












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
PermalinkIntroducción
La degradación de los pastizales, causada principalmente por el sobrepastoreo, es un problema de importancia en México. A la fecha, se han perdido cerca de 6.5 millones de hectáreas de este bioma (INECC, 2015). Una alternativa para recuperar los pastizales degradados es la resiembra de pastos. En esta práctica lo ideal es utilizar vegetación nativa; no obstante, los programas de revegetación con este tipo de especies han sido poco exitosos debido a su limitada capacidad de establecimiento (Quero-Carrillo et al., 2016).
Los problemas de establecimiento de pastos nativos se deben a diferentes factores; por ejemplo, el mesocótilo crece para llevar al coleóptilo hasta la superficie del suelo, lo cual induce a que las raíces adventicias nazcan cerca de la superficie del suelo en condiciones desfavorables de humedad (Moreno-Gómez et al., 2012). La raíz primaria de especies nativas alcanza rápidamente el límite de su capacidad para captar agua después de la emergencia (Wilson et al., 1976); debido a esto, la tasa de transpiración de las hojas excede a la de captación de agua por la raíz en condiciones de sequía, lo cual resulta en la muerte de plántulas (Wilson et al., 1976). En este sentido, la proporción raíz-parte aérea también es un factor de importancia para compensar la pérdida de agua por transpiración debido a que entre mayor sea la proporción de raíces de una planta, más probabilidades tendrá de poder abastecer las necesidades de agua de la parte aérea (Freschet et al., 2015). Otra respuesta anatómica que presentan las plantas para disminuir la transpiración es reducir su área foliar (Monclus et al., 2006), y un factor adicional que influye sobre la pérdida de agua por transpiración son las características estomáticas (Lawson y Blatt, 2014). Los estomas son poros microscópicos localizados en la epidermis de las plantas, los cuales controlan el intercambio de agua y dióxido de carbono (CO2) entre la planta y la atmósfera (Lawson et al., 2014). Está documentado que la densidad estomática (DE) está positivamente correlacionada con la traspiración (Doheny-Adams et al., 2012; Tanaka et al., 2013). Con base en lo anterior, poblaciones con adaptaciones fenotípicas como mesocótilos cortos, proporción de raíces alta, área foliar menor y densidades estomáticas bajas podrían incrementar su capacidad de establecimiento y contribuir a incrementar el éxito de programas de revegetación de pastizales degradados con especies nativas.
El pasto banderita [Bouteloua curtipendula (Michx.) Torr.] es una de las especies nativas más utilizadas en rehabilitación de pastizales en el norte de México. Esta especie se reproduce principalmente de forma apomíctica, lo cual facilita la conservación de caracteres seleccionados a través de generaciones, lo que puede representar una ventaja en términos de mejoramiento genético; no obstante, ha mostrado una capacidad limitada de establecimiento en los programas de rehabilitación donde se ha utilizado. Investigaciones previas han demostrado que existe diversidad genética amplia dentro de esta especie en México (Morales et al., 2016; Morales-Nieto, 2006); sin embargo, se desconoce si esta especie posee diversidad fenotípica en cuanto a las características ligadas al establecimiento y si su comportamiento está relacionado con la cantidad de precipitación. Por lo anterior, el objetivo del presente estudio fue evaluar el desarrollo morfológico y las características estomáticas en plántulas de poblaciones de pasto banderita en dos condiciones de humedad que simulan los patrones de precipitación históricos de Chihuahua, y analizar su diversidad genética mediante marcadores AFLP para relacionar diferencias genéticas con las respuestas agronómicas en sequía simulada.
Materiales y métodos
Condiciones ambientales
El presente estudio se realizó en condiciones de invernadero durante 21 días, del 10 al 31 de agosto de 2019. Se evaluó el desarrollo de plántulas de diez poblaciones de pasto banderita en dos condiciones de humedad. La temperatura media durante el experimento fue de 26.5 ± 5.4 °C, con una mínima de 17.1 y máxima de 44.3 °C. La humedad relativa media fue de 52.5 ± 16.7 %, con una mínima de 20.5 y una máxima de 65.8 %.
Material genético
Se evaluaron nueve poblaciones originarias de Chihuahua, México (Cuadro 1), y como testigo se utilizó la variedad comercial Niner. La semilla utilizada fue producida y recolectada dos años antes del experimento (2017), a partir de plantas conservadas ex situ en el Rancho Experimental La Campana del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), donde previamente se había recolectado semilla de diferentes poblaciones, generado plantas y transplantado en campo.
Cuadro 1 Localización geográfica y condiciones ambientales del lugar de origen de las poblaciones de pasto banderita (Bouteloua curtipendula).
| Población/Variedad | Municipio | Longitud | Latitud | Altitud (m) | Precipitación media anual (mm) | Temperatura media anual (°C) |
| Niner | Socorro† | -106.893 | 34.057 | 1409 | 260 | 13.7 |
| HDP | Hidalgo del Parral | -105.668 | 27.016 | 1772 | 489 | 17.6 |
| SFO | San Francisco del Oro | -105.827 | 26.952 | 1870 | 491 | 16.3 |
| NCG | Nuevo Casas Grandes | -107.843 | 30.331 | 1526 | 316 | 16.6 |
| Janos | Janos | -107.845 | 31.244 | 1361 | 315 | 17.0 |
| Riva Palacio | Riva Palacio | -106.549 | 28.543 | 1835 | 432 | 14.6 |
| Namiquipa | Namiquipa | -107.402 | 29.248 | 1841 | 442 | 14.1 |
| Bachiniva | Bachiniva | -107.248 | 28.774 | 2038 | 450 | 13.8 |
| Ahumada | Ahumada | -106.386 | 29.820 | 1627 | 361 | 16.4 |
| Chihuahua | Chihuahua | -106.569 | 29.643 | 1713 | 320 | 16.6 |
†Ubicado en el estado de Nuevo México, Estados Unidos de América.
Tratamientos, diseño y unidad experimental
Los genotipos fueron sometidos a dos condiciones de humedad, obtenidas de dos de las secuencias de humedadsequía desarrolladas con la metodología propuesta por Esqueda et al. (2005) (Cuadro 2). Estas secuencias simulan dos de los patrones de precipitación pluvial más comunes en el mes de julio para el estado de Chihuahua, México (medio = normal y bajo = sequía). La evaluación se realizó con un diseño experimental de bloques completos al azar con arreglo factorial (poblaciones y niveles de sequía) con cinco repeticiones, donde los bloques se colocaron perpendicularmente a la trayectoria que siguió la radiación solar incidente durante el día. La unidad experimental consistió de una planta crecida en vaso de unicel.
Cuadro 2 Secuencias de riego utilizadas en el experimento para simular dos condiciones de humedad (normal y sequía) a través de dos patrones de precipitación de verano más comunes para el estado de Chihuahua, México.
| Patrón | Con | Sin | Con | Sin | Con | Sin | Con | Sin | Con | Sin | Con | Sin | Días con riego | Lamina de riego total (mm) |
| Sequía | 1 | 2 | 1 | 2 | 1 | 1 | 1 | 1 | 2 | 4 | 2 | 3 | 8 | 80 |
| Normal | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 4 | 2 | 1 | 10 | 100 |
Con: número de días con riego, Sin: número de días sin riego. La lamina de riego utilizada en cada evento fue de 10 mm (equivalente a un volumen de agua de 78.5 mL). Patrones tomados de Esqueda et al. (2005).
Manejo agronómico
La siembra se llevó a cabo en vasos de unicel de 473 mL, utilizando suelo de origen aluvial con textura francoarcillosa y pH de 8.5. Se prescindió del uso de fertilizante durante la evaluación debido a que no es una práctica que se realice en programas de rehabilitación de pastizales, para lo que se utiliza principalmente el pasto banderita. La siembra se realizó a una profundidad de 8 mm, sembrando aproximadamente 20 semillas (con cubiertas) por recipiente para asegurar emergencia; sin embargo, cinco días después de la emergencia se seleccionó la plántula más vigorosa en cada recipiente y se eliminó el resto para mantener únicamente una planta por recipiente.
Los riegos se realizaron manualmente, de manera lenta y controlada para no dañar las plantas durante la aplicación. En cada evento de riego se aplicó una lámina de agua de 10 mm sobre la superficie del suelo colocado en los vasos de unicel de 10 cm de diámetro. Esta lámina equivale a un volumen de 78.5 mL.
Evaluación fenotípica
La evaluación se realizó a los 21 días después de la emergencia, debido a que durante las primeras tres semanas es cuando se presenta la mayor tasa de mortalidad en pastos de zonas áridas (Wilson et al., 1976). El tamaño de muestra fue de cinco plantas por población y tratamiento de humedad. Los muestreos se hicieron por método destructivo; para ello, las plantas se extrajeron de los recipientes y se lavaron las raíces con agua corriente para remover el sustrato, cuidando de no perder raíces en el proceso. Las variables fenotípicas evaluadas fueron longitud de raíz seminal (LRS, cm) desde la semilla hasta la punta de las raíces seminales; longitud del mesocótilo (LDM, cm) desde la semilla hasta el inicio del coleóptilo; longitud acumulada de raíces adventicias (LARA, cm) como la sumatoria de la longitud de todas las raíces adventicias por planta; peso de raíces (PR, g) y peso de parte aérea (PPA, g) separando estos dos componentes en la base del coleóptilo y una vez separadas, fueron almacenadas en bolsas de papel y secadas en una estufa a 65 °C durante 72 h para posteriormente pesarlas en una balanza analítica (Viper BC, Mettler Toledo, Columbus, Ohio, EUA); proporción raíz-parte aérea; área foliar (AF, cm2) se estimó a través de fotografías digitales del total de las hojas de cada plántula, analizadas con ayuda del software ImageJ; eficiencia en el uso del agua (EUA) se calculó dividiendo la producción total de biomasa (raíz + parte aérea) entre el volumen total de riego utilizado en cada tratamiento de humedad; densidad estomática (DE, estomas mm-2) y área estomática (AE, μm2). La DE y el AE se evaluaron en tres hojas por planta tomando impresiones de la lámina foliar con pegamento líquido de cianoacrilato; para ello, se colocó una gota de pegamento en un portaobjetos, donde posteriormente se colocó la parte media de la hoja, se dejó secar por 2 min y se retiró. Las impresiones tomadas, de aproximadamente 1 cm, se observaron en microscopio de contraste de fases con un aumento de 400 diámetros (400X). Se tomaron fotografías de cinco campos ópticos de 0.0945 mm2 por muestra, seleccionados al azar; en cada campo óptico se contabilizó el número total de estomas y el valor obtenido se expresó por unidad de área (1 mm2). El área estomática se calculó con la información de tres estomas por campo óptico, seleccionados al azar. Esta medición se realizó con ayuda del software Zen Core versión 2. En total, en cada tratamiento de humedad se analizaron 75 campos ópticos y se midieron 225 estomas por población.
Evaluación genética
Las poblaciones fueron analizadas genéticamente a través de marcadores moleculares AFLP (Polimorfismos en la longitud de fragmentos amplificados), para verificar diferencias genotípicas entre poblaciones y evaluar su relación con las diferencias fenotípicas encontradas. Se colectaron hojas de cada uno de los individuos evaluados por población, las cuales fueron utilizadas para la extracción de ADN siguiendo el método descrito por Doyle y Doyle (1990). El análisis AFLP se llevó a cabo con el método propuesto por Vos et al. (1995). Las enzimas de restricción utilizadas fueron EcoRI y MseI, con un nucleótido extra en los iniciadores (EcoRI + A y MseI + A). Se utilizaron cuatro combinaciones de iniciadores marcados con fluorescencia: MseI + CTG - EcoRI + AAG, MseI + CTG - EcoRI + ACT, MseI + CAG - EcoRI + AGG y MseI + CAG - EcoRI + AAC. La separación de los fragmentos amplificados se hizo en un analizador de DNA LI-COR (LI-COR Biosciences, Lincoln, Nebraska, EUA) y el patrón de bandas obtenido fue utilizado para formar una matriz binaria de presencia (1) o ausencia (0) de éstas.
Análisis estadístico
Los datos de las respuestas fenotípicas fueron analizados mediante análisis de varianza (ANDEVA) y las medias comparadas mediante la prueba LSD (Fisher’s Least Significant Difference; Diferencia Mínima Significativa). El modelo estadístico para el análisis de los datos correspondió a un arreglo de dos factores (población y nivel de humedad). Los datos obtenidos en el tratamiento de sequía fueron comparados en un análisis de agrupamiento por el método de ligamento de Ward. Estos análisis fueron realizados con el software R estudio versión 4.1.1. Los datos genéticos fueron analizados por el método de agrupamiento UPGMA (Método del grupo de pares no ponderados con media aritmética) y el coeficiente Dice, con el software Past versión 4.03 (Hammer y Harper, 2001). Finalmente, las matrices de distancias genéticas y fenotípicas obtenidas de los análisis de agrupamiento (morfológico-fisiológico y genético) fueron utilizadas para realizar una prueba de Mantel y evaluar la relación entre la respuesta fenotípica de las poblaciones ante las condiciones de sequía simulada y las distancias genéticas que presentan entre sí.
Resultados y discusión
Los efectos de la población, patrón de precipitación y su interacción resultaron significativos (P ≤ 0.05) en todas las variables evaluadas, excepto en LRS (P > 0.05). Esta interacción significativa implica que ciertas poblaciones pueden responder de manera diferente en condiciones variables de precipitación, lo que sugiere que algunas poblaciones podrían estar mejor adaptados a situaciones de sequía, siendo esto valioso para programas de mejoramiento y rehabilitación de pastizales. La variedad Niner presentó mayor longitud acumulada de raíces adventicias (P ≤ 0.05) en ambos patrones de precipitación; sin embargo, fue de los más afectados (P ≤ 0.05), ya que su LARA se redujo 40.9 % en el tratamiento de sequía simulada con respecto al patrón medio. Por otro lado, la población de Riva Palacio fue la que obtuvo menor LARA en ambos tratamientos de humedad (P ≤ 0.05; Figura 1).
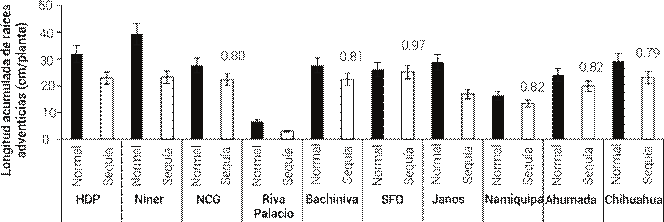
Figura 1 Longitud acumulada de raíces adventicias de 10 poblaciones de pasto banderita (Bouteloua curtipendula) en dos condiciones de humedad, simuladas a través de dos patrones de precipitación pluvial de verano más comunes en el estado de Chihuahua, México(Esqueda et al., 2005). Los números arriba de las barras representan la proporción de la condición de sequía con respecto a la condición normal de humedad. Los asteriscos indican diferencias significativas entre tratamientos de humedad (P ≤ 0.05). HDP: Hidalgo del Parral, NCG: Nuevo Casas Grandes, SFO: San Francisco del Oro. Las líneas verticales representan ± el error estándar de la media.
Las semillas de pastos nativos son capaces de emerger rápido y en porcentaje alto después de una resiembra de pastizales. No obstante, la mayoría de las plántulas emergidas comúnmente mueren durante las primeras semanas de vida, en la etapa de establecimiento (James et al., 2011; Lauenroth et al., 1994). La muerte de plántulas ha sido asociada con la transición de las estructuras radicales cuando las raíces seminales mueren y dan paso a las raíces adventicias (Briske y Wilson, 1977; Torres y Villalobos, 2024). Las plántulas que no son capaces de desarrollar un sistema funcional de raíces adventicias mueren, debido a la capacidad limitada que tienen las raíces seminales para captar agua (Larson et al., 2015; Wilson et al., 1976). Por esta razón, el desarrollo de raíces adventicias en condiciones de sequía es fundamental para el establecimiento de las plántulas. En este sentido, la población de Riva Palacio puede tener desventajas en establecimiento, ya que fue la que obtuvo la LARA menor en ambos tratamientos de humedad. Las poblaciones más afectadas por la reducción de humedad fueron Janos con 55.1 % y Riva Palacio con 41.3 % en disminución de su LARA. La variedad Niner también fue de las más afectadas con 40.9 % de reducción de LARA. Debido a esto, dichas poblaciones también podrían presentar desventajas en el establecimiento.
Respecto a la LDM, la población de Namiquipa fue la única que presentó diferencias (P ≤ 0.05) y fue la de mayor LDM en ambos patrones (Figura 2); no obstante, la LDM no varió entre niveles de humedad (P > 0.05) para ninguna de las poblaciones. Una limitante para el establecimiento del pasto banderita es que es de tipo panicoide (crecimiento de verano). Los pastos tipo panicoide y los de tipo festucoide (crecimiento en temporada fría) presentan importantes diferencias estructurales en su sistema radical, mismas que afectan el establecimiento. En ambos tipos de pastos la raíz primaria es de vida corta y debe ser remplazada por las raíces adventicias, las cuales parten del mesocótilo (Newman y Moser, 1988); no obstante, en pastos panicoides el mesocótilo se encuentra cerca de la superficie del suelo, por lo que las raíces adventicias crecen en condiciones desfavorables de humedad (Hyder et al., 1971). Por esta razón, poblaciones con mesocótilos cortos pueden desarrollar raíces adventicias a mayor profundidad del suelo en condiciones de humedad más favorables. En este sentido, la población de Namiquipa puede tener desventajas de establecimiento, debido a que presentó la longitud mayor del mesocótilo en ambos patrones de precipitación. En el resto de las poblaciones no hubo diferencias significativas entre sí.
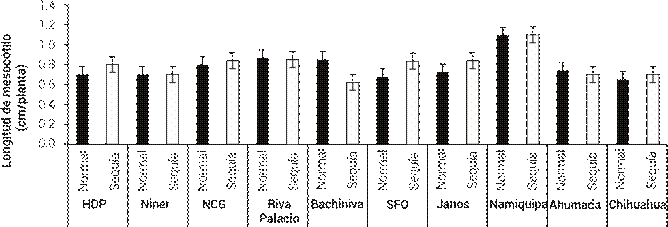
Figura 2 Longitud de mesocótilo de 10 poblaciones de pasto banderita (Bouteloua curtipendula) en dos condiciones de humedad simuladas a través de dos patrones de precipitación pluvial de verano comunes en el estado de Chihuahua, México(Esqueda et al., 2005). HDP: Hidalgo del Parral, NCG: Nuevo Casas Grandes, SFO: San Francisco del Oro. Las líneas verticales representan ± el error estándar de la media.
La producción de biomasa de las poblaciones presentó un comportamiento variable ante la reducción de humedad (Figura 3). Las poblaciones más afectadas en producción de biomasa fueron Niner, San Francisco del Oro y Janos, ya que su biomasa se redujo en 56, 47 y 44 %, respectivamente, al comparar los dos tratamientos de humedad. Por otro lado, la producción de biomasa de las poblaciones Nuevo Casas Grandes y Namiquipa no disminuyó en la condición de sequía; sin embargo, su proporción raízparte aérea sí presentó un cambio (P ≤ 0.05). El peso de raíces de estas poblaciones se incrementó en 64 y 47.7 %, mientras que su biomasa área se redujo en 62.5 y 18.0 %, respectivamente. Por esta razón, la proporción raízparte aérea se incrementó a más del triple en la población Nuevo Casas Grandes y 2.75 veces en la de Namiquipa. El incremento en peso de raíces que presentaron estas poblaciones en condiciones de sequía concuerda con lo reportado por Mansoor et al. (2015). Estos investigadores encontraron que ecotipos de pasto buffel (Pennisetum ciliare) tolerantes al estrés hídrico producen una cantidad de raíces mayor cuando se desarrollan en condiciones de sequía. Por otra parte, el incremento en peso de raíces de las poblaciones Nuevo Casas Grandes y Namiquipa podría reflejarse en una capacidad mayor para adquirir nutrientes y agua del suelo. Así mismo, el incremento en la proporción de raíz-parte aérea de estas poblaciones puede incidir en una mayor capacidad de establecimiento debido a que a mayor proporción de raíces-parte aérea de una planta, mayor es la capacidad de abastecer las necesidades de agua de la parte aérea y sobrevivir durante una sequía (Freschet et al., 2015).
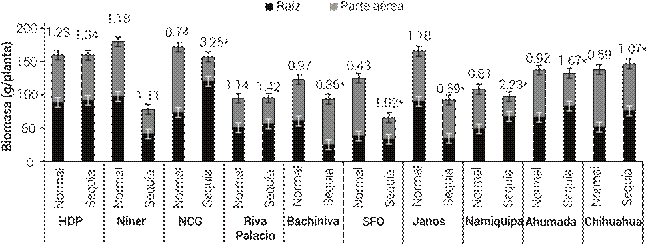
Figura 3 Producción de biomasa radical y de parte aérea de 10 poblaciones de pasto banderita (Bouteloua curtipendula) en dos condiciones de humedad simuladas a través de dos patrones de precipitación pluvial de verano comunes en el estado de Chihuahua, México(Esqueda et al., 2005). Los números arriba de las barras representan el coeficiente raíz/ parte aérea, para cada una de las condiciones (normal y sequía). Los asteriscos indican diferencias significativas entre tratamientos de humedad (ANOVA, P ≤ 0.05). HDP: Hidalgo del Parral, NCG: Nuevo Casas Grandes, SFO: San Francisco del Oro. Las líneas verticales representan ± el error estándar de la media.
En general, el AF de todas las poblaciones evaluadas disminuyó (P ≤ 0.05) en el tratamiento de sequía, con respecto al tratamiento de humedad normal (Figura 4) debido a que las plantas tienden a disminuir su superficie foliar en condiciones de estrés hídrico, como una respuesta para disminuir la pérdida de agua por transpiración (Monclus et al., 2006); no obstante, la magnitud de la disminución varió entre poblaciones (P ≤ 0.001). Las poblaciones que mostraron menor disminución fueron Niner (23.3 %), San Francisco del Oro (29.8 %) y Janos (31.1 %); en contraste, las poblaciones de Nuevo Casas Grandes, Namiquipa, Ahumada y Chihuahua, además de incrementar su proporción raíz-parte aérea, disminuyeron su área foliar en 51, 54, 49 y 59 %, respectivamente en la condición de sequía, lo que indica la presencia de mecanismos de adaptación a tal condición.
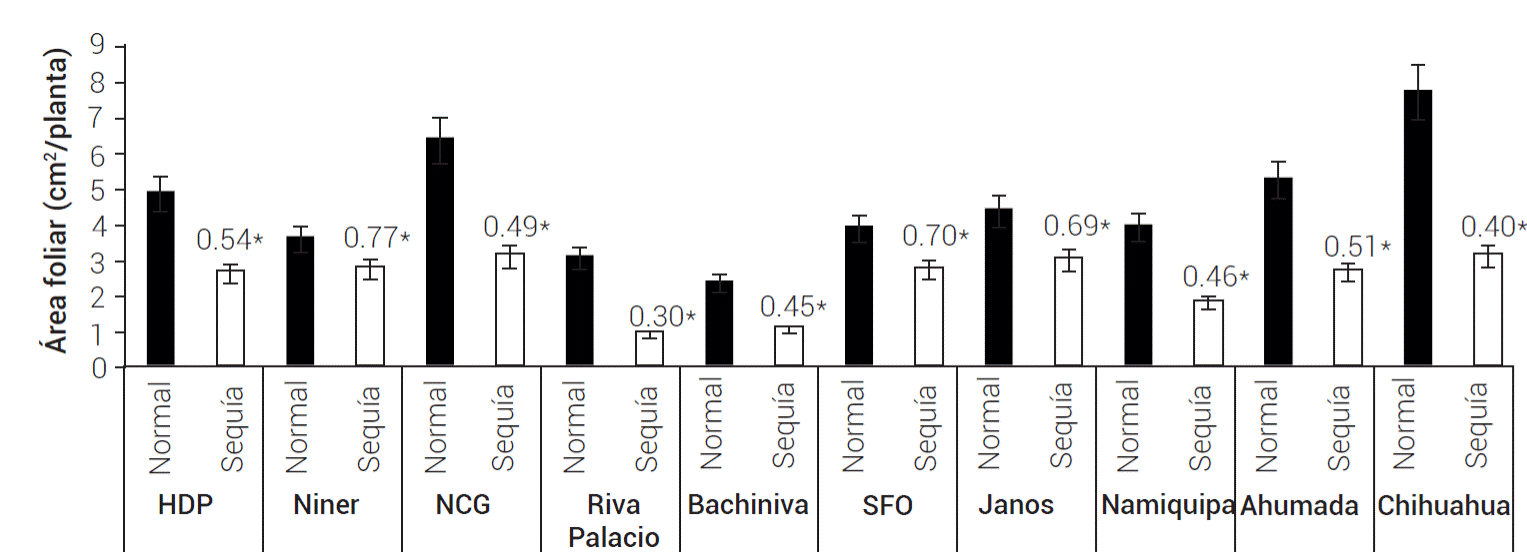
Figura 4 Área foliar de 10 poblaciones de pasto banderita (Bouteloua curtipendula) en dos condiciones de humedad simuladas a través de dos patrones de precipitación pluvial de verano comunes en el estado de Chihuahua, México(Esqueda et al., 2005). Los números arriba de las barras representan la proporción de la condición de sequía con respecto a la condición normal de humedad. Los asteriscos indican diferencias significativas entre tratamientos de humedad (ANDEVA, P ≤ 0.05). HDP: Hidalgo del Parral, NCG: Nuevo Casas Grandes, SFO: San Francisco del Oro. Las líneas verticales representan ± el error estándar de la media.
Las variables DE y AE también presentaron una respuesta variable al estrés por sequía. La mayoría de las poblaciones presentaron una disminución (P ≤ 0.05) en la DE del haz cuando fueron sometidos a condiciones de sequía. En el envés, cinco poblaciones presentaron un incremento (P ≤ 0.05), dos una disminución (P ≤ 0.05) y tres no mostraron cambios (P > 0.05) en DE cuando estuvieron en sequía. Por otro lado, el AE de la mayoría de las poblaciones mostró un incremento (P ≤ 0.05) de entre 8 y 53% en la condición de sequía, tanto en el haz como en el envés (Cuadro 3), lo que concuerda con Hong et al. (2018), quienes observaron que las plantas pueden modificar su densidad estomática como medida adaptativa a las condiciones ambientales.
Cuadro 3 Características estomáticas de 10 poblaciones de pasto banderita (Bouteloua curtipendula) en dos condiciones de humedad simuladas a través de dos patrones de precipitación pluvial de verano comunes en el estado de Chihuahua, México(Esqueda et al., 2005).
| Población/variedad | Densidad estomática (estomas mm-2) | Área estomática (μm2) | ||||||
| Haz | Envés | Haz | Envés | |||||
| Normal | Sequía | Normal | Sequía | Normal | Sequía | Normal | Sequía | |
| Niner | 143cd | 156a | 154cd | 171b* | 242cd | 249ef | 294bc | 281d |
| Hidalgo del Parral | 144cd | 125b* | 161bc | 149c* | 217de | 257def* | 278cd | 286d* |
| San Francisco del Oro | 141cd | 152a | 157bc | 175b* | 270bc | 270def | 318ab | 336bc |
| Nuevo Casas Grandes | 146cd | 176a* | 166bc | 184ab * | 198e | 241f* | 248de | 293d* |
| Janos | 139d | 149.2 a | 146cd | 169b* | 271bc | 275cde | 305bc | 311cd |
| Riva Palacio | 170ab | 154a* | 175ab | 205a* | 201e | 309bc* | 219e | 306cd* |
| Namiquipa | 160abc | 152a | 147cd | 177b* | 269bc | 280cd | 301bc | 307cd |
| Bachiniva | 151bcd | 121bc* | 146cd | 147c | 285ab | 323b* | 329ab | 357ab* |
| Ahumada | 175a | 148a* | 195a | 198a | 213de | 261def* | 241e | 291d* |
| Chihuahua | 139d | 111c* | 137d | 132d | 303a | 362a* | 343a | 378a* |
Letras en superíndice diferentes por columna indica diferencias significativas entre poblaciones (DMS, P ≤ 0.05). * Indica diferencias (P ≤ 0.05) entre condiciones de humedad, detectadas por análisis de varianza.
La densidad y área de los estomas son parámetros importantes en la eco-fisiología de las plantas, ya que controlan la pérdida de agua por transpiración y la captura de CO2(Lawson y Blatt, 2014; Lawson et al., 2014). Estudios previos han encontrado que la densidad estomática correlaciona positivamente con la transpiración (Doheny-Adams et al., 2012; Franks et al., 2015; Tanaka et al., 2013), y densidades estomáticas bajas pueden favorecer la eficiencia en el uso del agua, asimilación de CO2 y actividad fotosintética en pastos (Bertolino et al., 2019); además, pastos con estomas grandes pueden presentar menor transpiración en comparación con plantas de estomas pequeños (Xu y Zhou, 2008); por lo tanto, especies o poblaciones con densidades estomáticas bajas pueden ser más tolerantes a sequía que aquellas con mayor cantidad de estomas (Caine et al., 2019; Dunn et al., 2019; Hughes et al., 2017); esto concuerda con Monzón-Burgos et al. (2023), quienes encontraron que poblaciones de pasto banderita con pocos estomas con gran área estomática tuvieron mayor capacidad de establecimiento en campo en comparación con poblaciones de muchos estomas con área estomática pequeña. En este sentido, la población de Chihuahua puede tener ventajas adaptativas y mayor establecimiento en campo debido a que fue la que presentó menor densidad y mayor área de estomas en ambos tratamientos de humedad. La Figura 5 ilustra las diferencias en DE y AE entre algunas de las poblaciones, en haz y envés.

Figura 5 Microestructura foliar del haz y el envés de plántulas de tres genotipos de pasto banderita (Bouteloua curtipendula) en dos condiciones de humedad, simuladas a través de dos de los patrones de precipitación pluvial de verano comunes en el estado de Chihuahua, México(Esqueda et al., 2005). Las líneas blancas representan el contorno de los estomas y los números el área estomática.
La eficiencia en el uso del agua (EUA) no varió entre poblaciones (P > 0.05) en el patrón de precipitación normal, pero sí (P ≤ 0.01) en el tratamiento de sequía (Cuadro 4). La tolerancia a sequía es un factor crítico en el establecimiento y crecimiento de los pastos, sobre todo en regiones áridas o semiáridas. Esta puede definirse como la capacidad de las plantas para mantener su metabolismo y producción de biomasa en condiciones de disponibilidad de agua limitada (Jones et al., 2015), y puede caracterizarse por una mayor eficiencia en el uso del agua (Mårtensson et al., 2017). Por esta razón, los pastos que son eficientes en el uso del agua suelen ser más resistentes a la sequía debido a que pueden mantener su crecimiento y funciones vitales incluso por períodos prolongados de escasez de agua, lo que los hace más adecuados para la revegetación en áreas con patrones de precipitación erráticos (Mganga et al., 2021). En este sentido, el uso de poblaciones de pastos con EUA alta puede contribuir a incrementar la probabilidad de éxito de los programas de restauración de pastizales degradados (Kiniry y Kim, 2020; Mganga et al., 2021). Las poblaciones Nuevo Casas Grandes, Ahumada y Chihuahua podrían presentar una capacidad mayor de establecimiento en programas de revegetación debido a que mostraron mayor EUA (P ≤ 0.05) en el tratamiento de sequía.
Cuadro 4 Eficiencia en el uso del agua de 10 poblaciones de pasto banderita (Bouteloua curtipendula) en dos condiciones de humedad simuladas a través de dos patrones de precipitación pluvial de verano comunes en el estado de Chihuahua, México(Esqueda et al., 2005).
| Población/variedad | Eficiencia en el uso del agua (mg mL-1) | |
| Normal | Sequía | |
| Hidalgo del Parral | 2.34 a | 3.11 ab |
| Niner | 2.01 a | 2.31 bc |
| San Francisco del Oro | 1.05 a | 2.25 bc* |
| Nuevo Casas Grandes | 1.65 a | 4.09 a* |
| Janos | 2.20 a | 2.06 bc |
| Riva Palacio | 1.51 a | 1.31 c |
| Namiquipa | 1.18 a | 2.41 bc* |
| Bachiniva | 1.93 a | 1.63 bc |
| Ahumada | 1.72 a | 2.89 ab* |
| Chihuahua | 1.82 a | 3.03 ab* |
Letras diferentes por columna indican diferencias significativas entre poblaciones (P ≤ 0.05). * Indica diferencias (P ≤ 0.05) entre condiciones de humedad, detectadas por análisis de varianza.
La sequía simulada generó un incrementó en la EUA (P ≤ 0.05) de cinco de las poblaciones; en el resto de ellas la EUA no varió (P > 0.05). Las poblaciones en las que se incrementó la EUA se caracterizaron por ser de las de mayor longitud acumulada de raíces adventicias, proporción raíz-parte aérea y área estomática en el tratamiento de sequía; además, mostraron área foliar y densidad estomática menor en comparación con el resto de las poblaciones. Estos resultados sugieren que las variables longitud acumulada de raíces adventicias, proporción raízparte aérea, área foliar, densidad y área estomática son importantes para el uso eficiente del agua y pueden ser utilizadas como criterio de selección de poblaciones de pastos para revegetación de pastizales.
De acuerdo con el análisis de AFLPs, las poblaciones analizadas presentaron diversidad genética alta, ya que el 56 % de los loci analizados fueron polimórficos. El patrón de agrupamiento con los datos genéticos fue similar al presentado en los resultados de la evaluación fenotípica en condiciones de sequía (Figura 6); además, la prueba de Mantel reveló una correlación débil (r = 0.36) pero significativa (P ≤ 0.05) entre las distancias genéticas y las fenotípicas. Estos resultados sugieren que las diferencias genéticas influyeron parcialmente sobre las diferencias en las respuestas fenotípicas de las poblaciones ante las condiciones de sequía simulada; a su vez, esto sustenta la hipótesis de que las poblaciones de pasto banderita analizadas presentan una respuesta fenotípica diferente en las condiciones de sequía y ésta se relaciona con diferencias genéticas. La correlación débil entre los datos de AFLPs y los fenotípicos puede deberse a que los marcadores AFLP toman muestras de todo el genoma aleatoriamente y no fragmentos genómicos con una función conocida sobre características de interés (Harris et al., 2010).
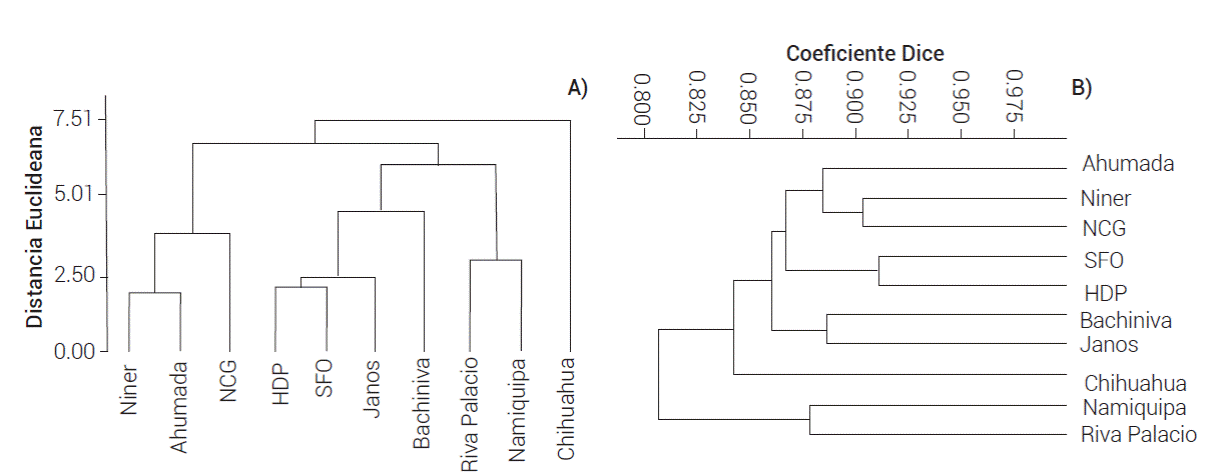
Figura 6 Dendrogramas de agrupamiento de 10 poblaciones de pasto banderita (Bouteloua curtipendula). A) Elaborado con la información de 10 variables fenotípicas evaluadas en condiciones de sequía simulada y método de ligamento Ward. B) Elaborado con la información de 186 fragmentos de AFLPs y el método de agrupamiento UPGMA.
Conclusiones
El desarrollo de plántulas y las características estomáticas de poblaciones de pasto banderita varía dependiendo del nivel de humedad. La correlación significativa entre las distancias fenotípicas y genéticas sugiere que las diferencias en la respuesta fenotípicas de las poblaciones ante las condiciones de sequía simulada fueron están relacionadas con sus diferencias genéticas. Los resultados obtenidos indican que una respuesta favorable ante el estrés hídrico está determinada por varios factores en pasto banderita. Las poblaciones que pudieran estar más adaptadas al estrés hídrico incrementan su proporción raíz-parte aérea, área estomática y uso eficiente del agua cuando se desarrollan en condiciones de sequía. Asimismo, dichos poblaciones disminuyen su área foliar y densidad estomática. En este estudio, las poblaciones que presentaron este comportamiento fueron las originarias de Nuevo Casas Grandes, Ahumada y Chihuahua, México. Por lo tanto, estas poblaciones tienen potencial para convertirse en nuevas variedades de pasto banderita y ser utilizadas en programas de revegetación de pastizales degradados o para reconvertir áreas agrícolas a ganaderas.

