Taxonomy and systematics
Contribution to the systematic value of the leaf trichomes in the Cordiaceae (Boraginales)
Contribución al valor sistemático de los tricomas foliares en las Cordiaceae (Boraginales)
Nazari, Hamida
 http://orcid.org/0009-0008-0779-9069
http://orcid.org/0009-0008-0779-9069
Ghahremaninejad, Farrokhb*
http://orcid.org/0000-0001-5860-9976
http://orcid.org/0009-0008-0779-9069Ghahremaninejad, Farrokhb*
http://orcid.org/0000-0001-5860-9976
-
Publication dates-
September 26, 2025
, 2025
- Article in PDF
- Article in XML
- Automatic translation
- Send this article by e-mail
- Share this article +
Abstract
Leaf trichomes are vital for plant physiology, defense, and ecological interactions. This study utilized scanning electron microscopy to examine the leaf trichomes of 7 species in the family Cordiaceae, including 5 species from the genus Cordia and 2 from Varronia. Various types of trichomes, including glandular, non-glandular, and stellate, were observed. Our findings highlight differences in trichome types, density, and distribution both within and between genera. This research enhances the understanding of anatomical diversity and the significance of leaf trichomes in Cordiaceae, providing a foundation for future studies in plant biology. The high diversity of trichome structure in the studied species indicates their importance for their classification. Therefore, the results of this study provide a useful insight into the diversity of trichomes and their potential as a taxonomic tool.
Keywords::
Indumentum, Glandular, Non-glandular, Stellate trichomes, Taxonomy
Introduction
The family Cordiaceae in Boraginales includes 2 large genera, Cordia L. and Varronia P. Browne with a total of 368 accepted species (POWO, 2024) or 350 (APG 4, 2016). Its members are usually shrubs or trees that are distributed in tropical and subtropical regions (Luebert et al., 2016). The classification of this family and its subfamilies has been debated by botanists for years. In most references Cordiaceae has been recognized as a family or as part of Heliotropiaceae and Boraginaceae. Finally, based on the results of Luebert et al. (2016), this family was accepted as a separate family in Boraginales. Cordiaceae is the only family in Boraginales with plicate cotyledons and generally a twice-dichotomous style with 4 stigmatic branches (Luebert et al., 2016). Johnston (1951) introduced the genus Cordia with 5-7 sections for Cordiaceae and molecular work placed Varronia as the sister genus of Cordia (Miller & Gottschling, 2007). Species of Cordia are found principally in tropical and subtropical regions of the American, Asian, and African continents. In this genus, there are many species cultivated as ornamentals, and for wood and medicinal applications, with extensive use by traditional communities (Matias et al., 2015). Varronia is a Neotropical genus with most of its species naturally distributed in Brazil (Silva & Melo, 2019).
-
POWO, 2024Plants of the World. Online, 2024
-
APG 4, 2016An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IIIBotanical Journal of the Linnean Society, 2016
-
Luebert et al., 2016Familial classification of the BoraginalesTaxon, 2016
-
Luebert et al. (2016)Familial classification of the BoraginalesTaxon, 2016
-
Luebert et al., 2016Familial classification of the BoraginalesTaxon, 2016
-
Johnston (1951)Studies in the Boraginaceae XX. Representatives of three subfamilies in eastern AsiaJournal of the Arnold Arboretum, 1951
-
Miller & Gottschling, 2007Generic classification in the Cordiaceae (Boraginales): Resurrection of the genus Varronia R.BrTaxon, 2007
-
Matias et al., 2015The genus Cordia: Botanists, ethno, chemical and pharmacological aspectsRevista Brasileira de Farmacognosia, 2015
-
Silva & Melo, 2019A new species and a new record of Varronia (Cordiaceae) from BrazilSystematic Botany, 2019
Leaf trichomes are specialized structures that grow on the epidermis of leaves. They play an important role in how plants adapt to their environment and interact with it (Wagner, 1991); also, they are a powerful tool in solving taxonomic problems (e.g., Ghahremaninejad et al., 2012). Trichomes play a significant role in the leaf texture of Cordiaceae species, contributing to their diagnostic value within the family. Studies have shown that the family possesses both glandular and non-glandular leaf trichomes, which influence their rough texture (Amer et al., 2016). Notably, in some regions, the abrasive surface of these leaves (including those of Cordia sebestena L.) has led to their traditional use as natural sandpaper (Flowers of India, 2025). Cordiaceae exhibits a wide range of leaf trichome morphologies, yet comprehensive studies elucidating their diversity and functional significance remain limited. Trichome morphology, density, and distribution are known to vary extensively even within closely related taxa, reflecting adaptation to diverse ecological niches and selective pressures (Rodríguez & Healey, 2000). In addition to their functional roles, the structural and functional diversity of leaf indument in Cordiaceae are indispensable taxonomic tools. Trichome characteristics often serve as apomorphic characters that assist botanists in resolving complex taxonomic issues (e.g., Beilstein et al., 2006; Ghahremaninejad, 2004; Kong & Hong, 2019; Steyn & Van Wyk, 2021). However, many studies within this family have paid insufficient attention to trichome features (e.g., Amer et al., 2016; Taroda, 1984). Trichome morphology exhibits remarkable diversity across the plant kingdom, playing a significant role in the taxonomy and ecology of various plant families, including the Boraginaceae and Cordiaceae (Selvi & Bigazzi, 2001). Limited studies on Cordiaceae trichomes reveal a high diversity of trichomes both among closely related species (Tölke et al., 2013) and at the genus level (Silva et al., 2023). This diversity in trichome morphology suggests intricate evolutionary adaptations that may serve various ecological functions.
-
Wagner, 1991Secreting glandular trichomes: more than just hairsPlant Physiology, 1991
-
Ghahremaninejad et al., 2012Leaf epidermal features of Salix species (Salicaceae) and their significanceAmerican Journal of Botany, 2012
-
Amer et al., 2016Taxonomic revision of genus Cordia L. (Boraginaceae) in EgyptEgypt Journal of Botany, 2016
-
Flowers of India, 2025Scarlet Cordia (Cordia sebestena)Flowers of India, 2025
-
Rodríguez & Healey, 2000Morphological diversity of trichomes in Loasaceae and its systematic significanceAnnals of the Missouri Botanical Garden, 2000
-
Beilstein et al., 2006Brassicaceae phylogeny and trichome evolutionAmerican Journal of Botany, 2006
-
Ghahremaninejad, 2004Value of trichome characteristics for separating of bifurcating Astragalus (Fabaceae) on sectional levelTurkish Journal of Botany, 2004
-
Kong & Hong, 2019Leaf micromorphology of the Persicaria sect. Cephalophilon (Polygonaceae) and its systematic re-evaluationPhytotaxa, 2019
-
Steyn & Van Wyk, 2021Taxonomic significance of trichomes in the genus Acanthopsis Harv. (Acanthaceae, tribe Acantheae)Adansonia, 2021
-
Amer et al., 2016Taxonomic revision of genus Cordia L. (Boraginaceae) in EgyptEgypt Journal of Botany, 2016
-
Taroda, 1984Taxonomic studies on Brazilian species of Cordia L. (Boraginaceae), 1984
-
Selvi & Bigazzi, 2001Leaf surface and anatomy in Boraginaceae tribe Boragineae with respect to ecology and taxonomyFlora, 2001
-
Tölke et al., 2013Leaf anatomy with emphasis on separation of two species of Varronia P.Br. (Cordiaceae) of the Brazilian semi-arid regionBrazilian Journal of Botany, 2013
-
Silva et al., 2023A contribution of leaf anatomy to the taxonomy of Varronia P. Browne (Cordiaceae, Boraginales)Flora, 2023
Given the existing literature on Cordia and Varronia, trichomes have critical taxonomic features in Cordiaceae, particularly for differentiating closely related genera. This study aims to examine the trichome morphology of both genera using SEM to clarify their taxonomic relationships. Scanning electron microscopy enhances trichome trait comparison by providing greater detail (Gonçales et al., 2023). We anticipated identifying distinct trichome characteristics to resolve existing taxonomic ambiguities. By focusing on these morphological characters, we seek to enhance the understanding of Cordiaceae taxonomy.
-
Gonçales et al., 2023Floral indumentum of Miconia: In search of diagnostic characters for the tribe MiconieaeRevista Mexicana de Biodiversidad, 2023
Material and methods
Samples were obtained from Herbarium specimens of the Naturhistorisches Museum Wien (W) and Botanical Museum Berlin-Dahlem (B), acronyms according to Thiers (2025). Trichome morphology was studied using SEM at the Natural History Museum (NHM), Vienna. Leaves were soaked in 40-degree water for 10 minutes to restore trichomes and then air-dried for 1 hour. Leaf samples were attached to SEM stubs with conductive adhesive tabs and coated with a thin layer of gold for conductivity, applied with a sputter coater. Imaging was performed with a JEOL JXA 6610LV SEM at the Vienna Museum. Digital images from various regions were captured, documenting trichome characteristics such as type, shape, size, density, and distribution for each species. Additionally, a voucher table listing the information and codes of the specimens used was prepared (Table 1).
-
Thiers (2025)Index Herbariorum: a global directory of public herbaria and associated staff (Continuously updated)New York Botanical Garden’s Virtual Herbarium, 2025
Table 1
The voucher table of studied specimens.
The voucher table of studied specimens.
| No. | Species | Locality | Collector and number | QR code | Herbarium acronym | Type of trichome |
|---|---|---|---|---|---|---|
| 1 | Cordia africana Lam. | Ethiopia | U. i. 285 | W0254121 | W | Non-glandular |
| 2 | Cordia alliodora (Ruiz & Pav.) Oken | Brazil | s.c. 893 | W0058028 | W | Non-glandular |
| 3 | Cordia bicolor A.DC. | Brazil | Schomburgk 678 | W0010617 | W | Stellate |
| 4 | Cordia decandra Hook. & Arn. | Chile | K.H. & Rechinger 63378 | W0254122 | W | Non-glandular |
| 5 | Cordia myxa L. | Greece | Rainer KARL | W0254123 | W | Non-glandular |
| 6 | Varronia bullata L. | - | - | W0254157 | W | Glandular, non-glandular |
| 7 | Varronia guanacastensis (Standl.) J.S.Mill. | Costa Rica | Zamora et al. 6340 | B101137807 | B | Glandular, non-glandular (short pyramidal shape) |
Results
Electron microscopy studies of 5 species of Cordia and 2 species of Varronia illustrate the diversity in the size, shape, and distribution of trichomes in Cordiaceae. Variation was observed both within and between genera. Two main types of trichomes were identified among the studied species, glandular and non-glandular. Glandular trichomes were observed only in Varronia species, while non-glandular were observed in species of both genera. In addition, a special type of stellate trichome, which mostly has more than 10 arms, as a first-time report, was observed in Cordia bicolor A. DC.
Among the investigated species, glandular trichomes with a size of 100 to 120 μm were observed only in Varronia bullata L. and V. guanacastensis (Standl.) J.J. Mill. (Fig. 1). A type of trichome was observed in Cordia myxa L., which is bag-like and different from non- glandular trichomes. The samples used in this research were herbarium specimens, but for accurate diagnosis of this trichome, fresh material is needed.
Thumbnail
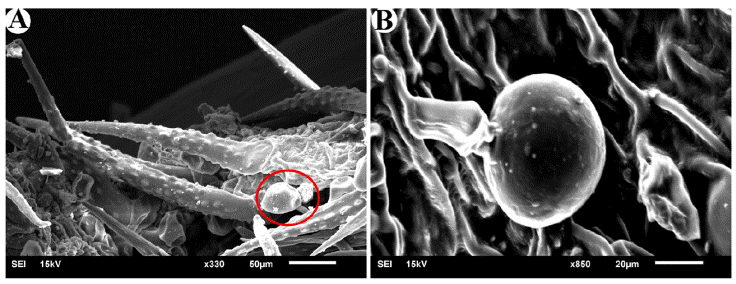
Figure 1
Glandular trichomes. A, Glandular trichomes (Icircled) next to non-glandular trichomes in Varronia bullata; B, glandular trichomes in Varronia guanacastensis.
Glandular trichomes. A, Glandular trichomes (Icircled) next to non-glandular trichomes in Varronia bullata; B, glandular trichomes in Varronia guanacastensis.
In all 7 studied species, non-glandular trichomes were observed on both adaxial and abaxial leaf surfaces. In Cordia africana Lam., the upper surface of the leaf has scattered short trichomes that exist in the form of small protrusions distant from one another, while the back surface of the leaf in this species (Fig. 2A) has depressions where different trichomes are densely present in these depressions. In C. alliodora (Ruiz & Pav.) Oken., simple trichomes are observed on both surfaces. On the abaxial surface of the leaf, in addition to this type of trichome, there are smaller and curved trichomes that do not have small projections on their surface (Fig. 2B). On the abaxial surface of C. decandra Hook. & Arn. leaves, in addition to short and long simple trichomes, which are smooth and sometimes curved, horn-shaped hard trichomes are also observed (Fig. 2C). In C. myxa, in addition to the specific type of trichomes mentioned (referring to glandular trichomes), very small trichomes that include a widened base and a protrusion (Fig. 2D) are observed in nearly regular rows on the adaxial surface of the leaf. On the abaxial surface, long trichomes with small projections are observed. Varronia bullata also has simple trichomes, which are similar to the trichomes of the mentioned species, except that on the adaxial surface, trichomes are completely dormant (Fig. 3A), but on the abaxial surface, in addition to greater diversity and density, some standing or twisted trichomes are also seen (Fig. 3B). In Varronia guanacastensis, a special type of trichome was observed, which covered the adaxial surface of the leaf in addition to many glandular trichomes. These trichomes are short and have a conical end (Fig. 3C). On the abaxial surface, a type of horn-like non-glandular trichome is also seen (Fig. 3D).
Thumbnail
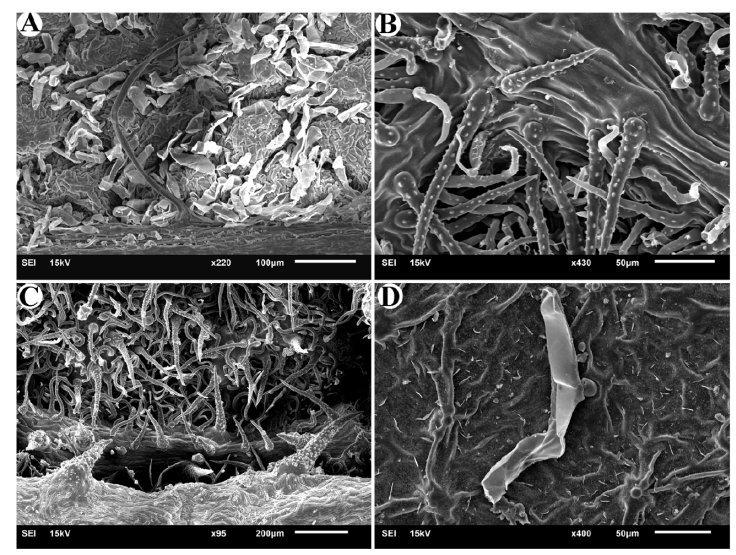
Figure 2
Non-glandular trichomes in studied species of Cordia. A, Abaxial surface in Cordia africana; B, abaxial surface in Cordia alliodora; C, abaxial surface in Cordia decandra; D, abaxial surface in Cordia myxa,
Non-glandular trichomes in studied species of Cordia. A, Abaxial surface in Cordia africana; B, abaxial surface in Cordia alliodora; C, abaxial surface in Cordia decandra; D, abaxial surface in Cordia myxa,
Thumbnail
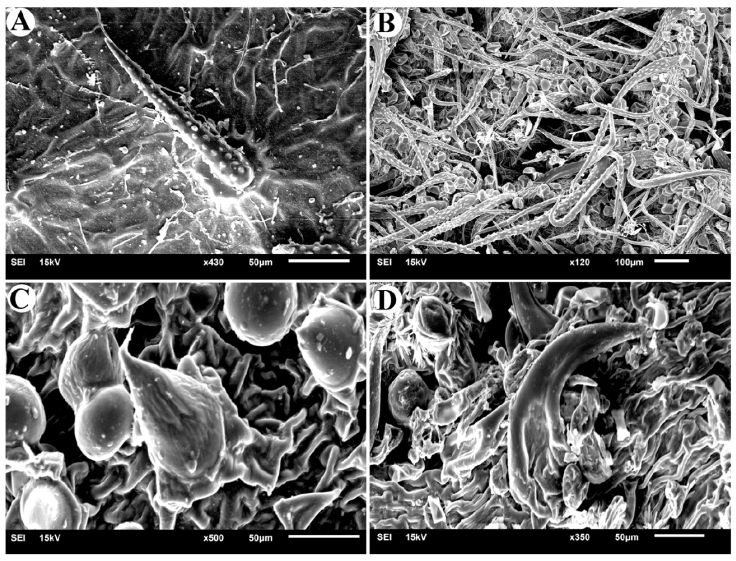
Figure 3
Non-glandular trichomes in studied species of Varronia. A, Adaxial surface in Varronia bullata; B, abaxial surface in Varronia bullata; C, adaxial surface in Varronia guanacastensis; D, abaxial surface in Varronia guanacastensis.
Non-glandular trichomes in studied species of Varronia. A, Adaxial surface in Varronia bullata; B, abaxial surface in Varronia bullata; C, adaxial surface in Varronia guanacastensis; D, abaxial surface in Varronia guanacastensis.
Stellate trichomes are a type of branched trichome that have several arms arising from a common base (either stalked or sessile). This type of trichome was observed only in Cordia bicolor. The number of arms of these trichomes is between 10 and 17, and the length of each arm is between 50 and 100 μm. Figure 4 shows the studied specimen and the observed stellate trichome. The distribution and density of trichomes on the surface of the leaf can be seen with the naked eye most of the time. Figure 5 shows the leaves of the studied species where trichome density can be seen on their surface.
Thumbnail

Figure 4
Stellate trichome in Cordia bicolor. A, studied specimen (W0010617); B, leaf surface visible with a normal magnification (X4); C, stellate trichomes are densely distributed on the leaf surface; D, stellate trichome with more than 10 arms.
Stellate trichome in Cordia bicolor. A, studied specimen (W0010617); B, leaf surface visible with a normal magnification (X4); C, stellate trichomes are densely distributed on the leaf surface; D, stellate trichome with more than 10 arms.
Thumbnail
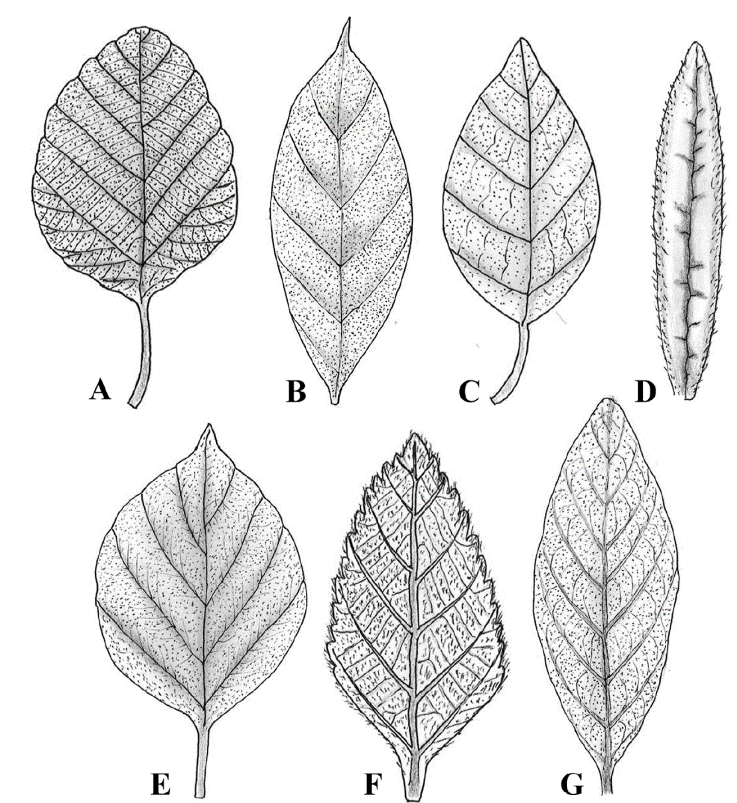
Figure 5
The distribution of trichomes on the adaxial leaf surface of the studied species of Cordiaceae. A, Cordia africana; B, Cordia alliodora; C, Cordia bicolor; D, Cordia decandra; E, Cordia myxa; F, Varronia bullata; G, Varronia guanacastensis.
The distribution of trichomes on the adaxial leaf surface of the studied species of Cordiaceae. A, Cordia africana; B, Cordia alliodora; C, Cordia bicolor; D, Cordia decandra; E, Cordia myxa; F, Varronia bullata; G, Varronia guanacastensis.
Microscopic examination of these 7 species revealed that the adaxial leaf surface generally has a lower density of trichomes compared to the abaxial surface, which has a higher density. On the adaxial surface of the leaf of Cordia africana, there are trichomes in the form of small protrusions that are sparse, while on the abaxial surface, the trichomes have a different shape and are densely placed inside the depressions, while long trichomes are rarely seen. In Cordia alliodora, C. bicolor, C. decandra, and C. myxa the type of trichome is similar on both surfaces, with the difference that they are scattered on the adaxial side and dense on the abaxial side. In the case of the 2 Varronia species, the trichomes are scattered on the adaxial surface of V. bullata and dense on the abaxial surface, but in V. guanacastensis, the trichomes are dense on both leaf surfaces, with the difference being that the type of trichome is different between the surfaces. Figure 6 shows the difference between trichome distribution on adaxial and abaxial leaf surfaces in all studied species.
Thumbnail

Figure 6
Differences in trichome distribution on adaxial and abaxial surfaces of some Cordia species. A, Cordia alliodora; B, Cordia myxa; C, Cordia africana.
Differences in trichome distribution on adaxial and abaxial surfaces of some Cordia species. A, Cordia alliodora; B, Cordia myxa; C, Cordia africana.
Discussion
Our study on leaf trichomes within Cordiaceae highlights significant morphological diversity, underscoring the diagnostic value of trichomes in taxonomy. Trichomes were categorized as either glandular (exclusively in Varronia species) or non-glandular (present in both genera), establishing clear taxonomic distinctions between the 2 genera. The restriction of glandular trichomes to Varronia supports the value of trichomes as reliable taxonomic markers, reinforcing molecular evidence that separates Varronia from Cordia. Despite the limited number of species examined, these findings offer a foundation for further studies across Cordiaceae. A notable finding was the discovery of stellate trichomes in Cordia bicolor, a first report for this species and probably rare within Boraginales. Stellate trichomes, characterized by multiple arms emerging from a common base, were observed solely in Cordia bicolor after an examination of over 100 Boraginales species (details in preparation). Figure 5 illustrates this unique stellate trichome, underscoring its taxonomic significance and its potential adaptive function.
Previous studies align with our findings on trichome diversity. Silva et al. (2023) examined leaf anatomy in 10 Varronia species from Brazil, identifying secretory and non-secretory trichomes as key taxonomic indicators. Similarly, Tölke et al. (2013) demonstrated variation in glandular and non-glandular trichomes among V. globosa Jacq. and V. leucocephala (Moric.) J.S. Mill., with unique glandular types and micropapillae-covered trichomes distinguishing each species. Demétrio et al. (2020) reported distinctive non-glandular trichomes and a single glandular type in V. polycephala Lam., a medicinal species, which also contained cystoliths and crystal sand idioblasts. These variations highlight trichomes as valuable taxonomic features and adaptive characters. While this study suggests a difference between Varronia and Cordia, the limited scope relative to the genera’s size precludes definitive conclusions. Further sampling of more species is needed to confirm this distinction in leaf trichomes.
-
Silva et al. (2023)A contribution of leaf anatomy to the taxonomy of Varronia P. Browne (Cordiaceae, Boraginales)Flora, 2023
-
Tölke et al. (2013)Leaf anatomy with emphasis on separation of two species of Varronia P.Br. (Cordiaceae) of the Brazilian semi-arid regionBrazilian Journal of Botany, 2013
-
Demétrio et al. (2020)Leaf anatomy of Varronia polycephala Lam. (Cordiaceae)Flora, 2020
Moreover, the distribution of trichomes, denser on the abaxial leaf surfaces, suggests adaptations to environmental pressures and provides additional taxonomic insight. Consistent trichome types on particular leaf surfaces across species indicate conserved characters, useful for classification within the family. This diversity, particularly in non-glandular trichomes that vary in shape, size, and density, emphasizes the evolutionary lability of trichomes and their dual roles in adaptation and taxonomy.
Our use of SEM enabled detailed trichome analysis, offering precise structural insights and enhancing taxonomic resolution. Nazari and Ghahremaninejad (2024, 2025) also found that trichome examination can verify generic separations within Boraginales, as demonstrated in their study on Heliotropiaceae. Collectively, these findings underscore the morphological and taxonomic significance of trichomes, establishing them as key indicators in botanical classification.
-
Nazari and Ghahremaninejad (2024A SEM study of leaf trichomes in HeliotropiaceaeIranian Journal of Botany, 2024
Acknowledgments
We sincerely thank all the invaluable contributors to this work. Special thanks to Heimo Rainer, head of the Botany Department at the Natural History Museum of Vienna, for his extensive support. Additionally, we appreciate Astrid Hille, Andreas Berger, Tanja Schuster, and Johannes Walter from the same department for their assistance. We are also grateful to Wenke Wegner, a SEM specialist, for supervising the microscopic imaging of the fragments. Our thanks extend to W herbarium and B herbarium for sharing their extensive botanical resources.
References
- Amer, W. M., Hamdy, R. S., & Mahdy, R. A. (2016). Taxonomic revision of genus Cordia L. (Boraginaceae) in Egypt. Egypt Journal of Botany, 6th International Conference, Menoufia University, 375-408. Links
- APG IV. (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society, 181, 1-20. https://doi.org/10.1111/boj.12385 Links
- Beilstein, M. A., Al-Shehbaz, I. A., & Kellogg, E. A. (2006). Brassicaceae phylogeny and trichome evolution. American Journal of Botany, 93, 607-619. https://doi.org/10.3732/ajb.93.4.607 Links
- Demétrio, A. M., Lusa, M. G., Lima, D. F., & Rodrigues, C. (2020). Leaf anatomy of Varronia polycephala Lam. (Cordiaceae). Flora, 271, 151677. https://doi.org/10.1016/j.flora.2020.151677 Links
- Flowers of India. (2025). Scarlet Cordia (Cordia sebestena). Retrieved March 7, 2025, from Retrieved March 7, 2025, from https://www.flowersofindia.net/catalog/slides/Scarlet%20Cordia.html Links
- Ghahremaninejad, F. (2004). Value of trichome characteristics for separating of bifurcating Astragalus (Fabaceae) on sectional level. Turkish Journal of Botany, 28, 241-245. Links
- Ghahremaninejad, F., Khalili, Z., Maassoum, A. A., Mirzaie- Nodoushan, H., & Riahi, M. (2012). Leaf epidermal features of Salix species (Salicaceae) and their significance. American Journal of Botany, 99, 769-777. https://doi.org/10.3732/ajb.1100019 Links
- Gonçales, R., Do Carmo, A. A. O., dos Santos, A. F., & de Souza, L. A. (2023). Floral indumentum of Miconia: In search of diagnostic characters for the tribe Miconieae. Revista Mexicana de Biodiversidad, 94, e945272. https://doi.org/10.22201/ib.20078706e.2023.94.5272 Links
- Johnston, I. M. (1951). Studies in the Boraginaceae XX. Representatives of three subfamilies in eastern Asia. Journal of the Arnold Arboretum, 32, 99-122. https://doi.org/10.5962/bhl.part.9730 Links
- Kong, M. J., & Hong, S. P. (2019). Leaf micromorphology of the Persicaria sect. Cephalophilon (Polygonaceae) and its systematic re-evaluation. Phytotaxa, 391, 167-184. https://doi.org/10.11646/phytotaxa.391.3.1 Links
- Luebert, F., Cecchi, L., Frohlich, M. W., Gottschling, M., Guilliams, C. M., Hasenstab-Lehman, K. E., et al. (2016). Familial classification of the Boraginales. Taxon, 65, 502-522. https://doi.org/10.12705/653.5 Links
- Matias, E. F. F., Alves, E. F., Silva, M. K. N., Carvalho, V. R. A., Coutinho, H. D. M., & da Costa, J. G. M. (2015). The genus Cordia: Botanists, ethno, chemical and pharmacological aspects. Revista Brasileira de Farmacognosia, 25, 542-552. https://doi.org/10.1016/j.bjp.2015.05.012 Links
- Miller, J. S., & Gottschling, M. (2007). Generic classification in the Cordiaceae (Boraginales): Resurrection of the genus Varronia R.Br. Taxon, 56, 163-169. https://doi.org/10.2307/25065747 Links
- Nazari, H., & Ghahremaninejad, F. (2024). A SEM study of leaf trichomes in Heliotropiaceae. Iranian Journal of Botany, 30, 173-183. https://doi.org/10.22092/ijb.2024.366196.1475 Links
- POWO. (2024). Plants of the World. Online. https://powo.science.kew.org/ Links
- Rodríguez, A., & Healey, P. L. (2000). Morphological diversity of trichomes in Loasaceae and its systematic significance. Annals of the Missouri Botanical Garden, 87, 71-86. https://doi.org/10.1093/aobpla/plab071 Links
- Selvi, F., & Bigazzi, M. (2001). Leaf surface and anatomy in Boraginaceae tribe Boragineae with respect to ecology and taxonomy. Flora, 196, 269-285. https://doi.org/10.1016/S0367-2530(17)30056-7 Links
- Silva, T. S., & de Melo, J. I. M. (2019). A new species and a new record of Varronia (Cordiaceae) from Brazil. Systematic Botany, 44, 692-696. https://doi.org/10.1600/036364419x15620113920752 Links
- Silva, T. S., Tölke, E. E. D., & de Melo, J. I. M. (2023). A contribution of leaf anatomy to the taxonomy of Varronia P. Browne (Cordiaceae, Boraginales). Flora, 304, 152299. https://doi.org/10.1016/j.flora.2023.152299 Links
- Steyn, H. M., & Van Wyk, A. E. (2021). Taxonomic significance of trichomes in the genus Acanthopsis Harv. (Acanthaceae, tribe Acantheae). Adansonia, 43, 163-176. https://doi.org/10.5252/adansonia2021v43a14 Links
- Taroda, N. (1984). Taxonomic studies on Brazilian species of Cordia L. (Boraginaceae) (Ph.D. Thesis). University of St. Andrews. St. Andrews, UK. Links
- Thiers, B. M. (2025). Index Herbariorum: a global directory of public herbaria and associated staff (Continuously updated). New York Botanical Garden’s Virtual Herbarium. http://sweetgum.nybg.org/ih/ Links
- Tölke, E. E. D., de Melo, J. I. M., Carmello-Guerreiro, S. M., & Lacchia, A. P. S. (2013). Leaf anatomy with emphasis on separation of two species of Varronia P.Br. (Cordiaceae) of the Brazilian semi-arid region. Brazilian Journal of Botany, 36, 189-201. https://doi.org/10.1007/s40415-013-0022-5 Links
- Wagner, G. J. (1991). Secreting glandular trichomes: more than just hairs. Plant Physiology, 96, 675-679. https://doi.org/10.1104/pp.96.3.675 Links