












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
Las abejas juegan un papel esencial en el planeta, no solo para la producción agrícola, sino también por su papel en la reproducción de plantas silvestres (Klein et al., 2007). Paradójicamente, el uso de plaguicidas en la agricultura extensiva ha provocado gran pérdida de polinizadores, lo que a su vez disminuye la productividad de los cultivos y pone en riesgo la seguridad alimentaria (Botías & Sánchez-Bayo, 2018). Se estima que el valor económico mundial de los servicios de polinización es de 235 a 577 mil millones de dólares al año (Cairns et al., 2017). Para incrementar el servicio de polinización en sistemas agrícolas abiertos se han implementado colonias domesticadas de la abeja de la miel, Apis mellifera (Peña & Carabalí, 2018), a pesar de que se ha demostrado que las abejas nativas también son visitantes de flores y polinizadores importantes en comunidades de plantas silvestres en áreas de alta biodiversidad (Garibaldi et al., 2013; Stanley et al., 2020).
Para el caso de los sistemas cerrados, como invernaderos, ha predominado el uso de abejorros de las especies Bombus terrestris y B. impatiens, los cuales se producen de manera masiva (Cerna-Chávez et al., 2015). Para el caso de México, estas especies de abejorros son exóticas, por lo cual su implementación en la agroindustria podría presentar riesgos ecológicos, como la importación de patógenos (Otterstatter & Thomson, 2008) y el desplazamiento de especies locales, como ha ocurrido en otros países (Inoue et al., 2008). Debido a esto es de gran relevancia la implementación de polinizadores nativos en la agricultura.
Los meliponinos, o abejas sin aguijón (Apinae: Meliponini), son una tribu de abejas eusociales que se distribuyen en las regiones tropicales y subtropicales del planeta (Hartfelder, 2008). Este grupo de abejas es particularmente abundante en el continente americano, con 60 géneros (Michener, 2022) y más de 600 especies (Roubik, 2023). Algunas especies de meliponinos poseen características favorables para su implementación como polinizadores en la agricultura (Quezada-Euán et al., 2023). La especie Nannotrigona perilampoides (Cresson), por ejemplo, es una especie de amplia distribución y que ha probado ser un excelente polinizador de chile habanero y tomate (Palma et al., 2008a, b; Quezada-Euán, 2009). Sin embargo, así como con otros meliponinos, su reproducción es lenta y su implementación a gran escala requiere el desarrollo de técnicas para su propagación masiva (Menezes, 2015).
La producción masiva de meliponinos ha sido un tema de interés y desarrollo en los últimos años (Quezada-Euán et al., 2023). El objetivo es solucionar el problema que representa la lentitud con la que se reproducen (Menezes, 2015). En condiciones naturales, una colmena puede tardar años en producir nuevos enjambres. Durante la enjambrazón, proceso en el cual las abejas forman nuevas colonias, las obreras se encargan de provisionar de recursos el nuevo nicho (Roubik, 2006; Oliveira et al., 2013). Una vez acondicionado el lugar, la abeja reina virgen realiza el vuelo nupcial, junto con machos de otras colonias cercanas y una vez apareada, un grupo de abejas obreras llevan a la abeja reina a su nuevo nido donde, al cabo de unos días, comenzará a ovipositar (Roubik, 2006; Oliveira et al., 2013). En la mayoría de los géneros de meliponinos (a excepción de Melipona), las reinas se desarrollan en celdas reales, que son de mayor tamaño, a diferencia de las celdas de zánganos y obreras. Todas las celdas de la colonia se abastecen con alimento larval (mezcla de miel, polen y sustancias glandulares), pero cuando las larvas reciben una mayor cantidad de alimento, se desarrollan como reinas. Este fenómeno se denomina “determinación trófica de castas” (Quezada-Euán, 2018).
Las técnicas desarrolladas hasta ahora plantean la producción masiva de reinas mediante la cría in vitro de larvas, bajo condiciones controladas de humedad, temperatura y cantidad de alimento larval. Sin embargo, los equipos de laboratorio utilizados suelen ser costosos y de difícil aplicación en condiciones rurales. En estudios realizados por Menezes et al. (2013) con Scaptotrigona depilis (Moure) se determinaron los parámetros de humedad y temperatura adecuados a lo largo del desarrollo larval; humedad del 100% durante los primeros 6 días y del 75% durante el resto del desarrollo, a una temperatura constante de 28°C, permitió mejorar la tasa de supervivencia, en comparación con estudios anteriores. Las condiciones de esterilidad en el desarrollo de las larvas también es un factor importante (Álvarez-Hidalgo et al., 2020; Van Looveren et al., 2023), el cual suele controlarse mediante el uso de autoclave y campana de flujo laminar. Estos equipos suelen ser costosos e inaccesibles para meliponicultores, por lo que es importante que se evalúen y desarrollen nuevas alternativas de bajo costo y accesibles para su aplicación en zonas rurales.
Si bien las técnicas de producción in vitro de reinas han dado buenos resultados, la fecundación y formación de nuevas colonias actualmente es un reto. Para solventar esta problemática Menezes (2015) ha planteado utilizar núcleos de fecundación: colonias sin reina, pero con suficiente número de obreras, en las cuales se introducen las reinas vírgenes para que sean aceptadas y fecundadas de manera natural en su vuelo nupcial. Posteriormente las reinas fecundadas son trasladadas a mini-colmenas: colmenas pequeñas las cuales permanecen confinadas y alimentadas artificialmente hasta que tengan suficientes obreras y los recursos necesarios para ser autónomas, proceso que dura alrededor de 4 meses (Menezes, 2015). Por ejemplo, se demostró que las obreras jóvenes (las cuales no tienen pigmentación) de la abeja sin aguijón Plebeia droryana (Friese) aceptan con mayor frecuencia reinas producidas in vitro, que obreras viejas (pigmentadas) (Dos-Santos et al., 2016).
En el presente estudio se realizaron varias modificaciones a la metodología propuesta por Menezes (2015) para la cría de reinas in vitro y la formación de nuevas colonias de N. perilampoides, con el objetivo de desarrollar un método para la producción masiva de colmenas de esta especie. El estudio se enfocó en evaluar (1) el efecto de la cantidad del alimento larval en el desarrollo, emergencia y tamaño de reinas y (2) el establecimiento de reinas fisogástricas (reinas fecundadas que presentan abdomen ensanchado) en núcleos de fecundación bajo diferentes condiciones (obreras de diferente edad) y (3) el efecto de la esterilidad del material y el proceso de traslarve en el desarrollo y la emergencia de reinas.
Materiales y métodos
Sitio de estudio. Los experimentos se realizaron en las instalaciones de El Colegio de la Frontera Sur (ECOSUR), Unidad Tapachula, Chiapas en los meses de febrero a abril y de noviembre a diciembre del 2022, y en un laboratorio de campo en Cholul, Mérida, Yucatán, en los meses de mayo a agosto del 2022. Para la crianza de reinas se realizaron 9 corridas experimentales, de las cuales 5 se llevaron a cabo en Tapachula y 4 en Mérida. De los 26 núcleos de fecundación, 10 se hicieron en Tapachula y 16 en Mérida.
Material biológico. Para el caso de los experimentos realizados en Tapachula, las colmenas de N. perilampoides (N=9) se obtuvieron de diversos meliponarios (para evitar parentesco) provenientes de zonas cercanas a Tapachula. Los experimentos realizados en Yucatán se llevaron a cabo con colonias (N=17) de meliponarios cercanos a Mérida y Cholul, también sin parentesco. En ambos sitios se verificó que hubiera presencia de colonias naturales de N. perilampoides que pudieran servir como fuente de machos para las reinas que se producirían in vitro y propiciar una fecundación natural.
Alimento larval. El alimento larval se obtuvo de aproximadamente 150 celdas que no tuvieran más de una semana de construcción. Se usó una jeringa estéril nueva de 5 ml para recolectar al menos 3 ml de alimento larval, el cual se depositó en un vial. Este procedimiento duró casi una hora, por lo que, para prevenir crecimiento microbiano, el vial se selló y se esterilizó mediante exposición a luz UV durante 15 min. Las larvas o huevos de las celdas de las que se obtuvo el alimento larval se descartaron.
El alimento larval se distribuyó en placas ELISA de 96 pozos y para prevenir contaminación, se alternó poner alimento en un pozo, dejando vacíos los de alrededor, dando una forma de tablero de ajedrez. Las cantidades evaluadas de alimento fueron 38, 40, 42, 44, 46 y 48 µl, que tuvieron entre tres a siete repeticiones por placa, dependiendo de la disponibilidad del alimento. La placa se colocó sin tapa, dentro de una caja Petri de 15 cm de diámetro y 2 cm de altura, con 30 ml de agua destilada estéril para mantener la humedad relativa necesaria. La caja Petri se envolvió con una película plástica adherente (Ekco México) y se expuso a luz UV por 15 min. Las cantidades de alimento larval evaluadas se eligieron con base en los resultados obtenidos de una prueba piloto realizada en ECOSUR, Unidad Tapachula, en donde se observó que las reinas se desarrollaron a partir de 38 µl. Cada placa ELISA corresponde a una corrida experimental (N=9).
Traslarve. Los traslarves se realizaron con una aguja de traslarve estéril (esterilizada con olla de presión o con autoclave, según el tratamiento). Sólo se eligieron larvas recién emergidas (de celdas de 5 a 6 días de haberse construido) y que aún no estuvieran recostadas sobre el alimento, después se colocaron cuidadosamente en cada uno de los pozos de la placa encima del alimento larval, a fin de evitar que sus espiráculos se taparan con el alimento larval. Este proceso se realizó en un espacio limpio (cabina para PCR o campana de flujo laminar, según el tratamiento) para no contaminar el alimento larval dentro de los pozos. Las condiciones de esterilidad evaluadas se agruparon en tres tratamientos: a) uso de olla de presión para la esterilización del material y cabina para PCR para los traslarves, b) autoclave para la esterilización del material y cabina para PCR para los traslarves y c) uso de autoclave para la esterilización del material y campana de flujo laminar para los traslarves.
Incubación. Las placas con el alimento y las larvas se colocaron en una incubadora a 29-30 °C. La humedad relativa se controló a 99 % durante los primeros 6 días, mediante 30 ml de agua administrados en la caja Petri y a 75 % el tiempo restante del desarrollo larval mediante la adición de 9 g de NaCl (Menezes et al., 2013).
Emergencia y medición de reinas. Con la finalidad de evitar el ahogamiento de las reinas emergentes, se retiró toda el agua de la caja Petri a partir del día 31 después del traslarve. Las reinas producidas se destinaron a la formación de núcleos o para la medición de su distancia intertegular (distancia lineal entre las tégulas de la abeja), que se utiliza como indicador del tamaño para comparar entre tratamientos (Añino et al., 2024). Para el primer caso se seleccionaron al azar reinas (N=24) de los tratamientos de dieta y se colectaron reinas vírgenes naturales (N=2) para destinarlas a los núcleos de fecundación. En el segundo caso se seleccionaron reinas de todos los tratamientos de dietas (N=46) y se colectaron otras de origen natural (N=15). Se sacrificaron por enfriamiento a -20 ºC y se colocaron sobre un portaobjetos con una planilla milimétrica bajo un microscopio estereoscópico (Zeiss, Modelo Stereo Discovery. V20, Alemania). Después se obtuvieron las fotografías mediante una cámara (Nikon, Modelo D810, Japón) y con la finalidad de medir la distancia intertegular se usó el programa informático ImageJ versión 1.53t (Schneider et al., 2012).
Introducción a núcleos. Las reinas destinadas a los núcleos se marcaron con plumones POSCA (PC-3M, MITSUBISHI PENCIL CO, Tokyo, Japón) y se introdujeron conforme emergieron, en núcleos huérfanos formados con 48 h de anticipación. Para ello se llevó a cabo un proceso de familiarización entre las reinas y sus respectivos núcleos, que consistió en colocar a las reinas dentro de viales agujerados y cerrados, los que se introdujeron en las colmenas. Los viales se abrieron 24 horas después de haberlos introducido a los núcleos. Los núcleos utilizados consistieron en cajas de madera de cedro de 13 cm x 13 cm x 9 cm de altura, con un grosor de madera de 2 cm y un volumen interior de 405 cm3, que corresponden a las dimensiones de gavetas de colmenas inteligentes Ailton Fontana (AF). Las colmenas inteligentes AF se forman con gavetas de madera y se clasifican según las estructuras de la colmena que albergan (nido, sobre nido y melario) (Cortes-Martínez & Olarte-Blandon, 2019). Debajo de la tapa de madera de los núcleos se colocó un acetato transparente para facilitar la observación sin afectar a las abejas. Se les proporcionó un pote de miel y uno de polen de colonias de la misma especie. Estos núcleos se ubicaron a la intemperie, bajo sombra, para permitir la salida y entrada natural de las obreras. Los tratamientos de los núcleos fueron los siguientes:
Núcleos formados (N=9) con 300 obreras viejas (pigmentadas) y un panal de cría próxima a emerger, con al menos 150 celdas.
Núcleos formados (N=11) con 300 obreras jóvenes (no pigmentadas) y un panal de cría próxima a emerger, con al menos 150 celdas.
Melario de colmena inteligente AF (N=3) con abundantes obreras de todas las edades y abundantes reservas de polen.
Melario de colmena inteligente AF (N=3) con abundantes obreras de todas las edades y abundantes reservas de miel.
Los núcleos de fecundación se monitorearon constantemente durante las primeras dos semanas, para determinar la aceptación o rechazo de las reinas. Posteriormente se realizaron revisiones cada dos días, hasta que los núcleos dejaran de tener actividad o que se observara a la reina marcada, con postura en celdas construidas. Las observaciones se llevaron a cabo a través de los acetatos transparentes colocados en la parte superior de los núcleos, con la finalidad de no afectar a las abejas durante las observaciones. Los criterios considerados como aceptación fueron: trofalaxis o transferencia de alimento entre la reina y las obreras, y la permanencia de la reina en el núcleo por más de una semana desde su introducción. Por otra parte, los criterios de rechazo fueron: mutilación o muerte de la reina. Se consideró como caso de éxito a las reinas que fueron capaces de aparearse e iniciar postura, para la formación de una nueva colonia.
Análisis estadísticos
Efecto de la cantidad de alimento larval en el desarrollo y tamaño de reinas. Para evaluar la eficacia de la técnica de cría de reinas in vitro se registró para cada tratamiento (38, 40, 42, 44, 46 y 48 µl de alimento larval): (a) el número de larvas puestas (traslarve), (b) el número de reinas obtenidas y (c) el porcentaje de eficiencia por tratamiento de las nueve corridas experimentales en cajas Petri.
Los datos de emergencia y tamaño de reinas se analizaron mediante un enfoque de GLM revisando los supuestos de normalidad y homocedasticidad. Para la emergencia de las reinas se utilizó el promedio por tratamiento, y para el análisis con los tamaños de reinas; se utilizó como indicador las distancias intertegulares. Todos los análisis estadísticos se realizaron en R versión 4.2.1 (R Development Core Team, 2020).
Para determinar la cantidad de alimento óptimo para el desarrollo in vitro de reinas, se calculó un factor de optimización (FDO). El factor es el producto del tamaño promedio de las reinas de cada tratamiento, por el porcentaje de emergencia correspondiente.
Efecto de las condiciones de esterilidad en el material y el proceso de traslarve. Los porcentajes de emergencia de las reinas se agruparon, según los tres tratamientos de esterilidad aplicados para el material utilizado durante los traslarves, a los que se les hicieron pruebas de normalidad y homocedasticidad, y se analizaron con un GLM con distribución Gaussiana.
Efecto de la configuración de núcleos de fecundación en la aceptación de reinas producidas in vitro . Se contó el número de núcleos en los que se observó rechazo de la reina (no se observó reina, o reina mutilada y muerta) o aceptación (trofalaxis con obreras o permanencia de la reina por más de una semana). Este recuento se sujetó a un análisis GLM asumiendo una distribución binomial. También se determinó el porcentaje de las reinas que se aparearon y formaron nuevas colonias.
Resultados
Efecto de la cantidad de alimento larval en el desarrollo, emergencia y tamaño de reinas. La cantidad de alimento suministrado a N. perilampoides tiene un efecto negativo significativo en el número de reinas producidas in vitro (Cuadro 1, F5,35 = 5.846, p < 0.001). Sin embargo, se encontró que la cantidad de alimento larval se relaciona positivamente con el tamaño de las reinas (Fig. 1, F5,60 = 2.440, p = 0.036). Se observó que, las reinas naturales tienen mayor variación en sus tamaños, mientras que los tratamientos de 38, 40, 42 y 46 µl de alimento larval, pueden formar reinas de tamaño semejante (alfa = 0.1). Por otro lado, las reinas desarrolladas con los tratamientos de 44 y 48 µl resultaron en reinas de mayor tamaño, que las reinas naturales (Cuadro 1, Fig. 1).
Cuadro 1 Distancia intertegular de las reinas de N. perilampoides criadas in vitro, proporción de emergencia y factor de optimización para cada tratamiento. Letras diferentes indican diferencias significativas.
| Dieta: alimento larval (µl) | Distancia intertegular (media ± DE) (n) mm | Emergencia de reinas (media) (n) | Factor de optimización (FDO) |
|---|---|---|---|
| 38 | 1.326 ± 0.035 ab, (10) | 0.718, (18) | 0.953 |
| 40 | 1.361 ± 0.027 ab, (9) | 0.762, (16) | 1.037 |
| 42 | 1.336 ± 0.048 ab, (8) | 0.762, (17) | 1.018 |
| 44 | 1.390 ± 0.031 b, (6) | 0.728, (18) | 1.012 |
| 46 | 1.339 ± 0.042 ab, (5) | 0.499, (12) | 0.668 |
| 48 | 1.376 ± 0.043 b, (8) | 0.573, (13) | 0.789 |
| Natural | 1.283 ± 0.126 a, (15) | N/A | N/A |
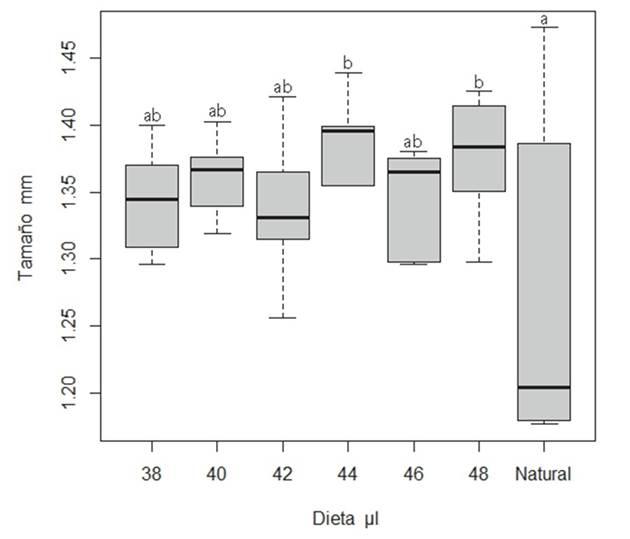
Figura 1 Tamaño promedio de las reinas de N. perilampoides (utilizando como indicador la distancia intergular) criadas in vitro, con distintas cantidades de alimento larval. Los grupos que difieren significativamente entre sí a un nivel de significancia del 90% están etiquetados con diferentes letras (a, b).
Efecto de la configuración de núcleos de fecundación en la aceptación de reinas producidas in vitro . No se observó una diferencia significativa en la aceptación de las reinas, en función a la estructura de las obreras en los núcleos de fecundación. En todos los núcleos de los cuatro tratamientos las obreras aceptaron a las reinas.
Efecto de las condiciones de esterilidad en el material y el proceso de traslarve. Se observó que las medidas para mantener condiciones de esterilidad de los materiales utilizados, al igual que en el proceso de traslarve, tienen un efecto significativo en la emergencia de las reinas (F2,35 = 16.458, p < 0.05). Se logró obtener una emergencia de reinas de hasta el 100 % utilizando las cantidades de alimento larval de 38, 40, 42, 44, 46 y 48 µl en condiciones de esterilidad estrictas, mediante el uso de autoclave y campana de flujo laminar (Cuadro 2, Fig. 2), cuyo promedio de emergencia fue del 90.5 ± 0.06 %. Para el caso en el que se usó únicamente autoclave para esterilizar las herramientas de trabajo y una cabina de PCR durante la extracción del alimento larval y los traslarves, se obtuvo 73.0± 10.6% en la emergencia de reinas. Finalmente, en el tercer caso sólo se usó una olla de presión para la esterilización del material y una cabina de PCR durante la extracción del alimento larval y los traslarves, consiguiéndose la menor emergencia en este estudio: 52.0 ± 16.8%.
Cuadro 2 Porcentaje de emergencia de reinas de N. perilampoides para cada corrida, lugar y tecnología utilizada para el control de la esterilidad durante el traslarve.
| Núm. Corrida | Sitio | Tecnología | Número de Larvas | Traslarves por tratamiento (µL de alimento larval) | Proporción de emergencia | Reinas producidas | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 38 | 40 | 42 | 44 | 46 | 48 | ||||||
| 1 | Tapachula | Autoclave | 35 | 6 | 5 | 5 | 6 | 6 | 7 | 0.657 | 23 |
| 2 | Tapachula | Autoclave | 17 | 3 | 3 | 3 | 3 | 3 | 2 | 0.882 | 15 |
| 3 | Mérida | Olla de presión | 23 | 4 | 4 | 4 | 4 | 4 | 3 | 0.434 | 10 |
| 4 | Mérida | Olla de presión | 23 | 4 | 4 | 4 | 4 | 4 | 3 | 0.304 | 7 |
| 5 | Mérida | Olla de presión | 25 | 5 | 4 | 4 | 4 | 4 | 4 | 0.560 | 14 |
| 6 | Mérida | Olla de presión | 25 | 3 | 3 | 3 | 3 | 3 | 3 | 0.760 | 19 |
| 7 | Tapachula | Autoclave + CFL | 18 | 3 | 3 | 3 | 3 | 3 | 3 | 0.888 | 16 |
| 8 | Tapachula | Autoclave + CFL | 17 | 3 | 3 | 3 | 3 | 3 | 6 | 1.000 | 17 |
| 9 | Tapachula | Autoclave + CFL | 18 | 3 | 3 | 3 | 3 | 3 | 3 | 0.833 | 15 |
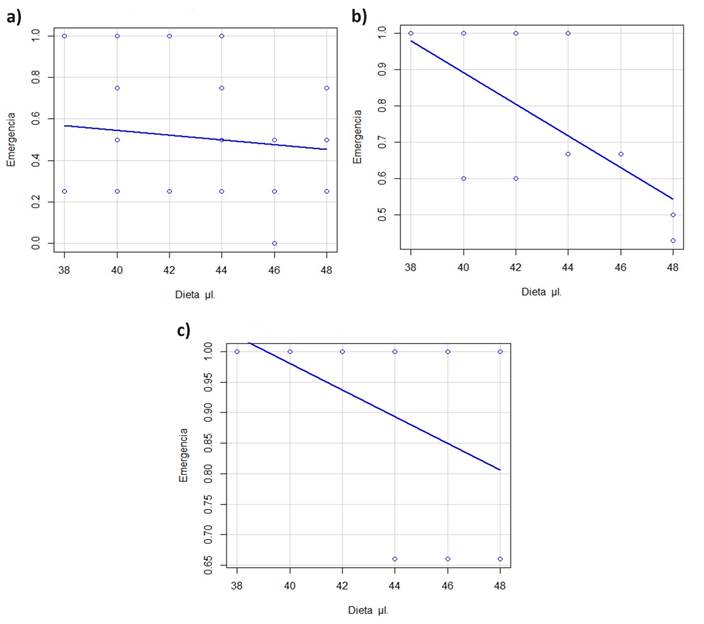
Figura 2 Probabilidad de emergencia de reinas de N. perilampoides obtenidas con distintas cantidades de alimento larval, utilizando tres condiciones de esterilidad durante el traslarve: a) con olla de presión, b) con autoclave y c) con autoclave y campana de flujo laminar.
Formación de nuevas colonias a partir de reinas criadas in vitro . Independientemente de los tratamientos de alimentación previos, de los 10 núcleos formados en Tapachula, ninguna reina criada in vitro fue capaz de fecundarse e iniciar postura. Para el caso de los núcleos formados en Mérida (N=16), tres resultaron exitosos, con reinas criadas in vitro que fueron capaces de aparearse e iniciar postura, que posteriormente se desarrollaron en colonias completas. Las reinas exitosas pertenecieron a los tratamientos de 38, 42 y 44 µl de alimento larval (obtenidas bajo el proceso de esterilización del material con olla exprés) y posteriormente fueron introducidas en melarios de colmenas AF, con una gran cantidad de potes de polen para los correspondientes a 38 y 44 µl y gran cantidad de potes de miel para el correspondiente a 42 µl.
Reinas in vitro óptimas. Se compararon los FDO y se obtuvo que las dietas de 40, 42 y 44 µl fueron las óptimas en función de la emergencia y tamaño de reinas, de las cuales las de 40 y 42 µl tuvieron tamaños similares a las reinas naturales (Cuadro 1).
Discusión y conclusión
Los resultados muestran que es posible la formación de colmenas completamente funcionales de N. perilampoides mediante la cría in vitro de reinas. También se observó que es posible obtener un elevado porcentaje de emergencia de reinas in vitro, utilizando entre 38 y 48 µl de alimento larval con estrictas condiciones de esterilidad.
La técnica desarrollada en este estudio logró hasta un 100% de emergencia de reinas criadas in vitro para N. perilampoides, en contraste con crías de reinas de Plebeia droryana, Heterotrigona itama, Frieseomelitta varia y Scaptotrigona depilis, en las cuales se han logrado emergencias de hasta el 45, 78, y 97.9 %, respectivamente (Baptistella et al., 2012; Menezes et al., 2013; Dos-Santos et al., 2016; Razali et al., 2021; Fahimee et al., 2022). Aspectos en común en estas investigaciones, son el control de una humedad elevada (< 95%) durante los días de alimentación de la larva y posteriormente una reducción por debajo del 85% (Menezes et al., 2013; Dos-Santos et al., 2016;). Otro aspecto relevante es el estadio del huevo o larva al momento de ser transferido a la placa ELISA. En investigaciones con F. varia se observó que la transferencia de huevos ocasionaba una mayor mortalidad, que la de larvas de primer estadio (Silva et al., 2022). Otra variación de la metodología utilizada por Baptistella et al. (2012) para la cría in vitro de reinas, fue transferir larvas grandes de celdas de obreras aún en proceso de alimentación y administrarles la cantidad adicional para que se determinaran como reinas. Con esa metodología se reportó una emergencia del 45.75%, no obstante, de éstas el 46% se determinó como obrera (Baptistella et al., 2012). En el presente estudio se traslarvaron larvas de N. perilampoides con pocas horas de haber eclosionado, y se tomaron únicamente aquellas que aún no se habían recostado sobre el alimento. Elegir larvas con estas características permitió una alta emergencia y un mejor control en la cantidad de alimento que la larva recibió durante su desarrollo.
Los datos revelan que la cantidad de alimento larval tiene un efecto positivo sobre el tamaño de las reinas, lo cual coincide con los resultados observados con P. droyana (Dos-Santos et al., 2016) y con N. perilampoides (Quezada-Euán et al., 2011). Fue interesante observar que las reinas naturales tuvieron tamaños muy diversos, los cuales coincidían con los obtenidos de tratamientos desde 38 hasta los 46 µl de alimento larval, pero no se observaron reinas naturales de tamaño correspondiente al tratamiento de 48 µl. Si bien se ha demostrado que reinas grandes poseen una mayor fecundidad que reinas de menor tamaño (Ribeiro et al., 2006), en este estudio sólo se fecundaron reinas correspondientes de dietas con 38 a 44 µl, por lo que no se pudo confirmar que reinas más grandes (obtenidas con dietas de 46 y 48 µl), fueran capaces de fecundarse naturalmente e iniciar postura de huevos. Asimismo, las celdas naturales de N. perilampoides que se evaluaron contenían 40 µl de alimento larval, por lo que 44 µl se ajusta a la recomendación de algunos autores, para administrar un 10% adicional de alimento y obtener reinas de tamaños similares a las naturales (Menezes et al., 2013; Dos-Santos et al., 2016). No obstante, conforme se aumentó la cantidad de alimento larval, disminuyó la probabilidad de emergencia de las reinas, posiblemente porque también aumentó la probabilidad de que éste se contaminara, provocando que la larva se enferme y muera. Esto se ha observado tanto en H. itama (Cockerell), como en F. varia (Lepeletier) (Baptistella et al., 2012; Razali et al., 2021). Por ello resulta de suma importancia la esterilidad en el manejo del alimento larval, especialmente cuando se use en cantidades mayores para la cría de reinas de mayor tamaño.
La esterilidad de los materiales utilizados y la asepsia durante el proceso de traslarve, son fundamentales para mantener una emergencia alta, hasta del 100 % en la cría in vitro de reinas. Sin embargo, algunos de los equipos que permiten esta esterilidad, como la autoclave y la campana de flujo laminar suelen ser costosos y de difícil acceso para productores pequeños o en zonas rurales. Pero es posible usar equipos de menor costo, como olla de presión y cámara de vidrio o acrílico con mecheros de gas o alcohol, que aparentemente son menos eficientes, pero pueden ayudar a realizar la cría de reinas con una emergencia de hasta el 76 %.
Las dietas de 40, 42 y 44 µl resultaron ser óptimas para una mayor emergencia (> 72%) y mayor tamaño de las reinas. Asimismo, se demostró que reinas de estos tratamientos fueron capaces de copular e iniciar postura de huevos, formando nuevas colonias. De estos tres tratamientos, los de 40 y 42 µl correspondieron a reinas que pueden ser de tamaños similares a las naturales. Aunque como se observa en la Figura 1, las reinas naturales, a pesar de su variación, tienden a ser más pequeñas.
En cuanto a la configuración de obreras en los núcleos de fecundación, no se observó ningún efecto con respecto a la aceptación de las reinas, por lo que, realizando el proceso de familiarización, el riesgo de rechazo de las reinas es casi nulo. Sin embargo, sí se observa una tendencia al rechazo por parte de las obreras viejas, este resultado coincide con lo observado con P. droryana (Dos-Santos et al., 2016). De las reinas aceptadas en Mérida, sólo el 18.75% (N = 3 de 16 núcleos) fueron capaces de aparearse e iniciar postura para formar nuevas colonias, las cuales correspondieron a dos núcleos de fecundación con abundantes potes de polen y uno con abundantes potes de miel. Esto da pie a cuestionamientos como; que la aceptación de las reinas no es suficiente para que estas puedan aparearse y formar una nueva colonia, o que se requieren de otros factores correspondientes a las características de los núcleos de fecundación, más allá de la presencia y configuración de las obreras. Por otro lado, de las reinas aceptadas en Tapachula, ninguna fue capaz de aparearse e iniciar postura. Posiblemente esto se deba a la baja presencia de zánganos en el sitio y las abundantes lluvias durante esta fase del experimento. En investigaciones realizadas con S. depilis y Tetragonisca angustula (Latreille), se obtuvieron porcentajes de efectividad del 26% y del 13.3 %, respectivamente (Menezes et al., 2013; Prato & Soares, 2013; Martínez-Cifuentes, 2015), muy similares a los resultados obtenidos en esta investigación.
En conclusión, es posible la producción masiva de colmenas de N. perilampoides con la técnica para la cría de sus reinas in vitro desarrollada en este estudio, logrando emergencia de hasta el 100% con cantidades de alimento larval desde 38 hasta 48 µl. La edad de las obreras no fue un factor que afectara la aceptación de las reinas. Las cantidades de alimento larval que demostraron producir reinas capaces de aparearse e iniciar postura fueron de 38 a 44 µl, de las cuales de 40 a 42 µl resultaron ser las óptimas, en función de una mayor probabilidad de emergencia y tamaño de las reinas producidas. También es importante considerar las condiciones de esterilidad en el proceso, el estadio de las larvas transferidas, y cómo estos factores pueden incidir en la emergencia. En cuanto a la formación de nuevas colonias, deben evaluarse otros parámetros con respecto a los núcleos de fecundación, esto con la finalidad de facilitar el apareamiento de las reinas y mejorar el porcentaje de éxito actual en la formación de nuevas colonias de esta especie (18.75%) y de otras abejas sin aguijón.

