Articles
From the bottom up - attributes of small vesper bats’ activity sites in an upland ecosystem in Chiapas, México

-
Publication dates-
December 02, 2024
May , 2023
- Article in PDF
- Article in XML
- Automatic translation
- Send this article by e-mail
- Share this article +
Abstract
We studied bats in a natural protected area and the influence zone in Chiapas, México. Focusing on small vesper bats (Vespertilionidae: Myotinae) in water sinkholes, flooded surfaces, and forested locations (sampling sites), our objectives were to: 1) differentiate the sampling sites based on abiotic and biotic variables, 2) compare the relative activity between the contrasting sampling sites, and 3) determine the relative importance of surrounding land covers amounts on the relative activity. We expected the following: 1) that sampling sites would be differentiable based on an interaction of environmental conditions and nocturnal flying insects’ biomass, 2) to find a significantly higher relative activity associated with water bodies as a reflection of potential drinking and feeding supplies, and 3) that forest cover would have a significant association in a positive direction with the relative activity as a reflection of roosting and commuting opportunities. We obtained weather data with a portable station, collected flying nocturnal insects with a passive trap, and recorded bats with ultrasonic detectors. We also used satellite imagery to calculate land covers amounts around the sampling sites. We performed a multivariate analysis to compare sampling sites, and elaborated correlation models of relative activity against surrounding land covers amounts. The only significant variable for differentiating sampling sites was wind speed; in water sinkholes, with lower mean value compared to flooded surfaces and forested locations, along with a comparatively higher percentage of dipterans in the samples. The mean relative activity of bats was significantly higher in water sinkholes, with maximum values reaching 95 %, which we can relate to environmental conditions and resources available. We found associations in a positive direction with forest cover, secondary forest and water surface, and negative with agricultural land and human development. Finally, we argue that some of the water sinkholes meet the criteria of small natural features supported by the unusual activity of the studied bats, and that targeted conservation actions will complement other strategies implemented in the area.
Keywords::
Forest loss, hierarchical partitioning, landscape level, local level, Myotis spp., small natural features, targeted conservation
Introduction
The accelerated loss of biodiversity because of human activities is a concern worldwide. Therefore, identifying which attributes in agricultural zones with a high rate of forest loss can back away from the decline of animal populations is crucial (Heim et al.2015). This way, multi-level management may contribute to more suitable habitats for wild species.
-
Heim et al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
For aerial insectivorous bats, localized resources (e. g., water, prey) can represent limiting factors for survival and reproductive success (Findley 1993). At the same time, landscape structure (e. g., forest amount and its spatial arrangement, linear elements, among other attributes) may facilitate commuting toward drinking and feeding sites (Fuentes-Montemayoret al.2017; Mendeset al.2017; Martinoet al.2019).
-
Findley 1993Bats. A Community Perspective., 1993
-
Fuentes-Montemayoret
al.2017Species mobility and landscape context determine the importance of local and landscape-level attributesEcological Applications, 2017
-
Mendeset
al.2017Bat richness and activity in heterogeneous landscapes: guild-specific and scale-dependent?Landscape Ecology, 2017
-
Martinoet
al.2019Activity records of the endangered Paraguaná moustached bat, Pteronotus paraguanensis, in the main vegetation types of the Paraguaná Peninsula, VenezuelaActa Chiropterologica, 2019
Fahrig (2003) argues that the number of individuals of any species should be, to some point, a positive function of the quantity of habitat available, such as native forests. The strong forest dependency some species of bats exhibit depends on roosting preferences and flight restrictions in open areas (Lackiet al.2007; Fuentes-Montemayoret al.2013; Parreira Peixotoet al.2018; Novella-Fernandezet al.2022). Forest loss can decrease structural connectivity for many species (Fischer and Lindenmayer 2007). However, this topic needs to be better studied in the Neotropics, specifically considering slow-flying bats in upland ecosystems.
-
Fahrig (2003Effects of habitat fragmentation on biodiversityAnnual Review of Ecology, Evolution, and Systematics, 2003
-
Lackiet
al.2007Bats in Forests: Conservation and Management, 2007
-
Fuentes-Montemayoret al.2013Fragmented forests in agricultural landscapes: The influence of forest character and landscape context on bats and their insect preyAgriculture, Ecosystems & Environment, 2013
-
Parreira Peixotoet al.2018A synthesis of ecological and evolutionary determinants of bat diversity across spatial scalesBMC Ecology, 2018
-
Novella-Fernandezet al.2022The role of forest structure and composition in driving the distribution of bats in Mediterranean regionsScientific Reports, 2022
-
Fischer and Lindenmayer 2007Landscape modification and habitat fragmentation: a synthesisGlobal Ecology and Biogeography, 2007
Flying away from vegetation cover may impose higher energetic costs for slower species (due to the stronger wind), such as vesper bats (Vespertilionidae), characterized by wings with low aspect ratio and loading (Norberg and Rayner 1987; Heim et al.2015). Moreover, their short and high-frequency modulated echolocation pulses are not well suited to open areas because they are more subject to atmospheric attenuation (Pettersson 2002; Jones and Rydell 2003; Frey-Ehrenboldet al.2013; Heim et al.2015).
-
Norberg and Rayner 1987Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocationPhilosophical Transactions of the Royal Society B: Biological Sciences, 1987
-
Heim
et al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
-
Pettersson 2002The properties of sound and bat detectorsBat Echolocation Research: Tools Techniques and Analysis, 2002
-
Jones and
Rydell 2003Attack and defense: Interactions between echolocating bats and their insect preyBat Ecology, 2003
-
Frey-Ehrenboldet
al.2013Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matricesJournal of Applied Ecology, 2013
-
Heim et
al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
We studied small vesper bats (Vespertilionidae: Myotinae) in water sinkholes, flooded surfaces, and forested locations (sampling sites), inside a natural protected area and the influence zone in Chiapas, southeast México. We based the selection of this group of bats on traits such as wing morphology and echolocation system related to the sensitivity to forest loss (Heim et al.2015). They also are essential predators of nocturnal flying insects (e. g., mosquitoes), which are potentially deleterious to humans and become pests, which should be a key consideration in sustainable land management (Gonsalveset al.2013; Heim et al.2015; Puig-Montserratet al.2020).
-
Heim et
al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
-
Gonsalveset
al.2013Mosquito consumption by insectivorous bats: Does size matters?PLos One, 2013
-
Heim et
al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
-
Puig-Montserratet al.2020Bats actively prey on mosquitoes and other deleterious insects in rice paddies: Potential impact on human health and agriculturePest Management Science, 2020
Our objectives were to 1) differentiate the sampling sites based on abiotic and biotic variables, 2) compare the relative activity between the contrasting sampling sites, and 3) determine the relative importance of surrounding land covers amounts on the relative activity. We expected 1) that sampling sites would be differentiable based on an interaction of environmental conditions and nocturnal flying insects’, 2) to find a significantly higher relative activity associated with water bodies as a reflection of potential drinking and feeding supplies, and 3) that forest cover would have a significant association in a positive direction with the relative activity as a reflection of roosting and commuting opportunities.
Materials and methods
Study area and sampling points. The study area is intermediate between lowlands and highlands, in the comiteca plateau (Meseta Comiteca), Chiapas, México (Figure 1). It includes the Lagunas de Montebello National Park (PNLM, by its initials in Spanish), listed in the RAMSAR convention (no. 1325). The main vegetation association inside the PNLM is a mixed forest dominated by pines (Pinusspp.) and oaks (Quercusspp.), with a canopy stratum between 20 to 35 m and an interior species-rich tree stratum below 20 m height (González and Ramírez 2013). The predominant land cover outside the PNLM is agricultural land. There is a dry season (approximately March to June), a rainy season (approximately July to October), and a windy season (approximately November to February).
-
González and Ramírez 2013Comunidades vegetales terrestresLa Biodiversidad en Chiapas: Estudio de Estado, 2013
Thumbnail
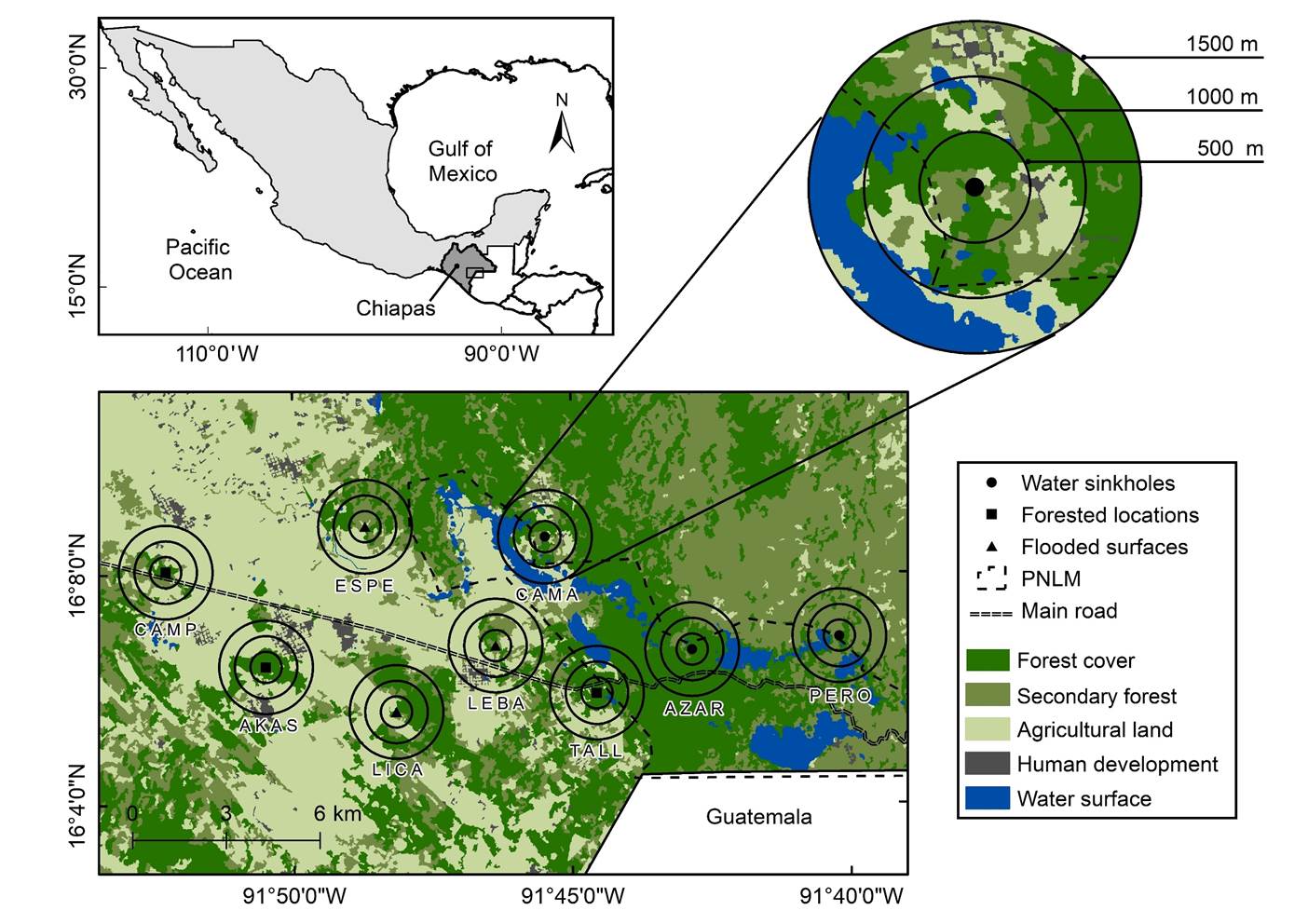
Figure 1
Map of the study area in Chiapas, México. We identify sampling points by different symbols, surrounded by concentric buffers in which we calculated land covers amounts.
Map of the study area in Chiapas, México. We identify sampling points by different symbols, surrounded by concentric buffers in which we calculated land covers amounts.
The sampling points were selected based on physiognomy and pre-sampling, with three repetitions each: water sinkholes, which are open areas with surrounding vegetation and have a permanent water surface several meters below the ground (Figure 2a), flooded surfaces, which are also open areas with surrounding vegetation, and have a fluctuating water surface that is superficial, and represent natural and human-induced elements (Figure 2b), and forested locations, such as clearings and trails inside the forest (Figure 2c). There were no considerable elevation differences in the sampling points, which started from 1,460 to 1,540 m, and accounting for extent restrictions for the spatial analysis, the minimum distance between them started from ~ 3.5 km. The samplings took place from July 2014 to July 2015 to obtain data on environmental conditions, insect collections, and bat recordings simultaneously at each event.
Thumbnail

Figure 2
Sampling points recognizable by physiognomy in the study area in Chiapas, México: a) water sinkholes, b) flooded surfaces, and c) forested locations.
Sampling points recognizable by physiognomy in the study area in Chiapas, México: a) water sinkholes, b) flooded surfaces, and c) forested locations.
Satellite imagery processing and land covers amounts. We used multispectral SPOT 5 orthorectified satellite images (spatial resolution of 10-m pixels) of 2015 (dry season). The latter images were subject to radiometric calibration and were provided by the Laboratorio de Información Geográfica y Estadística (LAIGE, by its initials in Spanish) of El Colegio de la Frontera Sur. We performed composition in false color using bands 3 (near infrared), 2 (red), and 1 (green), with a simple linear contrast. The land covers (classes) defined were (Figure 1): forest cover (mature forest), secondary forest (such as coffee crops and orchards), agricultural land (extensive crops such as maize and grasslands), human development (including urbanization and bare soil), and water surface (lakes, water sinkholes, and flooded surfaces). Following Fuentes-Montemayor et al. (2013), we grouped distinct forest types to avoid confounding information (e. g., adjacent types with undistinguishable limits).
-
Fuentes-Montemayor et al.
(2013Fragmented forests in agricultural landscapes: The influence of forest character and landscape context on bats and their insect preyAgriculture, Ecosystems & Environment, 2013
We performed a supervised classification in Idrisi version 17.0 (Clark Labs, Clark University). Following Lu and Weng (2007) and Eastman 2012, we implemented a segmentation routine that identifies adjacent pixels grouped by spectral similarity, so-called objects. We used a similarity threshold of 30 to obtain homogeneous segments of a size facilitating the selection of training samples. Then, we used the maximum likelihood classifier. Finally, we re-classified the image to improve the precision of the classification and to produce smooth edges between classes by using a distinctive classifier of the segmentation routine. An error matrix (see Verbyla 1995) assessed the overall classification accuracy from 100 field control points spread throughout the study area, obtained during 2014 to 2015, yielding an accuracy of 85 %.
-
Lu and Weng (2007A survey of image classification methods and techniques for improving classification performanceInternational Journal of Remote Sensing, 2007
-
Eastman 2012IDRISI selva tutorial: manual version 17. Clark Labs, 2012
-
Verbyla 1995Satellite remote sensing of natural resources, 1995
We calculated the area (ha) and percentage of each land cover in concentric buffers of 500-m (78.6 ha), 1,000-m (314.2 ha), and 1,500-m (706.8 ha) radii around the sampling points (Supplementary material 1), delimited in ArcGIS version 10.2.1 (ESRI, Inc.). We were looking to encompass the home range of small vesper bats (e. g., Myotis spp.; Owen et al. 2003; Coleman et al. 2014). Also, the nested design allowed us to explore responses by changing spatial scales and the non-overlapping buffers to avoid re-measuring land covers or pseudoreplication (Popescu and Gibbs 2010; Arroyo-Rodríguez et al. 2016).
-
Owen et al. 2003Home-range size and habitat used by the northern Myotis (Myotis septentrionalis)The American Midland Naturalist, 2003
-
Coleman et al. 2014Comparison of radio-telemetric home-range analysis and acoustic detection for little brown bat habitat evaluationNortheastern Naturalist, 2014
-
Popescu and Gibbs 2010Landscape ecology and GIS methodsAmphibian Ecology and Conservation: A Handbook of Techniques, 2010
-
Arroyo-Rodríguez et al. 2016Landscape composition is more important than landscape configuration for phyllostomid bat assemblages in a fragmented biodiversity hotspotBiological Conservation, 2016
Environmental conditions. We used a multi-function weather meter WM-350 WindMate® (WeatherHawk) to obtain monthly data on temperature (°C), relative humidity (%), and wind speed (KMPH). We took measurements three times per night at the beginning, half, and end of the sampling events; this way, we calculated means for comparison between sampling points. We used the mean calculation per night as input for multivariate analyses.
Insects’ biomass. For the collection of nocturnal flying insects, we used a Malaise trap. The latter is a passive trap, which we use to not interfere with recordings by attracting prey. We suspended the trap on trees near the sampling points and sampled around water surfaces near the shore. The trap was installed 1.5 m above ground. All specimens were preserved in 70 % alcohol inside plastic containers (Wickramasingheet al. 2004) and labeled for posterior processing and identification.
-
Wickramasingheet al.
2004Abundance and species richness of nocturnal insects on organic and conventional farms: effects of agricultural intensification on bat foragingConservation Biology, 2004
In the laboratory, we identified the specimens into the orders Diptera (dipterans) and Coleoptera (coleopterans). We pooled Trichoptera and Lepidoptera orders into the superorder Amphiesmenoptera (amphiesmenopterans) because we faced difficulties identifying alcohol-preserved samples. These orders are among the most common food elements for aerial insectivores, particularly this group of bats (see Whitaker 2004; Segura-Trujillo et al. 2018), and may reflect potential prey availability.
-
Whitaker 2004Prey selection in a temperate zone insectivorous bat communityJournal of Mammalogy, 2004
-
Segura-Trujillo et al. 2018Correspondence between ecomorphotype and use of arthropod resources by bats of the genus MyotisJournal of Mammalogy, 2018
We counted the number of individuals of each taxon in each sample (see Queiroz de Oliveira et al. 2015), which were dried later in a stove at 70 °C for 48 h (Bradley et al. 1993), and obtained biomass (g) using an Explorer™ Pro Analytical Balance (EP214C), with a readability of 0.1 mg (Ohaus Corporation). Following Queiroz de Oliveira et al. (2015), we divided it by the number of insects for standardized measurement. Finally, we calculated the mean per night at each sampling point.
-
Queiroz de Oliveira et al.
2015Activity of the insectivorous bat Pteronotus parnellii relative to insect resources and vegetation structureJournal of Mammalogy, 2015
-
Bradley et al. 1993Estimation of insect biomass by length and widthThe American Midland Naturalist, 1993
-
Queiroz de Oliveira et al. (2015)Activity of the insectivorous bat Pteronotus parnellii relative to insect resources and vegetation structureJournal of Mammalogy, 2015
Relative activity of bats. We used an Echo Meter EM3+ Ultrasonic Detector (Wildlife Acoustics, Inc.) to obtain full-spectrum bats recordings. We performed short-term passive recording sessions lasting 4 hrs or else standardized. We configured the EM3+ with 1) sample rate of 256 kHz, 2) WAV audio file format, 3) maximum duration of 15 sec, 4) frequency trigger of 15 kHz, 5) amplitude trigger of 18 dB, 6) trigger window of 1 sec, and 7) gain of 30 dB. We positioned the detector on a 1-m pole with a 45° upward angle, directed toward acoustic space in water sinkholes, flooded surfaces, and forested locations (gaps and trails). Each sampling point was visited twice during three consecutive nights. We stored the recordings in a 32 GB SD card (Kingston© Technology Corporation), and the total recording effort was 432 hrs.
We were interested in small vesper bats flying through the acoustic space of the sampling points. These bats exhibit slow and high-maneuverability flight determined by wing morphology, and high-frequency modulated echolocation pulses of short-range easily attenuated in open areas (Frey-Ehrenbold et al. 2013; Bader et al. 2015), though facing similar restrictions. Some of the species contained in this group of bats can be acoustically cryptic. Identification of those with similar body sizes can be challenging due to the high similarity of the echolocation pulse’s structure, and acoustic parameters overlap (see Jung and Kalko 2011; Williams-Guillén and Perfecto 2011; Estrada-Villegas et al. 2012). Therefore, we pooled recordings to avoid misclassifications.
-
Frey-Ehrenbold
et al. 2013Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matricesJournal of Applied Ecology, 2013
-
Bader
et al. 2015Mobility explains the response of aerial insectivorous bats to anthropogenic habitat change in the NeotropicsBiological Conservation, 2015
-
Jung and Kalko 2011Adaptability and vulnerability of high flying Neotropical aerial insectivorous bats to urbanizationDiversity and Distributions, 2011
-
Williams-Guillén and Perfecto 2011Ensemble composition and activity levels of insectivorous bats in response to management intensification in coffee agroforestry systemsPlos One, 2011
-
Estrada-Villegas et al. 2012Determinants of species evenness in a Neotropical bat ensembleOikos, 2012
The small vesper bats potentially occurring in sympatry in our study area are Rhogeessa tumida, M. nigricans, and M. keaysi pilosatibialis (Barquez and Diaz 2016; Miller et al. 2016; Solari 2019). They belong to the same ecomorphotype and foraging type (Fenton and Bogdanowicz 2002; Segura-Trujillo et al. 2018), classified as aerial/trawling insectivores that hunt in background clutter space such as forest edges and gaps (Schnitzler and Kalko 2001; Frey-Ehrenbold et al. 2013). Accordingly, we selected echolocation pulses starting from a final frequency of 45 kHz. Jung and Kalko (2011) and Estrada-Villegas et al. (2012) indicate the possibility of erroneously classifying Rhogeessa tumida as M. albescens or M. nigricans. We can also find the distribution limits of M. albescens and M. fortidens (see www.iucnredlist.org), but they are more associated with lowlands, so we discard them. Other Myotis species, such as M. velifer and M. californicus, produce lower final frequencies (see Orozco-Lugo et al. 2013; Zamora-Gutiérrez et al. 2016). Finally, Myotis elegans emits frequencies above 60 kHz (O’Farrell and Miller 1999), but it was not detected.
-
Barquez and Diaz
2016Myotis keaysi, 2016
-
Miller et al.
2016Rhogeessa tumida. In: IUCN 2022, 2016
-
Solari 2019Myotis nigricans, 2019
-
Fenton and
Bogdanowicz 2002Relationships between external morphology and foraging behaviour: bats in the genus MyotisCanadian Journal of Zoology, 2002
-
Segura-Trujillo
et al. 2018Correspondence between ecomorphotype and use of arthropod resources by bats of the genus MyotisJournal of Mammalogy, 2018
-
Schnitzler and Kalko 2001Echolocation by Insect-Eating BatsBioscience, 2001
-
Frey-Ehrenbold et al. 2013Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matricesJournal of Applied Ecology, 2013
-
Jung and Kalko (2011Adaptability and vulnerability of high flying Neotropical aerial insectivorous bats to urbanizationDiversity and Distributions, 2011
-
Estrada-Villegas et al. (2012Determinants of species evenness in a Neotropical bat ensembleOikos, 2012
-
Orozco-Lugo et al. 2013Descripción de los pulsos de ecolocalización de once especies de murciélagos insectívoros aéreos de una selva baja caducifolia en Morelos, MéxicoTherya, 2013
-
Zamora-Gutiérrez et al. 2016Acoustic identification of Mexican bats based on taxonomic and ecological constraints on call designMethods in Ecology and Evolution, 2016
-
O’Farrell and Miller 1999Use of vocal signatures for the inventory of free-flying Neotropical batsBiotropica, 1999
For recording processing, in the Bat Analysis Mode, we configured automatic classification using Auto ID for Bats-Bats of the Neotropics (México) with a neutral level of sensitivity, as we were interested in more identifications and not highly accurate ones (see User Guide). We automatically filtered noise files. We processed the recordings in Kaleidoscope Pro v. 5 (Wildlife Acoustics, Inc.). With Kaleidoscope Viewer, we displayed and verified the resulting recordings because automatic identification may produce false positives (see Auto ID for Bats; Rydellet al.2017).
-
Rydellet al.2017Testing the performances of automated identification of bat echolocation calls: A request for prudenceEcological Indicators, 2017
For the calculation of relative activity (a surrogate of bat abundance; Froidevaux et al. 2021), we considered the activity index proposed by Miller (2001). The latter index is based on the total 1-min blocks with evidence of echolocation pulses during constant periods; in our case, for a total of 240 min (4 hrs), otherwise standardized to the total minutes sampled. We considered evidence of at least one recording with a minimum of two consecutive echolocation pulses (MacSwiney et al. 2009; Heim et al. 2015). We expressed the calculations as percentages. For comparisons, we calculated the mean percentage of relative activity at each sampling point for six events.
-
Froidevaux et al. 2021Bat responses to changes in forest composition and prey abundance depend on landscape matrix and stand structureScientific Reports, 2021
-
Miller
(2001A method for determining relative activity of free flying bats using a new activity index for acoustic monitoringActa Chiropterologica, 2001
-
MacSwiney et al. 2009Insectivorous bats activity at cenotes in the Yucatan Peninsula, MéxicoActa Chiropterologica, 2009
-
Heim et al. 2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
Environmental conditions and insects’ biomass analysis. We were interested in differentiating sampling sites based on interacting abiotic and biotic variables. Therefore, we performed a stepwise discriminant analysis. We included the following variables: dipterans biomass, coleopterans biomass, amphiesmenopterans biomass, temperature, relative humidity, and wind speed. We based this analysis on Wilk’s Lambda (λ), the F statistic’s significance, and the independent contributions of variables in the model through Partialλand theF-removestatistic’s significance. We performed these calculations in STATISTICA® version 8.0 (StatSoft, Inc.).
Surrounding land covers amounts analysis. Hierarchical partitioning (hp) is a multivariate exploratory analysis that explains variance in the response variable attributable to univariate correlations with each independent variable (Radford and Bennett 2007). We employed hp to measure the relative importance of surrounding land covers amounts (Chevan and Sutherland 1991; Mac Nally 2000). This analysis has the advantage of addressing potential multicollinearity (Olea et al. 2010).
-
Radford and Bennett
2007The relative importance of landscape properties for forest birds in agricultural environmentsJournal of Applied Ecology, 2007
-
Chevan and
Sutherland 1991Hierarchical partitioningThe American Statistician, 1991
-
Mac Nally 2000Regression and model-building in conservation biology, biogeography and ecology: the distinction between - and reconcilitation of - ‘predictive’ and ‘explanatory’ modelsBiodiversity and Conservation, 2000
-
Olea et al. 2010Estimating and modelling bias of the hierarchichal partitioning public-domain software: implications in environmental management and conservationPlos One, 2010
We performed hp with routine hier.part included in package hier.part version 1.0-6 (Walsh and Mac Nally 2022) ran in R version 3.6.2 (R Core Team 2019). The response variable was the mean percentage of active 1-min blocks, and the independent variables were the percentage of each land cover. The variables were arcsine-transformed [ASIN(SQRT(x/100)], and we ran the models specifying a gaussian distribution, with the goodness of fit based on R 2 . We tested the statistical significance by comparing randomization (1000 permutations) with routine rand.hp included in package hier.part version 1.0-6 (Mac Nally 2002; Walsh and Mac Nally 2022) ran in R version 3.6.2 (R Core Team 2019). We used the generated Z-scores to establish statistical significance based on the upper 95 % (Z ≥ 1.65) confidence limit (Walsh and Mac Nally 2022). The hp does not indicate the direction of paired associations, which we determined by non-parametric correlations.
-
Walsh and Mac Nally 2022Package ‘hier.part’, 2022
-
R Core Team 2019R: A language and environment for statistical computing, 2019
-
Mac Nally 2002Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variablesBiodiversity and Conservation, 2002
-
Walsh and Mac Nally 2022Package ‘hier.part’, 2022
-
R Core Team 2019R: A language and environment for statistical computing, 2019
-
Walsh and Mac Nally 2022Package ‘hier.part’, 2022
Results
Sampling sites. We provide descriptive statistics of environmental conditions, insect collections, and single-variable statistical comparisons between sampling sites (Supplementary materials 2, 3, and 4). The stepwise procedure resulted in a statistically significant model, including only the wind speed, for the differentiation between sampling sites (λ = 0.5259, F (2.24) = 10.8143, P = 0.0004), which exhibited the lowest mean values in water sinkholes in all measurements. We include the calculations for the variables not in the model in Supplementary material 5. The stepwise procedure excluded insect collections. However, we observed a comparatively higher percentage of dipterans in water sinkholes in the samples (Supplementary material 3).
Relative activity of bats. We include the calculations of relative activity in the sampling sites in Supplementary material 6. The difference was statistically significant between sampling sites (K-W = 28.78, P = 5.638E-7; Figure 3), specifically between water sinkholes and flooded surfaces (Dunn post-test, mean rank difference 14.750, P = 0.002), with higher mean value in water sinkholes; between water sinkholes and forested locations (Dunn post-test, mean rank difference 27.250, P = 5.228E-6), with higher mean value in water sinkholes; between flooded surfaces and forested locations (Dunn post-test, mean rank difference 12.500, P = 0.004), with higher mean value in flooded surfaces.
Thumbnail
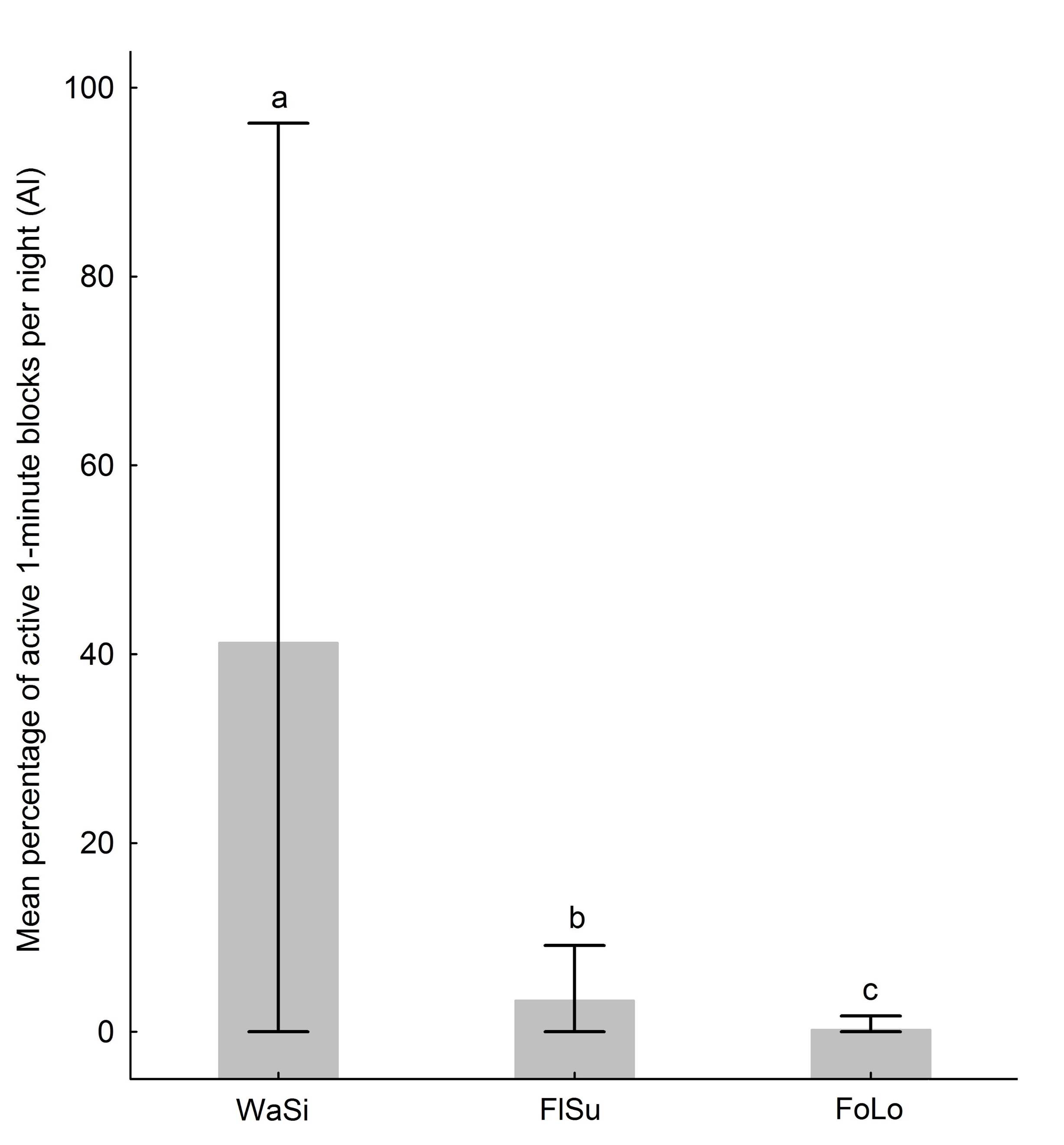
Figure 3
Mean percentage of 1-min blocks per night with acoustic evidence of small vesper bats. The whiskers indicate minimum-maximum values. The different letters indicate statistically significant differences with a 95 % confidence level. WaSi = water sinkholes, FlSu = flooded surfaces, FoLo = forested locations.
Mean percentage of 1-min blocks per night with acoustic evidence of small vesper bats. The whiskers indicate minimum-maximum values. The different letters indicate statistically significant differences with a 95 % confidence level. WaSi = water sinkholes, FlSu = flooded surfaces, FoLo = forested locations.
Surrounding land covers amounts. Invariably, we observed associations in a positive direction with forest cover, secondary forest, and water surface. We observed associations in a negative direction with agricultural land and human development, except for the latter class in the 500-m buffer (9 % of the variance), where a small surface characterized it. In the 500-m buffer, we established that the relative activity is mainly determined by forest cover and secondary forest, explaining 25 % and 26 % of the variance (respectively), and agricultural land explaining 34.7 % of the variance (Figure 4). In the 1,000-m buffer, we established that the relative activity is mainly determined by forest cover and secondary forest, explaining 36.4 % and 20.6 % of the variance (respectively), and agricultural land explaining 23.8 % of the variance (Figure 4). In the 1,500-m buffer, we established that the relative activity is mainly determined by forest cover and water surface, explaining 20.3 % and 42.9 % of the variance (respectively), and agricultural land explaining 28.7 % of the variance (Figure 4). We did not find statistically significant associations based on the upper 95 % confidence limit (Supplementary material 7).
Discussion
In the study area, we could differentiate water sinkholes, flooded surfaces, and forested locations based on the wind speed, with the lowest mean values in water sinkholes in all measurements. Although the insect collections did not stand out in the model, we observed a comparatively higher percentage of dipterans in water sinkholes in the samples. Some dipterans are strongly associated with calming water because their pupae and larvae are susceptible to increasing wind speed (Gillies and Wilkes 1981; Rutledge 2008). The characteristics of water sinkholes, specifically the surface several meters below the ground, can be associated with environmental conditions’ buffering beneficial for this kind of insect.
-
Gillies and Wilkes 1981Field experiments with a wind tunnel on the flight speed of some West African mosquitoes (Diptera: Culicidae)Bulletin of Entomological Research, 1981
-
Rutledge 2008Mosquitoes (Diptera: Culicidae)Encyclopedia of Entomology, 2008
Many Myotis species consume small and soft prey, specifically the aerial ecomorphotypes such asM. nigricansandM. pilosatibialis, which exhibit short wavelength echolocation suitable for these targets (Gonsalveset al.2013), and biomechanical limitations (bite strength) correlated to body size (Segura-Trujillo et al. 2018). Their diet can be composed of several orders, including dipterans (Aguiar and Antonini 2008; Gamboa Alurralde and Díaz 2019; Ingalaet al.2021). However, bats eat a wide range of insects, most representatives seem to have flexible diets (Jones and Rydell 2003), and there is evidence that suggests that the diet responds to local fluctuations, as well as the abundance and type of prey (Salinas-Ramoset al.2015).
-
Gonsalveset
al.2013Mosquito consumption by insectivorous bats: Does size matters?PLos One, 2013
-
Segura-Trujillo et
al. 2018Correspondence between ecomorphotype and use of arthropod resources by bats of the genus MyotisJournal of Mammalogy, 2018
-
Aguiar and Antonini 2008Diet of two sympatric insectivores bats (Chiroptera: Vespertilionidae) in the Cerrado of Central BrazilRevista Brasileira de Zoologia, 2008
-
Gamboa Alurralde and Díaz 2019Feeding habits of four species of Myotis (Mammalia, Chiroptera) from ArgentinaMammal Research, 2019
-
Ingalaet al.2021Molecular diet analysis of neotropical bats based on fecal DNA metabarcodingEcology and Evolution, 2021
-
Jones and Rydell 2003Attack and defense: Interactions between echolocating bats and their insect preyBat Ecology, 2003
-
Salinas-Ramoset
al.2015Dietary overlap and seasonality in three species of mormoopid bats from a tropical dry forestMolecular Ecology, 2015
Thumbnail
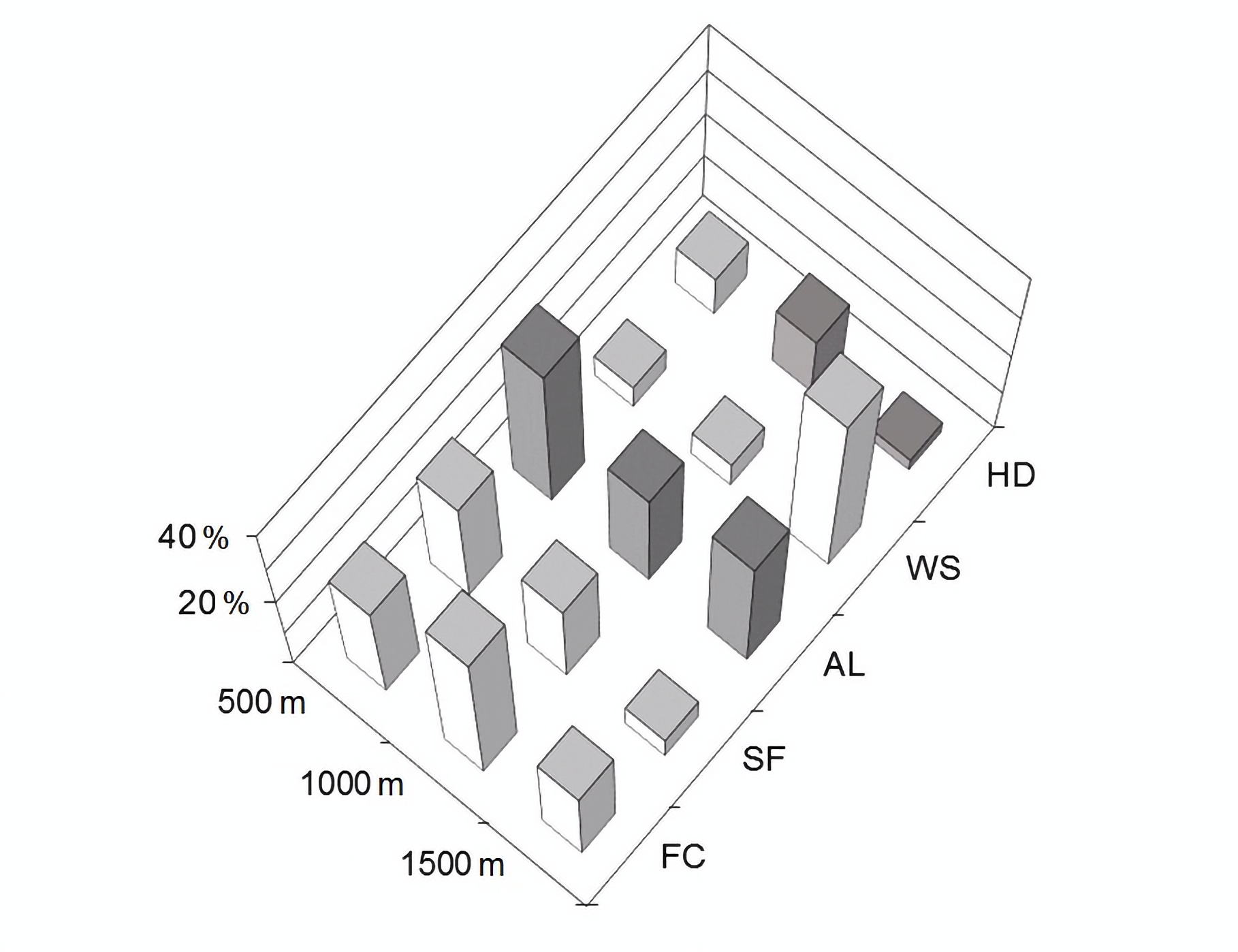
Figure 4
Hierarchical partitioning analysis showing the small vesper bats’ activity variance attributable to univariate correlations with each independent variable, expressed as percentages, in concentric buffers of 500-m, 1,000-m, and 1,500-m radii starting from the sampling points (centroid). The white bars indicate non-parametric positive correlations, and the gray bars negative correlations. FC = forest cover, SF = secondary forest, AL = agricultural land, WS = water surface, HD = human development.
Hierarchical partitioning analysis showing the small vesper bats’ activity variance attributable to univariate correlations with each independent variable, expressed as percentages, in concentric buffers of 500-m, 1,000-m, and 1,500-m radii starting from the sampling points (centroid). The white bars indicate non-parametric positive correlations, and the gray bars negative correlations. FC = forest cover, SF = secondary forest, AL = agricultural land, WS = water surface, HD = human development.
In water sinkholes, the small vesper bats exhibited a comparatively high relative activity in two of the three sampling sites and in most of the recording events, reaching up to 95 % of the time, specifically in AZAR (Supplementary material 6), and on average, it was significantly higher compared to flooded surfaces and forested locations. We also obtained a high rate of buzzes, which we determined by the output of the detector and subsequently by visual inspection of the recordings; these might represent drinking and/or feeding buzzes, but a detailed analysis distinguishing them based on their structure is necessary to make adequate interpretations (see Russoet al.2015). The physiological characteristics of bats demand high amounts of water and prey for reproductive success (MacSwiney et al. 2009; Seiboldet al.2013; López-Gonzálezet al.2016), and there is presence of at least one maternity colony inside the natural protected area, relate to our observations.
-
Russoet
al.2015The buzz of drinking on the wing in echolocating batsEthology, 2015
-
MacSwiney et al. 2009Insectivorous bats activity at cenotes in the Yucatan Peninsula, MéxicoActa Chiropterologica, 2009
-
Seiboldet al.2013Ponds in acidic mountains are more important providing drinking water than insect preyJournal of Zoology, 2013
-
López-Gonzálezet al.2016Activity of insectivorous bats is related to water availability in a highly modified Mexican temperate forestActa Chiropterologica, 2016
Invariably, we observed associations in a positive direction with forest cover and secondary forest. The latter associations could reflect feeding and commuting opportunities. Other studies report similar results, specifically positive correlations between the activity and abundance of this kind of bats and forest cover (native and planted forests) and higher activity in locations at a smaller distance to forested areas (Heim et al.2015; Rodríguez-San Pedro and Simonetti 2015; Fuentes-Montemayoret al.2017; Putet al.2019; Laurindoet al.2020; Falcãoet al.2021).
-
Heim et
al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
-
Rodríguez-San Pedro
and Simonetti 2015The relative influence of forest loss and fragmentation on insectivorous bats: does the type of matrix matter?Landscape Ecology, 2015
-
Fuentes-Montemayoret al.2017Species mobility and landscape context determine the importance of local and landscape-level attributesEcological Applications, 2017
-
Putet al.2019Bats respond negatively to increases in the amount and homogenization of agricultural land coveLandscape Ecology, 2019
-
Laurindoet al.2020Feeding habits define habitat use by bats in an agricultural landscape of the Atlantic ForestRevista Mexicana de Biodiversidad, 2020
-
Falcãoet al.2021Landscape structure shapes activity levels and composition of aerial insectivorous bats at different spatial scalesBiodiversity and Conservation, 2021
Fuentes-Montemayoret al.(2013) make note that the use of forests by aerial insectivorous bats can also be influenced by attributes such as tree density, among other forest structure variables, conditions that Rauchenstein et al. (2022) define as the “suitable foraging habitat”. Vegetation clutter (e. g., forest canopy openness or increased shrub cover) can influence prey abundance (Froidevauxet al.2021; Rauchenstein et al. 2022), restrict mobility and flight maneuverability (Estrada-Villegas et al. 2012; Fuentes-Montemayoret al.2013), and forest maturity (e. g., tree sizes) can be a limiting factor for tree roosting bats (Novella-Fernandez et al. 2022).
-
Fuentes-Montemayoret
al.(2013Fragmented forests in agricultural landscapes: The influence of forest character and landscape context on bats and their insect preyAgriculture, Ecosystems & Environment, 2013
-
Rauchenstein et al. (2022LiDAR metrics predict suitable forest foraging areas of endangered mouse-eared bats (Myotis myotis)Forest Ecology and Management, 2022
-
Froidevauxet al.2021Bat responses to changes in forest composition and prey abundance depend on landscape matrix and stand structureScientific Reports, 2021
-
Estrada-Villegas et
al. 2012Determinants of species evenness in a Neotropical bat ensembleOikos, 2012
-
Fuentes-Montemayoret al.2013Bat responses to changes in forest composition and prey abundance depend on landscape matrix and stand structureScientific Reports, 2021
-
Novella-Fernandez et al.
2022The role of forest structure and composition in driving the distribution of bats in Mediterranean regionsScientific Reports, 2022
We also observed an association in a positive direction with water surfaces, represented in the study area by lakes, water sinkholes and flooded surfaces. The latter elements could represent stepping-stones for many taxa in agriculture-dominated landscapes (Hunter Jr.et al.2017). Even for highly mobile organisms such as bats, a dense network of connecting elements might be beneficial and promote activity in open areas (Heim et al.2015). In the UK, Fuentes-Montemayoret al.(2013; 2017) found positive correlations between the abundance ofMyotisspecies to a larger proportion of surrounding water and decreasing distance between water bodies. In the Atlantic Forest of Brazil, Laurindoet al.(2020) found that the number of captures of insectivorous bats, including Myotisspecies, was significantly associated with the area covered by water bodies within highly fragmented agricultural landscapes.
-
Hunter
Jr.et al.2017Conserving small natural features with large ecological roles: An introduction and definitionBiological Conservation, 2017
-
Heim et
al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
-
Fuentes-Montemayoret al.(2013Fragmented forests in agricultural landscapes: The influence of forest character and landscape context on bats and their insect preyAgriculture, Ecosystems & Environment, 2013
-
Laurindoet
al.(2020Feeding habits define habitat use by bats in an agricultural landscape of the Atlantic ForestRevista Mexicana de Biodiversidad, 2020
We observed associations in a negative direction with agricultural land and human development, except for the latter class in the 500-m buffer, where a small surface characterized it. The latter associations could reflect the effect of an increasingly open area and unfavorable conditions for bats. For instance, Fuentes-Montemayoret al.(2017) found higher activity ofMyotisspecies in forests immersed in areas with a smaller proportion of urban areas. On the other hand, Laurindoet al.(2020) found a negative correlation between the number of captures of insectivorous bats with increased agricultural area.
-
Fuentes-Montemayoret al.(2017Species mobility and landscape context determine the importance of local and landscape-level attributesEcological Applications, 2017
-
Laurindoet al.(2020Feeding habits define habitat use by bats in an agricultural landscape of the Atlantic ForestRevista Mexicana de Biodiversidad, 2020
We explain the associations of relative activity to landscape covers amounts by natural history traits, particularly wing morphology, which is a surrogate for mobility, and echolocation system, which is a surrogate for perceptual range (Frey-Ehrenboldet al.2013; Baderet al.2015; Heim et al.2015; Fuentes-Montemayoret al.2017). The morphological and echolocation system traits of sympatric small vesper bats in the study area point to slow flight with more energetic cost in open areas and short-range high-frequency pulses that are more attenuated in open areas (Frey-Ehrenboldet al.2013; Baderet al.2015; Heim et al.2015). However, Fuentes-Montemayoret al.(2017) suggest that low-mobility species are more influenced by local conditions and the landscape becomes more important for high-mobility species, which perceive the environment at a coarser scale. The latter argument could support our observations, specifically the strong local-level response of small vesper bats to the presence of water sinkholes and the lack of significance of the associations at the landscape level.
-
Frey-Ehrenboldet al.2013Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matricesJournal of Applied Ecology, 2013
-
Baderet al.2015Mobility explains the response of aerial insectivorous bats to anthropogenic habitat change in the NeotropicsBiological Conservation, 2015
-
Heim et al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
-
Fuentes-Montemayoret al.2017Species mobility and landscape context determine the importance of local and landscape-level attributesEcological Applications, 2017
-
Frey-Ehrenboldet al.2013Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matricesJournal of Applied Ecology, 2013
-
Baderet al.2015Mobility explains the response of aerial insectivorous bats to anthropogenic habitat change in the NeotropicsBiological Conservation, 2015
-
Heim et al.2015The importance of landscape elements for bat activity and species richness in agricultural areasPLos One, 2015
-
Fuentes-Montemayoret
al.(2017)Species mobility and landscape context determine the importance of local and landscape-level attributesEcological Applications, 2017
Final remarks. The presence of water sinkholes and environmental conditions such as low wind speed possibly regulating prey availability are important drivers of small vesper bats’ activity in the study area. We found no significant associations for flooded surfaces and forested locations. However, flooded surfaces are particularly relevant to other groups of bats (e. g., Mormoopidae) and vertebrate fauna in the influence zone, such as anurans and migratory birds, representing a seasonally limiting resource.
We observed trends of positive correlation between relative activity to forest cover, secondary forest, and water surface and negative correlation to agricultural land and human development. Although we found no significant associations, we provide explanations based on the argument that small vesper bats react negatively to forest loss and increasingly open area, which we explain by wing morphology and echolocation system traits. Other authors suggest that, for low-mobility species, the landscape may become less important than local-level attributes.
Some of the water sinkholes analyzed in the study area meet the criteria of small natural features having ecological importance that is disproportionate to their size (Hunter Jr. 2017a; Hunter Jr.et al.2017b), as we determined for small vesper bats. The latter consideration is valuable for complementing large-scale conservation through targeted actions and should be further evaluated for its implementation in the area. The water sinkholes are located inside and right on the limits of the natural protected area, in the jurisdiction of federal authorities. The flooded surfaces and forested locations studied are communal goods in the influence zone exposed to human activities and their effects, such as pollution derived of the use of pesticides in the agricultural zones nearby, water exploitation for crops irrigation and cattle drinking supply, forest exploitation with consequent degradation and deforestation.
-
Hunter Jr.
2017Conserving small natural features with large ecological roles: A synthetic overviewBiological Conservation, 2017
-
Hunter Jr.et
al.2017Molecular diet analysis of neotropical bats based on fecal DNA metabarcodingEcology and Evolution, 2021
Finally, we point out that the adequate management of these types of landscape elements should be prioritized in the conservation agenda of the area in an effort of a consensus with ejidatarios (local authorities and ejido members). There is a need for a dialogue that should follow the socio-ecology premises, specifically a transdisciplinary approach to solving environmental problems, looking to benefit biodiversity, the continuity of ecosystem services, and human well-being.
Acknowledgements
We would like to thank the authorities of the Comisión Nacional de Áreas Naturales Protegidas (CONANP) for the permission to work in Lagunas de Montebello National Park. We are also grateful to landowners and local authorities for work permissions. This project was funded partially by Bat Conservation International (grant number SS14012, to the first author), Idea Wild (to the first author), and Instituto Politécnico Nacional, México (grant number SIP 20151481, to the third author). The first author received a scholarship awarded by the Consejo Nacional de Ciencia y Tecnología, México (number 244510). Finally, we would like to thank the anonymous reviewers for their contribution to improving this manuscript.
Literature cited
- Aguiar, L. M. S., and Y. Antonini. 2008. Diet of two sympatric insectivores bats (Chiroptera: Vespertilionidae) in the Cerrado of Central Brazil. Revista Brasileira de Zoologia 25:28-31. Links
- Arroyo-Rodríguez, V., et al. 2016. Landscape composition is more important than landscape configuration for phyllostomid bat assemblages in a fragmented biodiversity hotspot. Biological Conservation 198:84-92. Links
- Bader, E., et al. 2015. Mobility explains the response of aerial insectivorous bats to anthropogenic habitat change in the Neotropics. Biological Conservation 186:97-106. Links
- Barquez, R., and M. Diaz. 2016. Myotis keaysi. In: IUCN 2022. The IUCN Red List of Threatened Species. Version 2022-1. www.iucnredlist.org. Accessed on 7 December 2022. Links
- Bradley, E.S., et al. 1993. Estimation of insect biomass by length and width. The American Midland Naturalist 129:234-240. Links
- Chevan, A., and M. Sutherland. 1991. Hierarchical partitioning. The American Statistician 45:90-96. Links
- Coleman, L.S., et al. 2014. Comparison of radio-telemetric home-range analysis and acoustic detection for little brown bat habitat evaluation. Northeastern Naturalist 21:431-445. Links
- Eastman, J. R. 2012. IDRISI selva tutorial: manual version 17. Clark Labs. Clark University. Massachusetts, U.S.A. Links
- Estrada-Villegas, S., B. J. McGill, and E. K. V. Kalko. 2012. Determinants of species evenness in a Neotropical bat ensemble. Oikos 121:927-941. Links
- Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics 34:487-515. Links
- Falcão, F., et al. 2021. Landscape structure shapes activity levels and composition of aerial insectivorous bats at different spatial scales. Biodiversity and Conservation 30:2545-2564. Links
- Fenton, M. B., and W. Bogdanowicz. 2002. Relationships between external morphology and foraging behaviour: bats in the genus Myotis. Canadian Journal of Zoology 80:1004-1013. Links
- Findley, J. S. 1993. Bats. A Community Perspective. Cambridge University Press. Cambridge, Great Britain. Links
- Fischer, J., and D. B. Lindenmayer. 2007. Landscape modification and habitat fragmentation: a synthesis. Global Ecology and Biogeography 16:265-280. Links
- Frey-Ehrenbold, A., et al. 2013. Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. Journal of Applied Ecology 50:252-261. Links
- Froidevaux, J. S. P., et al. 2021. Bat responses to changes in forest composition and prey abundance depend on landscape matrix and stand structure. Scientific Reports 11:10586. Links
- Fuentes-Montemayor E, et al. 2013. Fragmented forests in agricultural landscapes: The influence of forest character and landscape context on bats and their insect prey. Agriculture, Ecosystems & Environment 172:6-15. Links
- Fuentes-Montemayor, E., et al. 2017. Species mobility and landscape context determine the importance of local and landscape-level attributes. Ecological Applications 27:1541-1554. Links
- Gamboa Alurralde, S., and M. M. Díaz. 2019. Feeding habits of four species of Myotis (Mammalia, Chiroptera) from Argentina. Mammal Research 64:511-518. Links
- Gillies, M. T., and T. J. Wilkes. 1981. Field experiments with a wind tunnel on the flight speed of some West African mosquitoes (Diptera: Culicidae). Bulletin of Entomological Research 71:65-70. Links
- Gonsalves, L., et al. 2013. Mosquito consumption by insectivorous bats: Does size matters? PLos One 8:e77183. Links
- González, E. M., and M. N. Ramírez. 2013. Comunidades vegetales terrestres. Pp. 21-42 in La Biodiversidad en Chiapas: Estudio de Estado. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Gobierno del Estado de Chiapas, Chiapas, México. Links
- Heim, O., et al. 2015. The importance of landscape elements for bat activity and species richness in agricultural areas. PLos One 10:e0134443. Links
- Hunter, M. L. 2017. Conserving small natural features with large ecological roles: An introduction and definition. Biological Conservation 211:1-2. Links
- Hunter, M. L., et al. 2017. Conserving small natural features with large ecological roles: A synthetic overview. Biological Conservation 211:88-95. Links
- Ingala, M. R., et al. 2021. Molecular diet analysis of neotropical bats based on fecal DNA metabarcoding. Ecology and Evolution 11:7474-7491. Links
- Jung, K., and E. K. V. Kalko. 2011. Adaptability and vulnerability of high flying Neotropical aerial insectivorous bats to urbanization. Diversity and Distributions 17:262-274. Links
- Jones, G., and J. Rydell. 2003. Attack and defense: Interactions between echolocating bats and their insect prey. Pp. 301-345, in Bat Ecology (Kunz, T. H., and M. B. Fenton, eds.). The University of Chicago Press. Chicago, U.S.A. Links
- Lacki, M. J., J. P. Hayes, and A. Kurta (eds.). 2007. Bats in Forests: Conservation and Management. The Johns Hopkins University Press. Baltimore, U.S.A. Links
- Laurindo, R. S., et al. 2020. Feeding habits define habitat use by bats in an agricultural landscape of the Atlantic Forest. Revista Mexicana de Biodiversidad 91:e913223. Links
- López-González, C., et al. 2016. Activity of insectivorous bats is related to water availability in a highly modified Mexican temperate forest. Acta Chiropterologica 18:409-421. Links
- Lu, D., and Q. Weng. 2007. A survey of image classification methods and techniques for improving classification performance. International Journal of Remote Sensing 28:823-870. Links
- Mac Nally, R. 2000. Regression and model-building in conservation biology, biogeography and ecology: the distinction between - and reconcilitation of - ‘predictive’ and ‘explanatory’ models. Biodiversity and Conservation 9:655-671. Links
- Mac Nally, R. 2002. Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables. Biodiversity and Conservation 1:1397-1401. Links
- MacSwiney M. C., F. M. Clarke, and P. A. Racey. 2009. Insectivorous bats activity at cenotes in the Yucatan Peninsula, México. Acta Chiropterologica 11:139-147. Links
- Martino, A. M. G., D. Borges, and J. M. Nassar. 2019. Activity records of the endangered Paraguaná moustached bat, Pteronotus paraguanensis, in the main vegetation types of the Paraguaná Peninsula, Venezuela. Acta Chiropterologica 21:165-174. Links
- Mendes, E. S., et al. 2017. Bat richness and activity in heterogeneous landscapes: guild-specific and scale-dependent? Landscape Ecology 32:295-311. Links
- Miller, B. W. 2001. A method for determining relative activity of free flying bats using a new activity index for acoustic monitoring. Acta Chiropterologica 3:93-105. Links
- Miller, B., et al. 2016. Rhogeessa tumida. In: IUCN 2022. The IUCN Red List of Threatened Species. Version 2022-1. www.iucnredlist.org. Accessed on 7 December 2022. Links
- Norberg, U. M., and J. M. V. Rayner. 1987. Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philosophical Transactions of the Royal Society B: Biological Sciences 316:335-427. Links
- Novella-Fernandez, R., et al. 2022. The role of forest structure and composition in driving the distribution of bats in Mediterranean regions. Scientific Reports 12:3224. Links
- O’Farrell, M. J., and B. W. Miller. 1999. Use of vocal signatures for the inventory of free-flying Neotropical bats. Biotropica 31:507-516. Links
- Olea, P. P., P. Mateo-Tomás, and Á. de Frutos. 2010. Estimating and modelling bias of the hierarchichal partitioning public-domain software: implications in environmental management and conservation. Plos One 5:e11698. Links
- Orozco-Lugo, C. L., et al. 2013. Descripción de los pulsos de ecolocalización de once especies de murciélagos insectívoros aéreos de una selva baja caducifolia en Morelos, México. Therya 4:33-46. Links
- Owen, S. F., et al. 2003. Home-range size and habitat used by the northern Myotis (Myotis septentrionalis). The American Midland Naturalist 150:352-359. Links
- Parreira Peixoto, F., P. H. Pereira Braga, and P. Mendes. 2018. A synthesis of ecological and evolutionary determinants of bat diversity across spatial scales. BMC Ecology 18:18. Links
- Pettersson, L. 2002. The properties of sound and bat detectors. Pp. 9-12 in Bat Echolocation Research: Tools Techniques and Analysis (Brigham, R. M., et al., eds.). Bat Conservation International. Texas, U.S.A. Links
- Popescu, V. D., and P. Gibbs. 2010. Landscape ecology and GIS methods. Pp. 339-360, in Amphibian Ecology and Conservation: A Handbook of Techniques (Kenneth Jr., C., ed.). Oxford University Press. New York, U.S.A. Links
- Puig-Montserrat, X., et al. 2020. Bats actively prey on mosquitoes and other deleterious insects in rice paddies: Potential impact on human health and agriculture. Pest Management Science 76:3759-3769. Links
- Put, J. E., L. Fahrig, and G. W. Mitchell. 2019. Bats respond negatively to increases in the amount and homogenization of agricultural land cover. Landscape Ecology 34:1889-1903. Links
- Queiroz de Oliveira, L., et al. 2015. Activity of the insectivorous bat Pteronotus parnellii relative to insect resources and vegetation structure. Journal of Mammalogy 96:1036-1044. Links
- Rauchenstein, K., et al. 2022. LiDAR metrics predict suitable forest foraging areas of endangered mouse-eared bats (Myotis myotis). Forest Ecology and Management 515:120-210. Links
- R Core Team. 2019. R: A language and environment for statistical computing. Foundation for Statistical Computing. http://R-project.org . Accessed 19 December 2019. Links
- Radford, J. Q., and A. F. Bennett. 2007. The relative importance of landscape properties for forest birds in agricultural environments. Journal of Applied Ecology 44:737-747. Links
- Rodríguez-San Pedro, A., and J. A. Simonetti. 2015. The relative influence of forest loss and fragmentation on insectivorous bats: does the type of matrix matter? Landscape Ecology 30:1561-1572. Links
- Russo, D., et al. 2015. The buzz of drinking on the wing in echolocating bats. Ethology 122:226-235. Links
- Rutledge, C. R. 2008. Mosquitoes (Diptera: Culicidae). Pp. 2476-2483, in Encyclopedia of Entomology (Capinera, J. L., ed.). Springer. Dordrecht, Netherlands. Links
- Rydell, J., et al. 2017. Testing the performances of automated identification of bat echolocation calls: A request for prudence. Ecological Indicators 78:416-420. Links
- Schnitzler, H.-U., and E. K. V. Kalko. 2001. Echolocation by Insect-Eating Bats. Bioscience 51:557-569. Links
- Salinas-Ramos, V. B., et al. 2015. Dietary overlap and seasonality in three species of mormoopid bats from a tropical dry forest. Molecular Ecology 24:5296-5307. Links
- Segura-Trujillo, C. A., M. R. Willig, and S. T. Álvarez-Castañeda. 2018. Correspondence between ecomorphotype and use of arthropod resources by bats of the genus Myotis. Journal of Mammalogy 99:659-667. Links
- Seibold, S., et al. 2013. Ponds in acidic mountains are more important providing drinking water than insect prey. Journal of Zoology 290:302-308. Links
- Solari, S. 2019. Myotis nigricans. In: IUCN 2022. The IUCN Red List of Threatened Species. Version 2022-1. www.iucnredlist.org. Accessed on 7 December 2022. Links
- Verbyla, D. L. 1995. Satellite remote sensing of natural resources. CRC Press. Boca Raton, U.S.A. Links
- Walsh, C., and R. Mac Nally. 2022. Package ‘hier.part’, version 1.0-6. https://cran.r-project.org/web/packages/hier.part/hier.part.pdf . Accessed 21 October 2022. Links
- Whitaker, J. O. 2004. Prey selection in a temperate zone insectivorous bat community. Journal of Mammalogy 85:460-469. Links
- Wickramasinghe, L. P., et al. 2004. Abundance and species richness of nocturnal insects on organic and conventional farms: effects of agricultural intensification on bat foraging. Conservation Biology 18:1283-1292. Links
- Williams-Guillén, K., and I. Perfecto. 2011. Ensemble composition and activity levels of insectivorous bats in response to management intensification in coffee agroforestry systems. Plos One 6:e16502. Links
- Zamora-Gutiérrez, V., et al. 2016. Acoustic identification of Mexican bats based on taxonomic and ecological constraints on call design. Methods in Ecology and Evolution 7:1082-1091. Links