, dioico, de 6 a 18 m de altura y que forma un tronco erecto de 30 a 50 cm de diámetro (Shemluck et al., 2003). Esta especie se encuentra restringida en el mundo, su distribución se limita a México, Honduras, Guatemala y El Salvador. El área de distribución potencial de esta especie en México representa el 0.5 % de su territorio (García-Aranda et al. 2010); esta área tiende a reducirse debido a la deforestación, cambio de uso del suelo y fragmentación del hábitat (Contreras-Medina et al., 2010). Asimismo, las plantas de T. globosa muestran crecimiento lento, capacidad reproductiva escasa y distribución dispersa. En México T. globosa se encuentra dentro de la Norma
Oficial Mexicana NOM-059-SEMARNAT (SEMARNAT, 2010) en la categoría de ‘protección especial’ (Antúnez, 2021), situación que conlleva a la necesidad de promover su conservación y recuperación. La regeneración y recuperación de las poblaciones de T. globosa se basa principalmente en la producción de semillas, puesto que es una fuente importante de variabilidad genética; no obstante, la producción de semillas es escasa (6 a 15 semillas maduras por árbol) y su germinación presenta algunos inconvenientes (Ramírez-Sánchez et al., 2011).
Asimismo, las especies del género Taxus producen un metabolito secundario conocido como taxol, el cual es utilizado con fines farmacéuticos para el tratamiento de algunos tipos de cáncer, como el de mama y de ovario (Wei et al., 2023) . La extracción de su corteza y hojas para la obtención del taxol ha promovido la explotación ilegal de esta especie (Jia et al., 2022). Se sabe que un árbol maduro produce 3 kg de corteza, a partir de los cuales solo es posible obtener 300 mg de taxol (Appendino et al., 1994).
Ramírez-Sánchez et al. (2018) mencionaron que las semillas de Taxus globosa presentan latencia morfofisiológica, por lo cual la germinación de estas semillas ocurre hasta después de uno o dos años de haber sido dispersadas. La latencia es un proceso mediante el cual las semillas no germinan aun cuando se les provee de las condiciones óptimas para que ésta ocurra (Pausas y Lamont, 2022). La latencia es una estrategia de las semillas que impide que germinen cuando se exponen a condiciones adversas, las cuales podrían poner en riesgo su supervivencia (Lennon et al., 2021). Entre los factores que pueden causar latencia destacan los anatómicos (estructura de la testa), fisiológicos (presencia de inhibidores de la germinación) y morfológicos (madurez del embrión) (Miransari y Smith, 2014).
En algunas semillas, la latencia puede ser causada por más de un factor (Pausas y Lamont, 2022). En las semillas con latencia morfofisiológica, el embrión no ha terminado de desarrollarse al momento que es dispersada (Zhang et al., 2023); además, las semillas pueden presentar inhibidores de la germinación en la testa o en el megagametofito (Chenyin et al., 2023). Al respecto, Liu et al. (2011) encontraron que compuestos fenólicos como los flavonoides, presentes tanto en la testa como en el embrión de las semillas de T. chinensis var. Mairei, tienen la capacidad de inhibir su germinación.
Por otro lado, las auxinas, citocininas, giberelinas y ácido abscísico, son hormonas con actividad importante en el desarrollo y germinación de las semillas, así como en la señalización para la comunicación entre sus tres componentes: embrión, endospermo y testa (Miransari y Smith, 2014). Uno de los tratamientos más utilizados para estimular la germinación de semillas latentes de especies forestales de climas templados es la estratificación (Song et al., 2023). Esta práctica consiste en exponer las semillas a temperaturas frías (4 a 10 °C) o cálidas (15 a 25 °C), en condiciones de humedad. Otras formas de liberar de la latencia a las semillas son la escarificación (física o química), la aplicación de ácido giberélico (AG3) o la combinación de estos métodos (Nawrot-Chorabik et al., 2021; Stejskalová et al., 2015).
Los trabajos relacionados con el rompimiento de la latencia de las semillas de T. globosa son escasos, por lo que es necesario llevar a cabo investigaciones enfocadas a conocer las causas de dicha latencia y las estrategias que permitan romperla. Por lo anterior, el objetivo de la presente investigación fue analizar los cambios morfológicos y fisiológicos en las semillas de T. globosa sometidas a distintos tratamientos para romper la latencia.
Materiales y métodos
Material biológico
Las semillas de Taxus globosa se recolectaron en la localidad El Chico, Hidalgo, México, de septiembre a diciembre del año 2021. Las semillas se colocaron en bolsas de papel y se mantuvieron en refrigeración a 4 °C, durante 90 días (hasta su uso). Antes de aplicar los tratamientos se formaron cuatro grupos de 15 semillas elegidas al azar, las cuales fueron seccionadas longitudinalmente a través del eje embrionario e inmediatamente sumergidas en una solución de tetrazolio 1 %, a temperatura ambiente, durante 24 h; posteriormente, se evaluó el grado de tinción en el embrión, considerando como viables aquellas semillas cuya área se tiñera en un 70 % y las que no se tiñeran como inviables (ISTA, 2016).
Tratamientos para romper la latencia
Cuatro repeticiones de 20 semillas cada una se sometieron a tres tratamientos; además, se consideró un testigo que consistió en semillas secas al tiempo de inicio (Cuadro 1). Previamente, las semillas se desinfectaron con dióxido de cloro y agua destilada en proporción de 40:60 v/v. Para la escarificación, las semillas fueron embebidas en agua durante 12 h y luego se sumergieron en nitrógeno líquido (N2) durante 10 s. Para la estratificación, las semillas previamente embebidas durante 12 h y escarificadas con N2, se expusieron a 4 oC (estratificación fría) o 25 oC (estratificación cálida) durante 30 días. Después de someter las semillas a los tratamientos, se colocaron sobre papel absorbente humedecido dentro de contenedores de plástico con tapa. Los recipientes que contenían las semillas se incubaron a 25/13 oC y 12 h luz/12 h oscuridad. Las semillas de los Tratamientos 2 y 3 se expusieron de manera consecutiva a las distintas condiciones señaladas en el Cuadro 1. Cada 30 días, durante 120 días, se evaluó el porcentaje de germinación y el porcentaje de semillas con testa abierta en los distintos tratamientos.
Cuadro 1 Tratamientos aplicados a las semillas de Taxus globosa para romper la latencia.
| Testigo | Tratamientos | ||
| 1 | 2 | 3 | |
| Semillas secas sin escarificar con N2 líquido, en el tiempo cero | Escarificación con N2 Sumergir las semillas por 10 s en N2 | Escarificación con N2 (10 s) Ácido
giberélico4/7: 300 mg L-1 (48
h) Estratificación fría: humedad + 4 °C, 30 d Estratificación cálida: humedad + 25 °C, 30 d. Estratificación fría: humedad + 4 oC, 30 d |
Escarificación con N2 (10 s) Estratificación
cálida: humedad + 25 °C, 30 d. Escarificación con N2 (10 s) Ácido giberélico 4/7: 300 mg L-1 (48 h) |
Análisis morfométrico y viabilidad
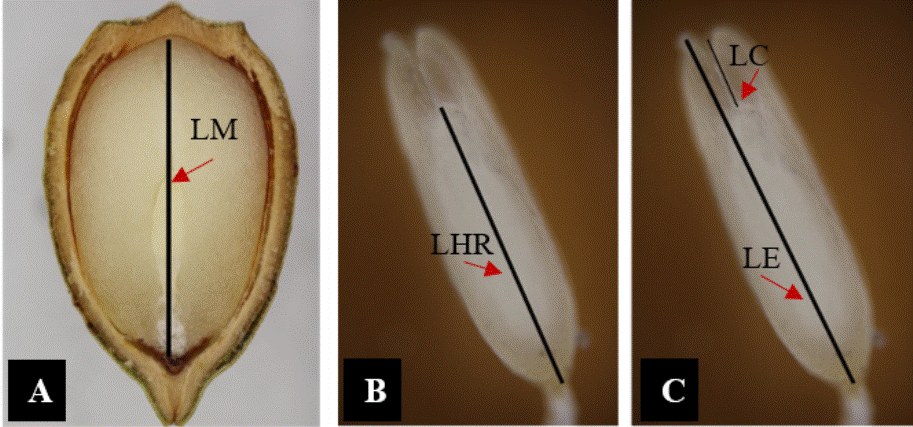
Se analizaron 10 semillas testigo al tiempo cero (semillas secas) y 10 semillas de cada tratamiento después de 120 días de su aplicación. Las semillas fueron seccionadas longitudinalmente y observadas con un microscopio estereoscópico (AmScope Discovery V20 Carl Zeiss®, Oberkochen, Alemania) y las imágenes se capturaron con una cámara (Cannon, EOS Rebel T7®, Tokio, Japón). Las variables evaluadas fueron longitud del megagametofito, longitud del embrión, la longitud de los cotiledones y del eje hipocótilo-radicular, todas ellas en mm (Figura 1). Las mediciones se hicieron con el software ImageJ 1.53k.
Cuantificación de hormonas y fenoles
La determinación de hormonas y fenoles se llevó a cabo en 800 mg de semillas expuestas a los distintos tratamientos después de 120 días, así como en el testigo al tiempo cero. Se analizaron semillas sin testa, a lo que se le denominó megagametofito, y la testa de éstas, mediante el protocolo desarrollado por Pan et al. (2010). El protocolo consistió en congelar el material vegetal con N2 líquido y molerlo; luego, se agregó una mezcla de propanol:agua:HCl concentrado (2:1:0.002 v/v). La mezcla con el material vegetal se agitó a 100 rpm por 30 min y se le añadió 1 mL de diclorometano; se agitó nuevamente y se centrifugó por 5 min. Después, se obtuvieron 900 µL de la parte baja del solvente, la cual se concentró con la ayuda de una corriente de nitrógeno y se añadieron 0.5 mL de metanol; al finalizar, se inyectaron 50 µL de la solución en el cromatógrafo líquido de alta resolución (HPLC) (Serie 200, Perkin Elmer®, Waltham, Massachusetts, EUA) y se utilizó una columna de la marca Rx/SB-C8 Rapid Res, HPLC Col. 4.6 × 75 (Perkin Elmer®, Waltham, Massachusetts, EUA).
Diseño experimental y análisis estadístico
Para analizar la viabilidad se utilizaron cuatro repeticiones de 15 semillas cada una, a partir de las cuales se obtuvo el valor promedio expresado en porcentaje y su error estándar. El experimento para probar el efecto de los tratamientos para romper la latencia de las semillas tuvo un diseño completamente al azar con tres tratamientos y un testigo (semillas secas al tiempo cero), con cuatro repeticiones de 20 semillas cada una. El análisis morfométrico también tuvo un diseño completamente al azar con tres tratamientos y un testigo; cada tratamiento constó de 10 semillas, considerando una semilla como una repetición. Para el análisis de hormonas y fenoles, el diseño experimental fue completamente al azar con dos factores: tratamientos para romper la latencia (tres niveles más un testigo al tiempo cero) y la estructura de la semilla (dos niveles); cada tratamiento constó de tres repeticiones. Los datos obtenidos se analizaron mediante análisis de varianza con el paquete estadístico R-Studio (R Core Team, 2021) y la comparación de medias se llevó a cabo con la prueba de Tukey (P ≤ 0.05). Para la variable ácido rosmarínico, previo al análisis, se llevó a cabo una transformación logarítmica de los datos.
Resultados y discusión
La viabilidad de las semillas al tiempo cero fue de 87.0 + 2.76 %, esto indica que la mayoría de las semillas que se utilizaron para probar los tratamientos estaban vivas y tenían la capacidad para germinar. Esta prueba permitió conocer de manera rápida y confiable si la falta de germinación de las semillas de T. globosa podría deberse a la baja viabilidad del lote o a que éstas presentaban latencia.
Efecto de los tratamientos en la germinación de las semillas
No fue posible observar semillas germinadas en ninguno de los tratamientos probados, sólo en el Tratamiento 1 (escarificadas con N2) 3.8 % de las semillas mostraron la testa abierta después de 90 días. Liu et al. (2011) reportaron que usar la estratificación fría (4 °C), seguida de la estratificación cálida (25 °C) y nuevamente estratificación fría, durante tres meses (en cada una), promovió la germinación de 23.7 % de las semillas de T. chinensis var. Mairei después de los 280 días. Por otra parte, Ramírez-Sánchez et al. (2018) encontraron que combinar la estratificación cálida (25 °C) con la estratificación fría (3 °C) y la aplicación de AG4/7 (500 ppm), seguida de escarificación con N2, promovió la germinación de 12 a 14 % de las semillas de T. globosa, 266 días después de aplicar los tratamientos. Lo anterior indica que la germinación de las semillas del género Taxus, aun con la aplicación de tratamientos para romper la latencia, podría requerir de al menos 266 días.
Análisis morfométrico
No se encontraron diferencias significativas en el tamaño de las semillas de T. globosa sometidas a los distintos tratamientos probados. Las semillas tuvieron una longitud promedio entre 6.0 y 6.5 mm, valores superiores a los observados por Ghimire et al. (2019) en semillas de T. cuspidata y T. caespitosa (5.42 y 5.25 mm, respectivamente). Por otro lado, la longitud del embrión en las semillas del Tratamiento 3 solo fue significativamente menor (0.94 mm) que el de las semillas del Tratamiento 1 (1.85 mm) (Cuadro 2). En cuanto a la longitud del megagametofito, no se observaron diferencias significativas entre las semillas de los distintos tratamientos y el testigo (Cuadro 2). Al respecto, el tamaño del megagametofito y del embrión de T. globosa al día cero (testigo) coincide con lo reportado por Ramírez-Sánchez et al. (2011), quienes encontraron que las semillas de T. globosa de la región centro de México mostraron una longitud promedio de 4.8 mm y 1.7 mm, respectivamente; asimismo, el tamaño de los cotiledones no fue significativamente distinto entre los tratamientos (1.65 mm, promedio), en tanto que la longitud del eje hipocótilo-radicular fue menor en las semillas del Tratamiento 3 (0.6 mm) (Cuadro 2). La aplicación del Tratamiento 3 (escarificación con N2 + estratificación cálida + escarificación con N2 + AG 4/7) pudo haber causado daño en las semillas, lo cual se manifestó por cambios en el color (oscurecimiento) de la testa, megagametofito y embrión, así como por la reducción en la longitud del embrión y del eje hipocótilo-radicular (Cuadro 2).
Cuadro 2 Análisis morfométrico de las semillas de Taxus globosa después de 120 días de exponerse a distintos tratamientos para romper la latencia.
| Tratamiento | Longitud del embrión | Longitud del megagametofito | Longitud de los cotiledones | Longitud del EHR |
| Testigo | 1.77 ab | 4.8 a | 1.9 a | 1.3 ab |
| 1 | 1.85 a | 4.8 a | 1.2 a | 1.4 a |
| 2 | 1.64 ab | 4.8 a | 1.8 a | 1.2 ab |
| 3 | 0.94 b | 4.7 a | 1.7 a | 0.6 b |
Los valores se expresan en mm. EHR: eje hipocótilo radicular. Medias con letras iguales dentro de una columna no son estadísticamente diferentes (Tukey, P ≤ 0.05).
Bian et al. (2018) evaluaron la longitud de la semilla y del embrión de T. yunnanensis, de los 30 a los 360 días de ser sembradas en arena húmeda a 16.5 °C. Observaron que la longitud del megagametofito fue de 6.89 y 6.94 mm a los 30 y 180 días, respectivamente, en tanto que la longitud del embrión fue de 2.58 y 2.69 mm a los 90 y 180 días.
Hormonas
Al día cero (testigo) fue posible detectar en el megagametofito la presencia de trans-zeatina, cis-zeatina, cinetina, ácido indolacético (AIA) y ácido abscísico (ABA), mientras que en la testa se encontró cis-zeatina, cinetina, ácido indolacético y ABA, en tanto que en los tratamientos se encontraron diferencias significativas en el contenido de trans-zeatina en el megagametofito, siendo las semillas del Tratamiento 1 (escarificadas con N2 líquido) las que produjeron la mayor cantidad (0.0116 µg mg-1) (Cuadro 3). Por otro lado, la cis-zeatina y el ABA no estuvieron presentes en el megagametofito de las semillas de ninguno de los tratamientos probados.
Cuadro 3 Fitohormonas presentes en el megagametofito y testa de semillas de Taxus globosa sometidas a distintos tratamientos para romper la latencia, después de 120 días.
| Trat | Estructura | TZ | CZ | CIN | AIA | ABA |
| (µg mg-1) | ||||||
| Testigo | Megagametofito | 0.00052 c | 0.02128 a | 0.01909 a | 0.01773 a | 0.00103 a |
| 1 | 0.01169 a | NP | 0.01109 a | NP | NP | |
| 2 | 0.01010 b | NP | NP | NP | NP | |
| 3 | 0.00857 c | NP | NP | 0.00915 a | NP | |
| Testigo | Testa | NP | 0.01385 b | 0.05646 a | 0.01884 a | 0.00295 a |
| 1 | NP | 0.02975 a | NP | NP | 0.00235 a | |
| 2 | 0.00884 c | NP | NP | NP | 0.00123 a | |
| 3 | 0.00980 b | NP | NP | NP | 0.00027 a | |
Medias con la misma letra en una columna no son estadísticamente diferentes (Tukey, P ≤ 0.05). TZ: trans-zeatina, CZ: cis-zeatina, CIN: cinetina, AIA: ácido indolacético, ABA: ácido abscísico, NP: No presente.
En cuanto a la cinetina, solo se detectó en el megagametofito de las semillas del Tratamiento 1, sin que hubiera diferencias significativas con respecto al testigo; asimismo, se encontró AIA en el megagametofito de las semillas del Tratamiento 3 (Cuadro 3). En cuanto a la testa, solo se detectó trans-zeatina en las semillas de los Tratamientos 2 (escarificación con N2 + AG4/7 + estratificación fría + estratificación cálida) y 3 (escarificación con N2 + estratificación cálida, escarificación con N2 + AG4/7), siendo mayor en el Tratamiento 3. En contraste, la cis-zeatina se encontró en la testa de las semillas del Tratamiento 1, siendo dos veces mayor que en el testigo (0.02975 µg mg-1). El ABA estuvo presente en concentraciones bajas en la testa de las semillas de todos los tratamientos probados. Las hormonas que se encontraron en las concentraciones más altas en las semillas testigo (megagametofito más testa) fueron cinetina (0.0755 µg mg-1), cis-zeatina (0.0351 µg mg-1) y AIA (0.0365 µg mg-1) (Cuadro 3), en tanto que en el Tratamiento 1, en el que algunas semillas lograron romper la testa, se encontró dos veces más cis-zeatina (0.0297 µg mg-1) que trans-zeatina (0.0116 µg mg-1) y cinetina (0.0110 µg mg-1). Las hormonas que estuvieron presentes en mayor concentración, tanto en las semillas testigo como en las expuestas a todos los tratamientos probados, fueron las citocininas (trans-zeatina, cis-zeatina, cinetina) y las auxinas (AIA). No fue posible detectar ácido giberélico en las semillas al día cero, ni después de 120 días de aplicar los tratamientos.
Kucera et al. (2005) mencionaron que las citocininas promueven la división celular durante la formación y desarrollo del embrión. Las auxinas por su parte, definen el patrón basal-apical del embrión y permiten que el endospermo se celularice (Matilla, 2020). El ABA es una fitohormona requerida para la síntesis de carbohidratos, proteínas y lípidos de reserva (Bian et al., 2018). Esta hormona también regula la maduración, la morfogénesis y la tolerancia a la desecación del embrión, además de evitar la germinación prematura y promover la latencia (Matilla, 2000; Sano y Marion-Poll, 2021; Warpeha y Montgomery, 2016). Considerando lo anterior, se puede inferir que las semillas de T. globosa al día cero y después de 120 días de la aplicación de los tratamientos, aún se encontraban en la etapa de división y expansión celular, así como de acumulación de reservas (Matilla, 2000).
La baja concentración de ABA presente en las semillas al día cero y en todos los tratamientos probados, podría indicar que la latencia de las semillas de T. globosa después de 120 días de ser expuestas a distintos tratamientos no se debe únicamente al efecto inhibidor de esta hormona. Lo anterior contrasta con lo observado por Bian et al. (2018), quienes después de someter las semillas de T. yunnanensis a una combinación de estratificación fría (4 °C) y cálida (25 °C) encontraron que el ABA es el principal inhibidor de la germinación de las semillas en esta especie; asimismo, la presencia de mayor concentración de citocininas y auxinas en las semillas de T. globosa que de ABA, así como no haber detectado giberelinas (estimulan la germinación al aumentar su concentración), al tiempo cero y después de 120 días de haber aplicado los tratamientos para romper la latencia, sugiere que las semillas aún se encontraban en desarrollo como se mencionó anteriormente. De lo anterior, se puede inferir que morfológica y fisiológicamente, el embrión no estaba en condiciones de iniciar el proceso de germinación al momento de ser cosechadas las semillas, ni después de 120 días.
Fenoles
En el megagametofito de las semillas testigo (día cero) se encontró naringenina, floretina, ácido sinápico (ACS), ácido ferúlico (ACF) y ácido rosmarínico (ACR), siendo la naringenina la que se presentó en mayor concentración (0.0045 µg mg-1), en tanto que en la testa de las semillas testigo se detectó naringenina, floretina, ACS, ácido p-hidroxibenzoico (ACH), ACF y ACR (Cuadro 4). En cuanto a los tratamientos, la catequina se detectó en el megametofito de las semillas de los tratamientos 1, 2 y 3; en las semillas de los tratamientos 2 y 3 se encontró ACH, y ACS solo en las de los tratamientos 1 y 3 (Cuadro 4). Por otro lado, en la testa de los tratamientos 1, 2 y 3 fue posible observar naringenina, floretina, ACH, catequina y ácido protocatecuico (ACP). La catequina se encontró tanto en el megagametofito como en la testa de las semillas de todos los tratamientos probados, siendo significativamente mayor en el Tratamiento 1 (Cuadro 4). La mayor concentración de fenoles se encontró en la testa de las semillas, tanto del testigo como del resto de los tratamientos. Aun cuando en las semillas testigo y del Tratamiento 1 (en el que se observó rompimiento de la testa) se encontró naringenina, floretina y ACH, la concentración de estos fenoles se redujo en 50 % en las semillas del Tratamiento 1 (120 días después de aplicar los tratamientos) (Cuadro 4). Los fenoles que se detectaron en mayor concentración en las semillas testigo (megagametofito más testa) al día cero fueron naringenina (0.0111 µg mg-1), floretina (0.0095 µg mg-1) y ACS (0.00875 µg mg-1), y después de 120 días de aplicar los tratamientos fueron el ACP (0.0074 µg mg-1), catequina (0.0036-0.0053 µg mg-1), naringenina (0.0033-0.0045 µg mg-1) y ACH (0.0043-0.0044 µg mg-1).
Cuadro 4 Fenoles presentes en el megagametofito y testa de semillas de T. globosa sometidas a distintos tratamientos, después de 120 días.
| Trat | Estructura | NAR | FLO | ACS | ACH | ACF | ACR | CAT | ACP |
| (µg mg-1) | |||||||||
| Testigo | Megagametofito | 0.00459 ab | 0.00386 b | 0.00260 b | NP | 0.00136 b | 0.00216 a | NP | NP |
| 1 | NP | NP | 0.00227 c | NP | NP | NP | 0.00188 b | NP | |
| 2 | NP | NP | NP | 0.00222 b | NP | NP | 0.00163 c | NP | |
| 3 | NP | NP | 0.00234 bc | 0.00219 b | NP | NP | 0.00185 b | NP | |
| Testigo | Testa | 0.00656 a | 0.00564 a | 0.00614 a | 0.00487 a | 0.00393 a | 0.00682 a | NP | NP |
| 1 | 0.00456 ab | 0.00384 b | NP | 0.00222 b | NP | NP | 0.00344 a | 0.00747 a | |
| 2 | 0.00447 ab | 0.00377 c | NP | 0.00218 b | NP | NP | 0.00197 b | 0.00747 a | |
| 3 | 0.00335 b | 0.00377 c | NP | 0.00217 b | NP | NP | 0.00183 b | 0.00747 a | |
Medias con la misma letra en una columna no son estadísticamente diferentes (Tukey, P ≤ 0.05). NAR: naringenina, FLO: floretina, ACS: ácido sinápico, ACH: ácido p-hidroxibenzoico, ACF: ácido ferúlico, ACR: ácido rosmarínico, CAT: catequina, ACP: ácido protocatecuico, NP: no presente.
Los fenoles tienen un rol importante en las semillas, ya que participan en funciones fisiológicas como la latencia o viabilidad de éstas (Lepiniec et al., 2006). Algunos autores han reportado que los compuestos fenólicos como el ácido ferúlico, el AHB y el catecol son inhibidores de la germinación de las semillas de lechuga, nogal, algodón, melón, maíz y diversas malezas (Chenyin et al., 2023). Distintos autores señalan que la presencia de fenoles como taninos condensados (PAs), catequina, ácido ferúlico, ácido sinápico, AHB, así como las ligninas, son posibles inhibidores de la germinación de semillas de distintas especies (Bewley, et al., 2013; Chenyin et al., 2023; Lepiniec et al., 2006); asimismo, la naringenina fue reportada como inhibidora de la germinación en Glycine max y Arabidopsis(Hernández y Munné-Bosch, 2012), en tanto que el ácido rosmarínico se considera un compuesto de defensa en las plantas (Petersen y Simmonds, 2003) y el ácido protocatecuico tiene actividad alelopática (Šćepanović et al., 2022).
Taxus globosa es una especie forestal de distribución restringida, que produce un número limitado de semillas, las cuales presentan latencia al momento de ser dispersadas. Aunque T. globosa puede propagarse de manera vegetativa, la propagación mediante semillas es importante para los programas de restauración y mejoramiento genético. Lo anterior ha generado la necesidad de conocer los mecanismos que ocasionan la latencia de las semillas de esta especie, con el fin de diseñar las estrategias que permitan romper dicho periodo de reposo y promover la germinación del mayor número de semillas en el menor tiempo; no obstante, los trabajos enfocados a estudiar las posibles causas de la latencia en esta especie son escasos (Ramírez-Sánchez et al., 2018).
Los resultados obtenidos en la presente investigación indican que, efectivamente, las semillas de T. globosa presentan latencia morfofisiológica, como fue propuesto por Ramírez-Sánchez et al. (2018). Lo anterior se basa en el hecho de haber encontrado que el embrión de T. globosa aún estaba en una etapa de expansión celular y síntesis de sustancias de reserva al momento de la cosecha, lo que indica que éste no había alcanzado su madurez, pudiendo causar latencia morfológica; asimismo, fue posible detectar fenoles, considerados como inhibidores de la germinación, tanto en la testa como en el megagametofito, los cuales podrían actuar en forma sinérgica con el ABA para promover la latencia fisiológica. Estos hallazgos permitirán establecer de manera más certera, los tratamientos que promuevan la maduración del embrión y la eliminación de los inhibidores de la germinación, con el fin de reducir el tiempo requerido para que las semillas de T. globosa germinen.
Conclusiones
Los tratamientos probados para romper la latencia no lograron promover la germinación de las semillas. Sólo las semillas escarificadas con nitrógeno líquido lograron abrir su testa a los 90 días de incubación; este mismo tratamiento estimuló el crecimiento del embrión. Las semillas contienen otros inhibidores de la germinación (fenoles) en el embrión y el megagametofito (testa y megagametofito), además del ABA. La presencia de auxinas y citocininas en mayor concentración en las semillas al día cero y después de 120 días de aplicar los tratamientos, así como la ausencia de giberelinas, sugieren que las semillas de T. globosa aún se encontraban en desarrollo cuando fueron dispersadas. Las semillas de T. globosa requieren de más de 120 días después de la cosecha para que el embrión alcance la madurez, sea capaz de percibir los estímulos ambientales y logre germinar. La presencia de inhibidores de la germinación, así como de embriones inmaduros confirma que las semillas de la especie en estudio presentan latencia morfofisiológica.

