












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
México es un centro destacado de diversidad para el género Pinus L.; en su territorio se distribuyen de manera natural 48 de sus especies (Sosa-Díaz et al., 2024), que constituyen cerca del 45% de las existentes en el mundo (Farjon, 1996). Los pinos habitan y dominan diversos tipos de vegetación a lo largo de los principales sistemas montañosos del país (Gernandt y Pérez-De La Rosa, 2014), por lo que representan un recurso relevante económica y ecológicamente (Moctezuma-López y Flores, 2020). Sin embargo, en las últimas décadas los bosques de pino han experimentado alteraciones notables y el deterioro de su hábitat (Sáenz-Ceja et al., 2022), lo que evidencia la necesidad de su conservación y uso sostenible. Para ello, se requiere generar información de las asociaciones ecológicas y su relación con el ambiente, lo que a su vez permitirá identificar, dentro de este género, grupos funcionales (Sánchez-González, 2008).
Un grupo funcional es un conjunto de especies que comparten rasgos biológicos y tienen una función similar en el ecosistema (Saldaña-Acosta et al., 2008). La clasificación de las especies en grupos funcionales optimiza la comprensión de los mecanismos de funcionamiento en ecosistemas con alta diversidad biológica (Lujan-Soto et al., 2015) y puede servir como guía para mantener la estructura natural de los ecosistemas forestales (Sandoval-García et al., 2020). En el contexto del manejo forestal, la agrupación de especies puede contribuir a una silvicultura mejorada y una planeación más efectiva de las actividades forestales (Lujan-Soto et al., 2015).
Las relaciones ecológicas entre especies, y de estas con su entorno, juegan un papel crucial en la composición de las comunidades vegetales (Estrada-Castillón et al., 2015). Adicionalmente, las interacciones entre estas relaciones y los filtros ambientales que operan a escalas local, regional y continental son determinantes para la distribución espacial y la diversidad de las especies (Sánchez-González y López-Mata, 2005; Arenas-Navarro et al., 2020b). Por otro lado, la comprensión de las relaciones ecológicas entre especies y de estas con el ambiente es esencial para predecir cómo responderán las comunidades vegetales a cambios ambientales.
En este contexto, las cuadrículas espaciales (CE) son un enfoque de análisis estructurado que subdivide el área de estudio en unidades geométricas regulares (celdas), para facilitar la organización, el análisis y la evaluación de factores ambientales asociados a la distribución de las especies (Arenas-Navarro et al., 2020b). El uso de CE para identificar patrones ecológicos, combinado con el método de Análisis de Correspondencia Canónica (ACC), método exploratorio de datos multivariados que emplea técnicas de ordenación y regresión para detectar relaciones no lineales entre especies y variables ambientales (ter Braak, 1989; McCune, 1997), proporciona una herramienta poderosa para comprender cómo las especies responden a las condiciones ambientales en áreas determinadas.
En México se han realizado estudios sobre la ordenación de comunidades vegetales con presencia de pinos a nivel de sitio y en gradientes altitudinales (Sánchez-González y López-Mata, 2003; Estrada-Castillón et al., 2015; Arenas-Navarro et al., 2020a). Aunque existen estudios que identifican grupos de especies de Pinus con afinidades estructurales (p. ej., Luján-Soto et al., 2015), resulta fundamental ahondar en el conocimiento sobre el ensamblaje ambiental que afecta la distribución y determinar los factores que ejercen mayor influencia en el patrón geográfico de estas especies. Para contribuir a llenar este hueco de conocimiento, y tomando como objeto de estudio las especies del género Pinus que se distribuyen de forma natural en México, esta investigación se propuso alcanzar dos objetivos: 1) contribuir al conocimiento sobre la similitud ecológica en la distribución de especies, y 2) determinar los factores ambientales (derivados de temperatura, precipitación, suelo y atmósfera) que mayor influencia tienen sobre la distribución espacial de las especies; ambos a nivel de paisaje en las principales regiones montañosas de México.
Materiales y Métodos
Registro de especies y área de estudio
Se recopiló una base de datos con las coordenadas geográficas de localidades en las que se reportó la presencia de pinos en México. Esta información se obtuvo de las siguientes fuentes: 1) bases de datos en línea (Global Biodiversity Information Facility (GBIF, 2024), Sistema Nacional de Información sobre Biodiversidad de México (CONABIO, 2024), SEINet Portal Network (SEINet, 2024), como motores de búsqueda web Google Scholar (Google, 2024), Web of Science (WoS, 2024) y Scopus (Scopus, 2024), y 2) literatura científica asociada al tema empleando las palabras clave: “longitud”, “latitud”, “coordenadas”, “Pinus”, “México”, en español e inglés.
Debido a que la taxonomía de Pinus se encuentra en constante actualización, los registros reunidos se homogeneizaron a nivel especie con base en el índice de nombres científicos de Farjon (2005), complementando con estudios recientes sobre determinación de especies (Pérez De La Rosa y Gernandt, 2017; Montes et al., 2019, 2022; Buck et al., 2020, 2023). Por otro lado, se excluyeron los datos ubicados en uso de suelo no forestal o fuera del rango altitudinal de cada especie según lo reportado por Farjon y Filer (2013) y Farjon (2017), además de los duplicados en 1 km2 (por especie). Los registros retenidos se acotaron a los principales sistemas montañosos (SM) de México (Fig. 1): Sierra Madre Occidental (SMOc), Sierra Madre Oriental (SMOr), Faja Volcánica Transmexicana (FVT), Sierra Madre del Sur (SMS), Sierras de Chiapas (SMCh) y Sierras de Baja California (SBC). La delimitación de estas regiones se realizó con base en las provincias fisiográficas de México propuestas por Morrone et al. (2017). La SMCh y SBC son espacialmente equivalentes a la Provincia de los Altos de Chiapas y la Provincia Californiana de Morrone et al. (2017).
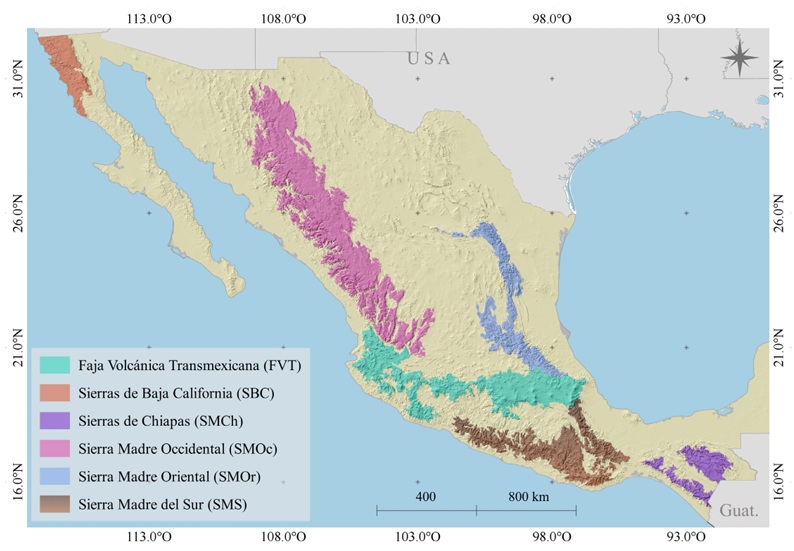
Figura 1: Provincias fisiográficas de México. La delimitación de los sistemas montañosos estudiados aquí se basó en las ecorregiones propuestas por Morrone et al. (2017).
Ordenación de las variables ambientales y presencia de especies
El medio físico del área de estudio se caracterizó a nivel pixel de ~1 km2 de resolución espacial con datos climáticos, atmosféricos, topográficos y de suelo obtenidos de las plataformas WorldClim (Hijmans et al., 2005) y SoilGrids (Hengl et al., 2014). Las variables utilizadas en el análisis se describen en el Cuadro 1.
Con base en el área ocupada por los registros de ocurrencia de especies de pino se construyó una cuadrícula espacial (CE) de ~10 km por lado. Esta fue empleada para generar una matriz ambiental y dos matrices de especies para cada SM. La primera registró el promedio de cada variable ambiental por celda. Las matrices de especies fueron de tipo binario (presencia-ausencia, 1 - 0, respectivamente) y de abundancia espacial (número de registros de una especie en cada celda de 10 × 10 km). Además, de acuerdo con Arenas-Navarro et al. (2020b) se calculó un indicador de calidad de celdas (Cc), el cual consistió en dividir la riqueza observada (conteo de especies por celda) entre la riqueza estimada (calculada con base en el estimador no paramétrico de Chao 2) por celda. El Cc varía de cero a uno, donde un valor cercano a uno indica celdas con calidad suficiente, mientras que un valor cercano a cero indicaría lo contrario. Las celdas con Cc ≤ 0.5 en cada SM se excluyeron en análisis subsecuentes.
Cuadro 1 Variables ambientales usadas para caracterizar el medio físico de los principales sistemas montañosos de México. 1=Información obtenida de WorldClim (Hijmans et al., 2005); 2=Información obtenida de SoilGrids (Hengl et al., 2014).
| Nombre de la variable | Tipo | Símbolo | Unidades |
| Precipitación anual | Precipitación1 | b12 | mm |
| Precipitación del mes más lluvioso | Precipitación1 | b13 | mm |
| Precipitación del mes más seco | Precipitación1 | b14 | mm |
| Estacionalidad de la precipitación | Precipitación1 | b15 | mm |
| Precipitación del trimestre más húmedo | Precipitación1 | b16 | mm |
| Precipitación del trimestre más seco | Precipitación1 | b17 | mm |
| Precipitación del trimestre más cálido | Precipitación1 | b18 | mm |
| Precipitación del trimestre más frío | Precipitación1 | b19 | mm |
| Temperatura media anual | Temperatura1 | b1 | °C |
| Rango diurno medio de la temperatura | Temperatura1 | b2 | °C |
| Estacionalidad de la temperatura | Temperatura1 | b4 | °C |
| Temperatura máxima del mes más cálido | Temperatura1 | b5 | °C |
| Temperatura mínima del mes más frío | Temperatura1 | b6 | °C |
| Rango anual de la temperatura | Temperatura1 | b7 | °C |
| Temperatura media del trimestre más húmedo | Temperatura1 | b8 | °C |
| Temperatura media del trimestre más seco | Temperatura1 | b9 | °C |
| Temperatura media del trimestre más cálido | Temperatura1 | b10 | °C |
| Temperatura media del trimestre más frío | Temperatura1 | b11 | °C |
| Orientación de la pendiente | Topografía1 | exp | ° de azimut |
| Altitud | Topografía1 | alt | msnm |
| Pendiente del terreno | Topografía1 | pen | % |
| Radiación solar | Atmósfera1 | rad | KJ m-2 día-1 |
| Presión de vapor de agua | Atmósfera1 | vap | kPa |
| Velocidad del viento | Atmósfera1 | vel | m s-1 |
| Densidad aparente del suelo | Suelo2 | dap | kg m-3 |
| Capacidad de intercambio catiónico | Suelo2 | cic | cmolc kg-1 |
| Contenido de carbono orgánico | Suelo2 | cos | ‰ |
| Partículas de arcilla en el suelo | Suelo2 | arc | % |
| Partículas de limo en el suelo | Suelo2 | lim | % |
| Partículas de arena en el suelo | Suelo2 | are | % |
| Potencial de hidrogeno | Suelo2 | pH |
Análisis estadístico: Relación especies-ambiente y entre especies
Para el análisis de los datos se utilizó el paquete de análisis ecológico de comunidades (Community Ecology Package) vegan v. 2.6-4 (Oksanen et al., 2022) del programa R (R Core Team, 2022). Inicialmente, para evitar redundancia de parámetros ambientales se realizó un proceso de reducción de variables mediante la función “ordistep” Choose a Model by Permutation Tests in Constrained Ordination (dirección=“forward”, permutaciones=500) del paquete vegan versión 2.6-4 (Oksanen et al., 2022) del programa R (R Core Team, 2022). Este proceso tuvo mayor efecto en la SBC y SMCh donde redujo hasta un 60% de las variables iniciales.
Posteriormente, a partir del conjunto de variables resultantes se implementaron dos versiones de análisis de correspondencia canónica (ACC; ter Braak, 1989) para identificar las variables ambientales que más influyen sobre la distribución espacial de los pinos en cada SM. El ACC emplea datos ambientales y de especies (matriz de presencia-ausencia o matriz de abundancia espacial en cada celda) y los relaciona mediante ejes canónicos, permitiendo interpretar de manera sencilla la relación entre ambos elementos (Borcard et al., 2018). En la versión uno del ACC se empleó la matriz ambiental y la matriz de presencia-ausencia (ACCPA); en la versión dos se empleó la matriz ambiental y la matriz de abundancia espacial (ACCAA). Ambos ACC se realizaron mediante la función “cca” Correspondence Analysis and Redundancy Analysis del paquete vegan versión 2.6-4 (Oksanen et al., 2022) de R (R Core Team, 2022).
Para cada sistema montañoso se evaluó un ACCAP y ACCAA y se eligió el de mayor inercia explicada, con respecto a la inercia total, para representar la relación especies-ambiente. Para el ACC elegido se ejecutaron pruebas de permutación (1000 repeticiones) para conocer el número de ejes y variables significativas (p<0.05). A partir de estas pruebas se seleccionaron las diez variables significativas (p<0.05) con mayor aporte a la inercia explicada por el ACC, mismas que fueron consideradas las más influyentes sobre el patrón espacial de las especies de pino en cada SM. Posteriormente, con base en las coordenadas de las especies representadas en un diagrama de ordenación derivado del ACC, se definió un número óptimo de grupos (clusters) mediante el método de agrupación “kmeans” y la función “fviz_nbclust” Determining and Visualizing the Optimal Number of Clusters del paquete “factoextra” v. 1.0.7 (Kassambara y Mundt, 2020) del programa R (R Core Team, 2022). Las especies incluidas en cada grupo se consideraron elementos biológicos cuya distribución espacial posee rasgos ecológicos similares (Sánchez-González y López-Mata, 2003; Saldaña-Acosta et al., 2008). Estos grupos de especies representan una aproximación a la definición de grupos funcionales de especies de pino en cada SM.
Resultados
Distribución y registro de especies de pino en los SM
La base final de registros de especies de pino de los SM se conformó por 46 especies (Apéndice 1) y 20,525 registros (registros no repetidos por especie dentro de 1 km2). Pinus leiophylla Schiede ex Schltdl. & Cham. y P. pseudostrobus Lindl. tuvieron el número mayor de registros (3335 y 2160, respectivamente). Contrariamente, Pinus radiata D. Don y P. vallartensis Pérez de la Rosa tuvieron solo nueve y ocho registros, respectivamente. La distribución de registros por SM fue de 31.6% para la SMOc, 26.5% para la FVT, 18.3% para la SMOr, 12.3% para la SMS, 6.1% para la SMCh y 5.3% para la SBC, con 23, 27, 24, 17, 12 y 9 especies, respectivamente.
El SM con mayor número de celdas (~10 × 10 km) fue la SMOc (1605) mientras que la SBC tuvo el menor con 170. El Cc (indicador de calidad de celdas) causó que se excluyeran del análisis 32, 21, 11, 11 y 3 celdas para la SMOc, FVT, SMS, SMOr y SMCh, respectivamente (Cc≤0.5). Para las celdas retenidas, el promedio (±1 Ds) del valor de Cc fue de 0.89 (±0.15) para la FVT, 0.91 (±0.14) para SMS, 0.92 (±0.15) para la SMOc, 0.94 (±0.12) para SMCh y SMOr y 0.96 (±0.19) para las SBC. En el mismo orden, el Cc mínimo fue 0.60, 0.57, 0.54, 0.57, 0.60 y 0.67, respectivamente para cada SM.
Influencia de variables ambientales en la distribución de pinos
Todos los análisis canónicos realizados fueron significativos (F=6.8 a 16.3; p=0.001; Apéndice 2). Los ACCPA lograron explicar porcentualmente el 13.3, 17.5, 18.8, 21.3, 22.2 y 39.1 de la inercia total para la SMOc, FVT, SMS, SMOr, SMCh y SBC, respectivamente. En el mismo orden, los ACCAA explicaron porcentualmente el 17.7, 23.8, 24.1, 29.9, 28.7 y 57.6 de la inercia total. Por tanto, los ACC más adecuados para identificar grupos funcionales y la relación especies-ambiente fueron los basados en la matriz ambiental y de abundancia espacial (ACCAA). Al menos cuatro ejes canónicos de cada SM resultaron significativos (p<0.05, Apéndice 3). En promedio, los dos primeros ejes explicaron el 49.5% de la inercia y hasta 72.8% por los cuatro primeros. El menor porcentaje de inercia explicada por los primeros dos ejes canónicos fue para la FVT y SMOr (35.6%), mientras que para la SMCh y SBC ocurrió lo contrario (63.1 y 69.8%, respectivamente).
La altitud, la velocidad del viento, la radiación solar, la presión de vapor de agua y la precipitación fueron las variables ambientales que determinaron en mayor medida la distribución de pinos en los SM estudiados (Apéndice 4, Cuadro 2). La presión de vapor de agua (vap) explicó el mayor porcentaje de inercia en la SMOc y la SMS (19.7% y 32.6%, respectivamente). La precipitación del mes más lluvioso (b13) fue crucial en la SMOr (17.9%) y la estacionalidad de la precipitación (b15) en la SBC (32.5%). La altitud destacó en las SMCh (38.5%), mientras que la temperatura media del cuarto más seco (b9) fue relevante en la FVT (17.2%) (Apéndice 4, Cuadro 2).
La correlación (r) entre los factores ambientales y los ejes canónicos del ACCAA indicaron que el clima, el suelo y la topografía son impulsores importantes de la distribución geográfica de los pinos en México (Apéndice 5). En los SM del sur (SMCh y SMS), la temperatura se relacionó de forma negativa con el eje 1 y de forma contraria para el resto de SM. Contrariamente, la precipitación no mostró algún patrón claro y se relacionó de forma positiva y negativa con ambos ejes canónicos, mostrándose la mayor magnitud positiva con el eje uno de la SBC y la SMOr (r=0.9), mientras que en la SMOc y FVT se observó la mayor relación negativa (r=-0.7). Por su cuenta, los factores edáficos tuvieron la mayor relación negativa con el eje 1 de la SMOr (r=-0.8) y la mayor positiva en la SMS (r=0.7). Para el resto de SM, la relación entre el suelo y los ejes canónicos varió de |r| = 0.1 a 0.4. Los parámetros atmosféricos tuvieron coeficientes de mayor magnitud con el eje uno (|r promedio|= 0.5), observándose valores del orden de r=0.8 (SBC y SMOc) y r=0.9 (SMCh y SMS). A nivel de variable, la altitud (alt) y la velocidad del viento (vel) fueron los parámetros que incidieron en todos los SM. Ambas se relacionaron fuertemente con el eje uno en cada SM, aunque la mayor magnitud se observó en el caso de la primera (|r|= 0.3 a 0.9).
Asociación de especies y ambiente
A partir de los diagramas de ordenación de los ACCAA y el procedimiento de agrupación "kmeans" fue posible identificar al menos cuatro grupos de especies de pino con características ecológicas relativamente similares y su relación con los gradientes ambientales en cada SM.
En la SMOc, el grupo de especies conformado por P. devoniana Lindl., P. oocarpa Schiede ex Schltdl. y P. praetermissa Styles & McVaugh (Fig. 2, color verde) fue el más diferenciado por el ACCAA, y responde principalmente a la presión de vapor (vap), pero negativamente a velocidad del viento (vel). En el mismo SM, P. maximartinezii Rzed., P. remota (Little) D.K. Bailey & Hawksw. y P. pinceana Gordon fueron agrupadas y asociadas a suelos con alto contenido de arcilla (arc) y pH (Fig. 2, color naranja). Especies como P. leiophylla, P. strobiformis Engelm., P. durangensis Martínez y P. teocote Schiede ex Schltdl. & Cham.) fueron poco representados por la varianza explicada por los primeros dos ejes del ACCAA de la SMOc (Fig. 2, color rosa).
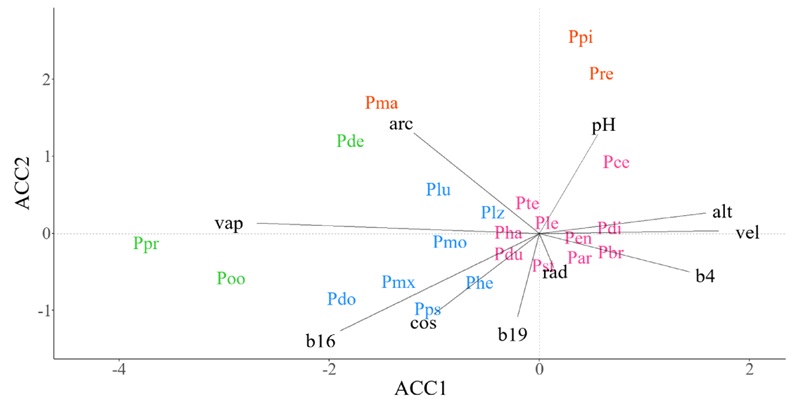
Figura 2: Diagrama de ordenación que resulta del Análisis de Correspondencia Canónica (ACCAA) entre especies de pino y promedio de las variables ambientales en celdas de ~10 × 10 km en la Sierra Madre Occidental (SMOc). Especies: Par=P. arizonica Engelm., Pbr=P. brachyptera Engelm., Pce=P. cembroides Zucc., Pde=P. devoniana Lindl., Pdi=P. discolor D.K. Bailey & Hawksw., Pdo=P. douglasiana Martínez, Pdu=P. durangensis Martínez, Pen=P. engelmannii Carrière, Pha=P. hartwegii Lindl., Phe=P. herrerae Martínez, Ple=P. leiophylla Schiede ex Schltdl. & Cham., Plz=P. lumholtzii B.L. Rob. & Fernald, Plu=P. luzmariae Pérez de la Rosa, Pma=P. maximartinezii Rzed., Pmx=P. maximinoi H. E. Moore, Pmo=P. montezumae Lamb., Poo=P. oocarpa Schiede ex Schltdl., Ppi=P. pinceana Gordon, Ppr=P. praetermissa Styles & McVaugh, Pps=P. pseudostrobus Lindl., Pre=P. remota (Little) D.K. Bailey & Hawksw., Pst=P. strobiformis Engelm., Pte=P. teocote Schiede ex Schltdl. et Cham. Variables ambientales: alt=altitud (m s.n.m.), arc=partículas de arcilla (%), b16=precipitación del trimestre más húmedo (mm), b19=precipitación del trimestre más frío (mm), b4=estacionalidad de la temperatura (°C), cos=contenido de carbono orgánico (‰), pH=potencial de hidrogeno, rad=radiación solar (KJ m-2 día-1), vap=presión de vapor de agua (kPa), vel=velocidad del viento (m s-1). El color en las especies indica la pertenencia de estas a un grupo funcional dentro del sistema montañoso.
En la SMOr, el grupo de especies que incluye a P. patula Schiede ex Schltdl. & Cham., P. ayacahuite Ehrenb. ex Schltdl., P. chiapensis (Martínez) Andresen y P. oocarpa se distribuyen conforme a la presencia de mayor precipitación en el mes más lluvioso (b13) (Fig. 3, color rosa). El grupo de pinos piñoneros (P. cembroides Zucc., P. pinceana, P. johannis M.-F. Robert, P. discolor D.K. Bailey & Hawksw., P. nelsonii Shaw y P. remota (Fig. 3, color naranja) responde a suelos con densidad aparente intermedia-alta (dap). Las especies P. hartwegii Lindl., P. culminicola Andresen & Beaman, P. arizonica Engelm., P. strobiformis y P. engelmannii Carrière (Fig. 3, color verde) se asociaron a ambientes de elevada altitud (alt) y velocidad del viento (vel). Por el contrario, la distribución del grupo encabezado por P. pseudostrobus y P. montezumae Lamb. (Fig. 3, color azul) fue poco explicado por gradientes ambientales específicos de la SMOr.
Cuadro 2: Factores ambientales y su contribución (%) a explicar la distribución de pinos en los sistemas montañosos de México según el Análisis de Correspondencia Canónico basado en datos de abundancia de especies de pino (ACCAA) en celdas espaciales de 10 × 10 km. El orden de los valores es: porcentaje de contribución al ACCAA (valor mínimo, valor máximo registrado en celdas de cada sistema montañoso de México). FVT=Faja Volcánica Transmexicana; SBC=Sierras de Baja California; SMCh=Sierras de Chiapas; SMOc=Sierra Madre Occidental; SMOr=Sierra Madre Oriental; SMS=Sierra Madre del Sur; b3=isotermalidad ((b2/b7) (×100)); b5=temperatura (T) máxima del mes más cálido (°C); b6=T mínima del mes más frío (°C); b8=T media del trimestre más húmedo (°C); b9=T media del cuarto más seco (°C); b10=T media del trimestre más cálido (°C); b11=T media del trimestre más frío (°C); b7=rango anual de T (b5-b6) (°C); b4=estacionalidad de la T (sd×10); b2=rango diurno medio de T (°C); b13=Precipitación (P) del mes más lluvioso (mm/10); b14=P del mes más seco (mm); b16=P del trimestre más húmedo (mm/10); b18=P del trimestre más cálido (mm/10); b19=P del trimestre más frío (mm/10); b15=estacionalidad de la precipitación (C.V./10); dap=densidad aparente del suelo (kg m−3); cos=carbono orgánico en el suelo (‰); arc=contenido de arcilla (%); alt=altitud (m s.n.m./10); pen=pendiente del terreno (%); rad=radiación solar (KJ m-2 día-1); vel=velocidad del viento (m s-1); vap=presión de vapor de agua (kPa). Significancia de la variable: *=0.001, §=0.003.
| Var | FVT | SBC | SMCh | SMOc | SMOr | SMS |
| Temperatura | ||||||
| b2 | 5.1* (9.9-19.3) | |||||
| b3 | 3.8* (58.1, 84.5) | |||||
| b4 | 6.8* (31.2, 67.5) | 7.1* (23.4, 70.9) | ||||
| b5 | 4.8* (14.8, 38.2) | |||||
| b6 | 6.9* (-2.8, 6.3) | 6.2* (2.8, 17.7) | 7.9* (0.9, 12.0) | |||
| b7 | 3.5* (15.5, 29.8) | |||||
| b8 | 3.1* (6.8, 27.7) | 8.3* (12.9, 26.7) | ||||
| b9 | 17.2* (5.8, 26.4) | 9.3* (9.7, 20.9) | ||||
| b10 | 3.4* (7.8, 28.6) | 3.2§ (13.2, 27.7) | 5.4* (10.1, 27.9) | |||
| b11 | 3.7* (8.1, 19.5) | 7.4* (7.3, 24.6) | ||||
| Precipitación | ||||||
| b13 | 3.9* (1.9, 8.9) | 17.9* (3.5, 50.1) | 3.0* (10.1, 71.1) | |||
| b14 | 10.6* (0.9, 72.2) | 5.3* (0, 2.1) | ||||
| b15 | 32.5* (5.5, 9.3) | |||||
| b16 | 2.4* (18.1, 92.6) | |||||
| b18 | 9.3* (15.2, 106.2) | 16.1* (0.9, 11.6) | 4.3* (19.1, 92.1) | |||
| b19 | 6.5* (2.0, 20.9) | 4.7* (2.1, 20.1) | ||||
| Suelo | ||||||
| dap | 5.3* (12.5, 15.1) | 12.6* (12, 14.6) | ||||
| cos | 11.6* (3.8, 34.2) | 2.0§ (16.1, 54.1) | ||||
| arc | 4.3* (16.3, 40.7) | 3.2§ (28.6, 43.1) | 2.4* (27.6, 49.0) | 3.0* (32.3, 45.6) | ||
| pH | 4.0* (5.3, 8.3) | 4.6* (5.4, 8.1) | ||||
| Topografía | ||||||
| alt | 6.4* (9.9, 405.3) | 7.1* (8.7, 239.2) | 38.5* (39.2, 279.2) | 2.6* (14.6, 290.6) | 5.8* (7.7, 283.3) | 2.2* (12.5, 330) |
| pen | 3.5§ (1.0, 23.6) | |||||
| Atmósfera | ||||||
| rad | 5.3* (18.4, 19.3) | 9.1* (16.9, 18.8) | 9.2* (17.1, 19.5) | 3.1* (16.1, 19.3) | ||
| vel | 5.1* (1.2, 4.4) | 3.9* (2.2, 3.4) | 3.4§ (1.8, 3.2) | 5.1* (1.4, 3.6) | 6.6* (1.3, 5.3) | 2.9* (1.5, 3.3) |
| vap | 12.1* (1.1, 2.4) | 19.7* (0.6, 2.1) | 32.6* (0.8, 2.4) | |||
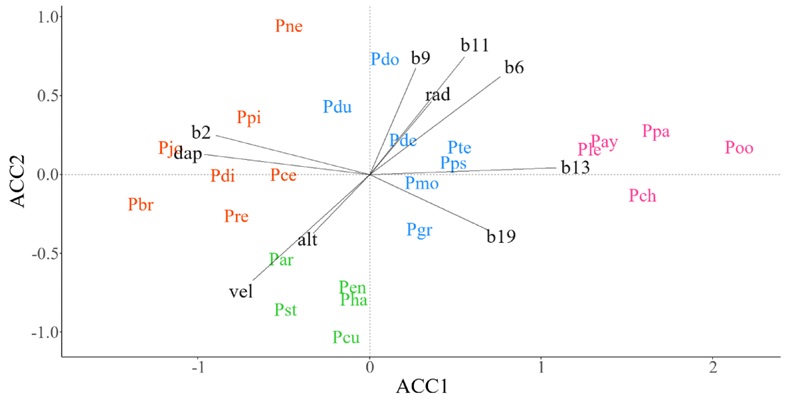
Figura 3: Diagrama de ordenación que resulta del Análisis de Correspondencia Canónica (ACCAA) entre datos de abundancia espacial (ACCAA) de especies de pino y valores ambientales promedio en celdas de ~ 10 × 10 km en la Sierra Madre Oriental (SMOr). Especies: Par=P. arizonica Engelm., Pay=P. ayacahuite Ehrenb. ex Schltdl., Pbr=P. brachyptera Engelm., Pce=P. cembroides Zucc., Pch=P. chiapensis Martínez, Pcu=P. culminicola Andresen & Beaman, Pde=P. devoniana Lindl., Pdi=P. discolor D.K. Bailey & Hawksw., Pdo=P. douglasiana Martínez, Pdu=P. durangensis Martínez, Pen=P. engelmannii Carrière, Pgr=P. greggii Engelm. ex Parl., Pha=P. hartwegii Lindl., Pjo=P. johannis M.-F.Robert, Ple=P. leiophylla Schiede ex Schltdl. & Cham., Pmo=P. montezumae Lamb., Pne=P. nelsonii Shaw, Poo=P. oocarpa Schiede ex Schltdl., Ppa=P. patula Schiede ex Schltdl. & Cham., Ppi=P. pinceana Gordon, Pps=P. pseudostrobus Lindl., Pre=P. remota (Little) D.K. Bailey & Hawksw., Pst=P. strobiformis Engelm., Pte=P. teocote Schiede ex Schltdl. et Cham. Variables: alt=altitud (msnm), b11=temperatura media del trimestre más frío (°C), b13=precipitación del mes más lluvioso (mm), b19=precipitación del trimestre más frío (mm), b2=rango diurno medio de la temperatura (°C), b6=temperatura mínima del mes más frío (°C), b9=temperatura media del trimestre más seco (°C), dap=densidad aparente del suelo (kg m-3), rad=radiación solar (KJ m-2 día-1), vel=velocidad del viento (m s-1). El color en las especies indica la pertenencia de estas a un grupo funcional.
En la FVT se identificaron cuatro grupos de especies con preferencias ecológicas relativamente similares (Fig. 4). El ACCAA claramente expuso un grupo liderado por P. jaliscana Pérez de la Rosa (Fig. 4, color naranja) y otro por P. maximinoi H.E. Moore (Fig. 4, color azul), los cuales se asociaron a temperaturas y precipitaciones elevadas en periodos cálidos (b10, b5, b8, b9, b18). Dentro del grupo de especies que incluye a P. cembroides, P. patula y P. chiapensis (Fig. 4, color rosa), se identificaron asociaciones particulares claras. Por ejemplo, la primera especie estuvo claramente asociada a valores altos de pH del suelo, mientras que P. chiapensis y P. patula se asociaron a ambientes con niveles altos de precipitación del mes más seco (b14; Fig. 4, color rosa). De forma particular, la distribución de P. leiophylla y P. pseudostrobus no mostraron una clara preferencia hacia un gradiente ambiental en particular (Fig. 4, color rosa).
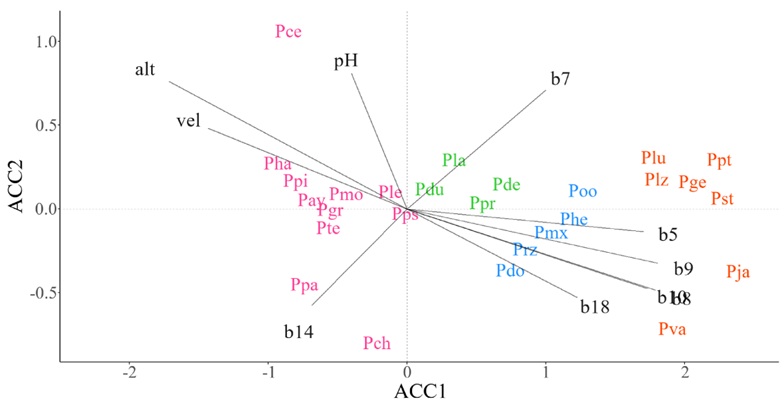
Figura 4: Diagrama de ordenación que resulta del Análisis de Correspondencia Canónica (ACCAA) entre datos de abundancia espacial de especies de pino y valores ambientales promedio en celdas de ~10 × 10 km en la Faja Volcánica Transmexicana (FVT). Especies: Pay=P. ayacahuite Ehrenb. ex Schltdl., Pce=P. cembroides Zucc., Pch=P. chiapensis Martínez, Pde=P. devoniana Lindl., Pdo=P. douglasiana Martínez, Pdu=P. durangensis Martínez, Pge=P. georginae Pérez de la Rosa, Pgr=P. greggii Engelm. ex Parl., Pha=P. hartwegii Lindl., Phe=P. herrerae Martínez, Pja=P. jaliscana Pérez de la Rosa, Pla=P. lawsonii Roezl ex Gordon, Ple=P. leiophylla Schiede ex Schltdl. & Cham., Plz=P. lumholtzii B.L. Rob. & Fernald, Plu=P. luzmariae Pérez de la Rosa, Pmx=P. maximinoi H. E. Moore, Pmo=P. montezumae Lamb., Poo=P. oocarpa Schiede ex Schltdl., Ppa=P. patula Schiede ex Schltdl. & Cham., Ppi=P. pinceana Gordon, Ppt=P. praetermissa Styles & McVaugh, Ppr=P. pringlei Shaw, Pps=P. pseudostrobus Lindl., Prz=P. rzedowskii Madrigal & M. Caball., Pst=P. strobiformis Engelm., Pte=P. teocote Schiede ex Schltdl. et Cham., Pva=P. vallartensis Pérez de la Rosa. Variables ambientales: alt=altitud (msnm), b10=temperatura media del trimestre más cálido (°C), b14=precipitación del mes más seco (mm), b18=precipitación del trimestre más cálido (mm), b5=temperatura máxima del mes más cálido (°C), b7=rango anual de la temperatura (°C), b8=temperatura media del trimestre más húmedo (°C), b9=temperatura media del trimestre más seco (°C), pH=potencial de hidrogeno, vel=velocidad del viento (m s-1). El color en las especies indica la pertenencia de estas a un grupo funcional.
En la Sierra Madre del Sur, el grupo formado por P. ayacahuite, P. hartwegii y P. patula fue claramente discriminado por el análisis (Fig. 5, color azul). Este contingente de especies se distribuye en áreas con abundante carbono orgánico en el suelo (cos) y donde la velocidad del viento (vel) es alta. Dentro del grupo conformado por P. maximinoi, P. chiapensis y P. oocarpa (Fig. 5, color rosa), las primeras dos se asociaron a elevada precipitación en periodos lluviosos (b13), mientras que la última fue estrechamente ligada a ambientes con alta presión de vapor de agua (vap) y temperaturas elevadas en el trimestre más cálido (b10) y frío (b11) del año. Dentro del mismo SM, P. leiophylla, P. teocote, P. montezumae, P. cembroides y P. pseudostrobus fue un grupo de especies (Fig. 5, color verde) relacionados con ambientes altitudinalmente intermedios. Por otro lado, el grupo de especies que incluyó a P. devoniana y P. pringlei Shaw (Fig. 5, color naranja) estuvo asociado a niveles altos de densidad aparente (dap) y porcentaje de arcilla en el suelo (arc).
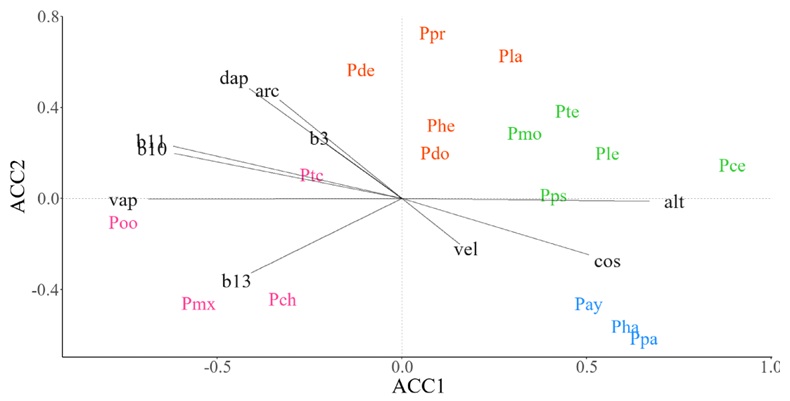
Figura 5 Diagrama de ordenación que resulta del Análisis de Correspondencia Canónica (ACCAA) basado en datos de abundancia espacial (ACCAA) de especies de pino y promedio de variables ambientales en celdas de ~10 × 10 km en la Sierra Madre del Sur (SMS). Especies: Pay=P. ayacahuite Ehrenb. ex Schltdl., Pce=P. cembroides Zucc., Pch=P. chiapensis Martínez, Pde=P. devoniana Lindl., Pdo=P. douglasiana Martínez, Pha=P. hartwegii Lindl., Phe=P. herrerae Martínez, Pla=P. lawsonii Roezl ex Gordon, Ple=P. leiophylla Schiede ex Schltdl. & Cham., Pmx=P. maximinoi H. E. Moore, Pmo=P. montezumae Lamb., Poo=P. oocarpa Schiede ex Schltdl., Ppa=P. patula Schiede ex Schltdl. & Cham., Ppr=P. pringlei Shaw, Pps=P. pseudostrobus Lindl., Ptc=P. tecunumanii F. Schwerdtf. ex Eguiluz & J.P. Perry, Pte=P. teocote Schiede ex Schltdl. et Cham. Variables ambientales: alt=altitud (msnm), arc=partículas de arcilla (%), b10=temperatura media del trimestre más cálido (°C), b11=temperatura media del trimestre más frío (°C), b13=precipitación del mes más lluvioso (mm), b3=isotermalidad (°C), cos=contenido de carbono orgánico (‰), dap=densidad aparente del suelo (kg m-3), vap=presión de vapor de agua (kPa), vel=velocidad del viento (m s-1). El color en las especies indica la pertenencia de estas a un grupo funcional.
En las Sierras de Chiapas se identificaron cinco grupos de especies de pino. El grupo mejor discriminado por el Análisis de Correspondencia Canónica lo constituyeron P. maximinoi, P. teocote y P. oocarpa, que responden a valores medios de temperatura (b8, b6 y b10), presión de vapor de agua (vap) y altitudes menores (alt) (Fig. 6, color azul). La especie P. hartwegii fue la única integrante de su grupo y su distribución se asoció a zonas con alta incidencia de radiación solar (rad) (Fig. 6, color naranja). Por otro lado, la altitud (alt) fue la variable que mejor caracterizó la distribución del grupo conformado por P. patula, P. montezumae, P. ayacahuite y P. pseudostrobus (Fig. 6, color rosa). En la SMCh, la distribución de P. chiapensis y P. douglasiana mostró mayor relación a sitios con alto contenido de arcilla (arc) y velocidad del viento (vel) (Fig. 6, color verde). Las especies P. devoniana y P. tecunumanii se asociaron con niveles intermedios de precipitación en el trimestre más cálido (b18) del año (Fig. 6, color morado).
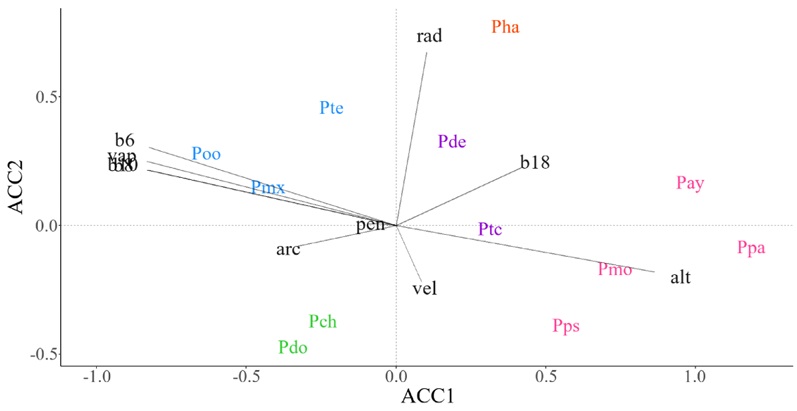
Figura 6: Diagrama de ordenación que resulta del Análisis de Correspondencia Canónica (ACCAA) basado en datos de abundancia espacial de especies de pino y promedio de variables ambientales en celdas de ~10 × 10 km en la Sierras de Chiapas (SMCh). Especies: Pay=P. ayacahuite Ehrenb. ex Schltdl., Pch=P. chiapensis Martínez, Pde=P. devoniana Lindl., Pdo=P. douglasiana Martínez, Pha=P. hartwegii Lindl., Pmx=P. maximinoi H. E. Moore, Pmo=P. montezumae Lamb., Poo=P. oocarpa Schiede ex Schltdl., Ppa=P. patula Schiede ex Schltdl. & Cham., Pps=P. pseudostrobus Lindl., Ptc=P. tecunumanii F. Schwerdtf. ex Eguiluz & J.P. Perry, Pte=P. teocote Schiede ex Schltdl. et Cham. Variables ambientales: alt=altitud (msnm), arc=partículas de arcilla (%), b10=temperatura media del trimestre más cálido (°C), b18=precipitación del trimestre más cálido (mm), b6=temperatura mínima del mes más frío (°C), b8=temperatura media del trimestre más húmedo (°C), pen=pendiente del terreno (%), rad=radiación solar (KJ m-2 día-1), vap=presión de vapor de agua (kPa), vel=velocidad del viento (m s-1). El color en las especies indica la pertenencia de estas a un grupo funcional.
En la SBC, el grupo conformado por P. jeffreyi Balf., P. lambertiana Douglas, P. californiarum D. K. Bailey, P. quadrifolia Parl. ex Sudw. y P. contorta Douglas ex Loudon (Fig. 7, color naranja) mostró poca preferencia por algún gradiente ambiental específico. Mientras que la distribución de P. radiata y P. muricata D. Don se caracterizó por tener las mayores precipitaciones en el mes más seco (b14) y suelos con alto contenido de arcilla (arc) (Fig. 7, color verde). Así mismo, P. coulteri D. Don (Fig. 7, color azul) y P. attenuata Lemmon (Fig. 7, color rosa) fueron las únicas especies de sus respectivos grupos.
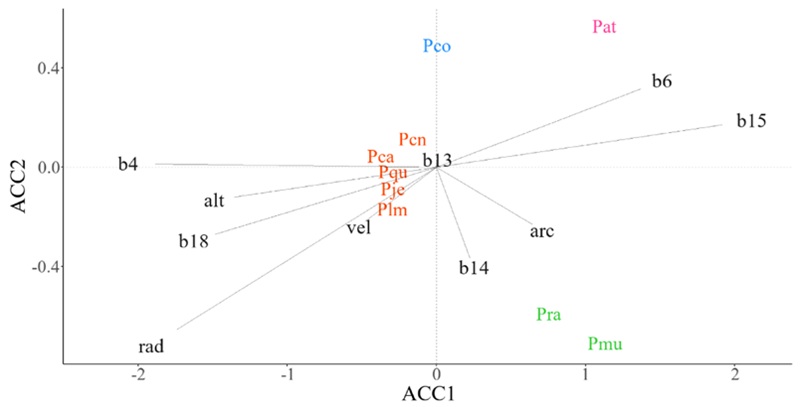
Figura 7: Diagrama de ordenación que resulta del Análisis de Correspondencia Canónica (ACCAA) basado en datos de abundancia espacial de especies de pino y promedio de variables ambientales en celdas de ~10 × 10 km en la Sierras de Baja California (SBC). Especies: Pat=P. attenuata Lemmon, Pca=P. californiarum D. K. Bailey, Pcn=P. contorta Douglas ex Loudon, Pco=P. coulteri D. Don, Pje=P. jeffreyi Balf., Plm=P. lambertiana Douglas, Pmu=P. muricata D. Don, Pqu=P. quadrifolia Parl. ex Sudw., Pra=P. radiata D. Don. Variables ambientales: alt=altitud (m s.n.m.), arc=partículas de arcilla (%), b13=precipitación del mes más lluvioso (mm), b14=precipitación del mes más seco (mm), b15=estacionalidad de la precipitación (mm), b18=precipitación del trimestre más cálido (mm), b4=estacionalidad de la temperatura (°C), b6=temperatura mínima del mes más frío (°C), rad=radiación solar (KJ m-2 día-1), vel=velocidad del viento (m s-1). El color en las especies indica la pertenencia de estas a un grupo funcional.
Discusión
Especies estudiadas
La composición (o estructura florística) de los ecosistemas con presencia de especies de Pinus en los SM de México, coincide con sus áreas descritas en estudios biogeográficos en México (Farjon, 1996; Sánchez-González, 2008; Sáenz-Ceja et al., 2022; Sosa-Díaz et al., 2024). Sin embargo, el número de especies registradas difiere al de otros estudios, p. ej., para las SBC se registraron nueve especies, resultado que coincide con Farjon (1996), pero difiere de las 10 mencionadas por Sáenz-Ceja et al. (2022) y las 19 mencionadas por Sánchez-González (2008). Tales diferencias se deben a que la taxonomía del género se encuentra en continua evolución, empleando nuevas y mejores técnicas para la determinación de especies. En esta investigación se consideraron como válidas las especies reconocidas por Farjon (2005) y se complementaron con estudios filogenéticos recientes (Pérez De La Rosa y Gernandt, 2017; Montes et al., 2019, 2022; Buck et al., 2020, 2023).
Desempeño del análisis canónico
La inercia explicada por los Análisis de Correspondencia Canónica apoyados en datos de abundancia (ACCAA) fue, en general, baja (30.3% en promedio). Las varianzas acumuladas fueron de 49.5 y 72.8% para los dos y cuatro primeros ejes canónicos. Este hallazgo es típico para este tipo de análisis, ya que las especies pueden responder de forma diferente a las variables ambientales (ter Braak, 1989; Borcard et al., 2018). Sin embargo, análisis similares realizados por Galindo-Jaimes et al. (2002) en rodales con disturbio dominados por especies de Pinus en Chiapas (México), explicaron 51% (dos ejes) de la varianza total de los datos ambientales y de especies, lo que es claramente superior a la inercia explicada en nuestro estudio. Para la FVT, en este estudio se obtuvo un valor de varianza explicada de 35.5%, el cual es también sustancialmente menor a 52 y 49.9% de varianza explicada para comunidades de pino-encino evaluadas a nivel de sitio en el occidente (Jalisco) y en un gradiente altitudinal del centro de México (Estado de México) reportadas por Arenas-Navarro et al. (2020a) y Sánchez-González y López-Mata (2003), respectivamente.
Es importante tener presente que la variación en los datos ambientales es dependiente del tamaño de las celdas espaciales. Aun así, se ha comprobado que el ACC tiene un desempeño robusto (ter Braak, 1989; McCune, 1997; Guisan et al., 1999) y proporciona información relevante sobre la coexistencia entre especies y su relación con gradientes ambientales (Guisan et al., 1999; Sánchez-González y López-Mata, 2003). Además, la significancia estadística del análisis (p=0.001), determinada a través de pruebas de permutación (1000 repeticiones), evidencia una aproximación robusta sobre la relación especies-ambiente (McCune, 1997; Sánchez-González y López-Mata, 2003) observadas en este estudio.
Relaciones ecológicas de especies de pino en México
Con base en un extenso análisis bibliográfico, se determinó que en México existe poca evidencia respecto a la identificación de grupos funcionales del género Pinus. Aunque existen estudios a nivel de sitio que reportan la coexistencia de especies del género, debido a las diferencias de escala y a los propósitos de tales estudios, consideramos no pertinente contrastar nuestros resultados con tales evidencias. Si consideramos estudios regionales, como la descripción de coníferas de Farjon (2017), puede deducirse que, a lo largo de su distribución, P. pseudostrobus se asocia con al menos 16 especies de pino. Así mismo, con base en su rango altitudinal, Sandoval-García et al. (2020) afirman que esta especie se asocia con al menos 17 y 12 especies en la FVT y la SMS, respectivamente. Tales evidencias son razonables, ya que P. pseudostrobus es la tercera especie mexicana de pino con el rango altitudinal (850-3250 m) y geográfico más amplio (Gernandt y Pérez-De La Rosa, 2014; Farjon, 2017), lo que le permite tolerar diversos ambientes, coexistir y asociarse con diversas especies de pino. Desafortunadamente, tales referencias no ofrecen detalles sobre las especies con mayor o menor afinidad ambiental a P. pseudostrobus. Uno de los principales aportes de esta investigación es la identificación de especies de pino con afinidades ambientales a nivel de sistema montañoso en México.
La identificación de grupos funcionales, como los abordados en este estudio, contribuyen a la comprensión de la afinidad ambiental en la distribución de las especies y pueden servir de respaldo en el diseño de estrategias de conservación y restauración basadas en especies con ecologías afines. Dentro del sector forestal mexicano, el agrupamiento de especies puede contribuir al diseño de esquemas de gestión basados en grupos de especies (Lujan-Soto et al., 2015). En este sentido, la agrupación de especies basada en rasgos morfológicos en bosques de Durango, México, realizada por Lujan-Soto et al. (2015) es coincidente con P. durangensis, P. engelmannii, P. arizonica, P. leiophylla, P. teocote y P. strobiformis como un grupo funcional en la SMOc (Fig. 2). La referencia anterior atendió aspectos productivos ya que, en conjunto, las especies mencionadas representan el mayor volumen de madera cosechada en algunas zonas de la SMOc (Moctezuma-López y Flores, 2020). Por el contrario, no se han hecho intentos por identificar grupos de especies de pino que puedan ser considerados con fines de restauración en ambientes degradados o poco productivos, como lo pueden ser las agrupadas por su afinidad a suelos con altos niveles de pH en la SMOc (Fig. 2) y a suelos con densidad aparente alta en la SMOr (Fig. 3).
La agrupación de especies de pino en gradientes ambientales específicos en cada SM tiene implicaciones geográficas y filogenéticas. Por un lado, al tener patrones geográficos coincidentes es de esperar que las especies se relacionen de forma similar con respecto a los gradientes ambientales (Gernandt y Pérez-De La Rosa, 2014). Por otro lado, al ser grupos de especies filogenéticamente relacionadas, no es sorpresivo que algunas de ellas presenten un alto nivel de conservadurismo de nicho, lo que de igual forma justifica una ecología relativamente similar. Por ejemplo, Ortiz-Medrano et al. (2016) afirmaron que P. cembroides, P. johannis y P. discolor presentan un fuerte conservadurismo de nicho, que deriva en preferencias ecológicas similares, lo cual puede justificar la cercanía evidente de estas especies en los gráficos de ordenación generados en este estudio (Fig. 2, 3). En otro estudio, Dvorak et al. (2009) explicaron que la historia evolutiva y geográfica de P. oocarpa ha derivado en una estrecha relación ecológica con P. tecunumanii, P. maximinoi y P. patula, condición que se reafirma en este estudio en la SMOr (Fig. 3), donde P. oocarpa y P. patula alcanzan su mayor latitud (Farjon y Filer, 2013) y en la SMS (Fig. 5) y la SMCh (Fig. 6), donde el resto de las especies tienen mayor relación con P. oocarpa.
Merece la pena mencionar que un factor no biológico que pudo haber forzado la agrupación de especies de pino en los análisis canónicos fue el uso de celdas espaciales de tamaño grande (~10 × 10 km) para simplificar los gradientes ambientales en cada SM y relacionarlos con los datos de ocurrencia. Desafortunadamente, el efecto de la magnitud de la resolución espacial para el estudio de la asociación de especies y su relación con el ambiente es un tema no explorado por la investigación científica.
Relación especies-ambiente
La historia evolutiva del género Pinus estuvo estrechamente ligada a la formación de los principales SM de México (Millar, 1993), siendo la topografía un elemento crucial en la diversificación de los linajes antiguos del género (Jin et al., 2021). A nivel de sitio, la topografía influye sobre el viento, temperatura, humedad, radiación solar, pH y flujo de nutrientes (Guisan y Zimmermann, 2000; Moeslund et al., 2013), que en conjunto condicionan el desarrollo de los árboles. Por ejemplo, la presencia de viento fuerte y altos niveles de radiación solar en altitudes elevadas, pueden limitar el reclutamiento de árboles jóvenes (Astudillo-Sánchez et al., 2019), aumentar la evapotranspiración y causar daños por desecación a árboles adultos (Han et al., 2012). Así mismo, en altitudes altas se propicia una disminución de la humedad, temperatura y oxígeno, lo que limita la descomposición de materia orgánica y condiciona la productividad (Sánchez-González y López-Mata, 2005), siendo esta situación común en hábitats de pinos de alta montaña como P. hartwegii (Sánchez-González y López-Mata, 2003; Astudillo-Sánchez et al., 2019) y P. culminicola (Estrada-Castillón et al., 2015). Lo anterior respalda el hecho de que la altitud, la velocidad del viento, la radiación solar y la presión de vapor de agua hayan sido factores que influyen en la distribución de pinos en los SM estudiados (Cuadro 2).
Los factores edáficos son impulsores importantes de la distribución espacial de las plantas y sus comunidades (Sánchez-González y López-Mata, 2005; Estrada-Castillón et al., 2015). Particularmente, los suelos con textura fina favorecen la retención de humedad (Moeslund et al., 2013), facilitan la descomposición de materia orgánica (Xia et al., 2020), mejoran la distribución y disponibilidad de nutrientes en el suelo y propician que el sistema radicular sea eficiente en los árboles (Thammanu et al., 2021). Lo anterior respalda nuestro hallazgo de que el porcentaje de arcilla en el suelo es un factor que influye en la distribución de los pinos, al resultar seleccionado en cuatro de los seis SM. Sin embargo, el porcentaje bajo de inercia (entre 2.4 y 4.3 %) contribuida por esta variable a la inercia total del ACCAA (Cuadro 2) sugiere que, aunque son importantes, los factores edáficos juegan un rol secundario en la distribución espacial a escala regional de los pinos.
La estacionalidad de la precipitación (b15) tuvo la mayor influencia en la distribución de los pinos de la SBC. Esto coincide con lo reportado por Macías-Rodríguez et al. (2014) quienes afirman que la precipitación y su ritmo estacional condicionan el desarrollo de la vegetación en México, y el crecimiento de especies como P. coulteri, P. quadrifolia y P. jeffreyi en la SBC (Martijena, 2003). Así mismo, Alba-López et al. (2003) encontraron que la presencia de especies de pino en Chiapas es influenciada principalmente por la altitud, lo que concurre con los resultados de este estudio, puesto que esa variable fue la que mayor varianza explicó en el ACC de la SMCh (Cuadro 2). En la FVT, más del 50 % de la varianza del ACC fue explicada por parámetros derivados de temperatura y precipitación y menos del 5% por variables de suelo (Cuadro 2). Este resultado coincide con lo reportado por Estrada-Castillón et al. (2015) y Sánchez-González y López-Mata (2003) en gradientes altitudinales del centro (FVT) y noreste (SMOr) de México. Esto indica que los factores edáficos afectan la distribución a escala local, mientras que el clima lo hace a escalas más generales (Sánchez-González y López-Mata, 2005).
Finalmente, el hecho de que algunas especies se hayan ubicado muy cerca del origen de los gráficos de ordenación del ACCAA, indica que tales especies no mostraron una relación clara con las variables representadas por los primeros dos ejes canónicos. Lo anterior puede deberse a dos razones principales. En primer lugar, es posible que dichas especies sean elementos comunes (especies generalistas) del SM en cuestión. Es decir, son especies que se encuentran ampliamente distribuidas en el espacio ambiental y no muestran afinidad particular hacia ambientes específicos. Por ejemplo, P. leiophylla en la SMOc (Fig. 2) y FVT (Fig. 4), y P. montezumae y P. pseudostrobus en la SMOr (Fig. 3). En segundo lugar, el uso del rango geográfico y ambiental incompleto de las especies en los análisis canónicos. Por ejemplo, la SBC representa la distribución marginal de P. contorta, P. jeffreyi y P. lambertiana, mientras que su distribución geográfica natural alcanza el sur de Alaska para la primera, y Oregón (EUA) en las dos restantes (Farjon y Filer, 2013). Es posible que la falta del espectro ambiental completo de estas especies dificulte la observación de una relación clara entre estas y los gradientes ambientales de la SBC.
Conclusiones
Este estudio presenta un análisis general de las asociaciones ecológicas del género Pinus a nivel de sistema montañoso en México y la relación de tales asociaciones con gradientes ambientales. Mediante análisis de correspondenciacanónica se identificaron al menos cuatro grupos de especies de pinos con preferencias ecológicas relativamente similares en cada sistema montañoso de México. Las variables ambientales mayormente incidentes sobre la distribución de los pinos en México son altitud, presión de vapor de agua, radiación solar, velocidad del viento y el porcentaje de arcilla en el suelo, aunque la magnitud en la importancia de tales factores varía a nivel regional. La identificación de especies con preferencias ecológicas similares es importante para gestionar el mantenimiento de la diversidad ecológica, identificar grupos de especies con mayor o menor resiliencia ante el cambio climático y diseñar esquemas de gestión forestal multipropósito.

