Research notes
Effect of environmental light conditions on the growth of the coral Orbicella faveolata in the Mexican Caribbean
Lorenzo-Jiménez, Yasmin1
 http://orcid.org/0009-0004-5930-1753
http://orcid.org/0009-0004-5930-1753
Gutiérrez-Estrada, Gabriela23
http://orcid.org/0009-0004-4910-1520
Carricart-Ganivet, Juan P2
http://orcid.org/0000-0001-7266-8905
Tortolero-Langarica, JJ Adolfo12*
http://orcid.org/0000-0001-8857-5789
http://orcid.org/0009-0004-5930-1753Gutiérrez-Estrada, Gabriela23
http://orcid.org/0009-0004-4910-1520Carricart-Ganivet, Juan P2
http://orcid.org/0000-0001-7266-8905Tortolero-Langarica, JJ Adolfo12*
http://orcid.org/0000-0001-8857-5789
-
Publication dates-
May 20, 2025
Jan-Dec , 2024
- Article in PDF
- Article in XML
- Automatic translation
- Send this article by e-mail
- Share this article +
Abstract.
Massive corals of the genus Orbicella are key organisms that help maintain the physical structure of Caribbean reefs. However, they are currently threatened by environmental changes, such as increased nutrient loads and pollution, which affect the optical properties of seawater and, consequently, limit reef development. Thus, analyzing the responses in the growth of coral species to changes in light environments can help us improve mitigation and conservation strategies for coral reefs. The objective of this study was to evaluate the effect of changes in environmental light conditions on the growth rate of Orbicella faveolata by comparing fragments transplanted from 9 m to 3 m depth and control fragments that were transplanted under the same light conditions (3 m). The fragments of both treatments showed similar growth (16-23%), as well as comparable extension and diameter values. The annual growth rate for the control fragments and transplantation treatment fragments was 1.04 ± 0.18 cm·y-1 and 1.11 ± 0.23 cm·y-1, respectively. The results of this study reveal that the coral O. faveolata can physiologically acclimate to new environmental light conditions after being transplanted from a deep environment to a shallow environment in the short-term (1-9 months). This suggests great potential for the use of O. faveolata in restoration strategies and management programs that aim to maintain the populations and structural framework of coral reefs in the Caribbean region.
Key words::
coral fragments, coral reefs, coral transplantation, massive corals, skeletal growth
INTRODUCTION
Coral reefs are among the most complex and diverse ecosystems on the planet. They are characterized as sites for feeding, reproduction, and shelter and host approximately 25% of all marine species (Sheppard et al. 2009, Veron 2010). Coral reefs are marine benthic formations formed by the accumulation of biogenic calcium carbonate (CaCO3) secreted by calcifying organisms, such as hermatypic corals, mollusks, and coralline algae (Sheppard et al. 2009). As primary producers, hermatypic corals play a key role in forming these physical structures; under optimal conditions, they can generate up to 10 kg of CaCO3·m3·y-1 (Chave et al. 1972, Kleypas et al. 1999). In addition to increasing the three-dimensionality and complexity of habitats, hermatypic corals are fundamental to the balance of various biogeochemical cycles and ecosystem services (Perry and Alvarez-Filip 2018).
-
Sheppard et al.
2009The Biology of Coral Reefs, 2009
-
Veron 2010A Reef in Time, 2010
-
Sheppard et al.
2009The Biology of Coral Reefs, 2009
-
Chave et al. 1972Carbonate production by coral reefsMar Geol, 1972
-
Kleypas et al.
1999Geochemical consequences of increased atmospheric carbon dioxide on coral reefsScience, 1999
-
Perry
and Alvarez-Filip 2018Changing geo-ecological functions of coral reefs in the AnthropoceneFunct Ecol, 2018
Hermatypic corals have the ability to form exoskeletons by precipitating CaCO3 in the form of aragonite crystals. This process is controlled by intrinsic factors, such as genotypic variation, symbiotic community, morphology, and coral sex, as well as extrinsic factors, such as temperature, seawater chemistry, and light environment (Foster et al. 1979, Todd 2008, Allemand et al. 2011). Coral growth rates depend on carbonate calcification, which requires energy resources derived from photosynthesis, which are translocated by symbionts (Colombo-Pallota et al. 2010).
-
Foster et al.
1979Phenotypic plasticity in the reef corals Montastraea annularis (Ellis & Solander) and Siderastrea siderea (Ellis & Solander)J Exp Mar Biol Ecol, 1979
-
Todd 2008Morphological plasticity in scleractinian coralsBiolo Rev, 2008
-
Allemand et al. 2011Coral calcification, cells to reefsCoral reefs: an ecosystem in transition, 2011
-
Colombo-Pallota et al. 2010Calcification in bleached and unbleached Montastraea faveolata: evaluating the role of oxygen and glycerolCoral Reefs, 2010
Both the symbiont and the host show outstanding phenotypic plasticity that allows them to develop in different light and hydrodynamic environments (Kaniewska et al. 2011). The skeletal architecture built by each polyp plays an important role in the internal light environment; optical (reflective) properties of the skeleton cause multiple light scattering, favoring photosynthesis in symbiont algae (Enríquez et al. 2017). Therefore, the role of the symbiotic community in the translocation of energy to the coral for the production of CaCO3 and construction of its skeleton is highly relevant (Enríquez et al. 2005, 2017; Colombo-Pallotta et al. 2010, Scheufen et al. 2017). In this sense, the absorption and reflection properties of hermatypic coral skeletons depend on the structural arrangement that makes up their internal architecture, which modulates the dispersion of light, influencing the amount of energy available for the metabolic processes of the host (Enríquez et al. 2017). Therefore, depending on light conditions, corals can have different morphologies at the corallite level and at the colony level (Graus and Macintyre 1976, 1982; Klaus et al. 2007). In conditions of low light availability (>20 m), corals generally show plating morphologies to capture the greatest amount of light, whereas in conditions of greater light availability (1-5 m), corals have more complex morphologies (e.g., columnar or hemispherical), and colonies use self-shade to avoid excess light (Kramer et al. 2021).
-
Kaniewska et al. 2011Importance of macro-versus microstructure in modulating light levels inside coral coloniesJ Phycol, 2011
-
Enríquez et al. 2017Key functional role of the optical properties of coral skeletons in coral ecology and evolutionProc R Soc B., 2017
-
Enríquez et al. 2005Multiple scattering on coral skeletons enhances light absorption by symbiotic algaeLimnol Oceanogr, 2005
-
2017Key functional role of the optical properties of coral skeletons in coral ecology and evolutionProc R Soc B., 2017
-
Colombo-Pallotta et al.
2010Calcification in bleached and unbleached Montastraea faveolata: evaluating the role of oxygen and glycerolCoral Reefs, 2010
-
Scheufen et al. 2017Changes in the number of symbionts and Symbiodinium cell pigmentation modulate differentially coral light absorption and photosynthetic performanceFront Mar Sci, 2017
-
Enríquez et
al. 2017Key functional role of the optical properties of coral skeletons in coral ecology and evolutionProc R Soc B., 2017
-
Graus and Macintyre 1976Light control of growth form in colonial reef corals: a computer simulationScience, 1976
-
1982Variation in forms of the Reef Coral Montastraea annularis (Ellis and Solander): A quantitative evaluation of growth response to light distribution using computer simulationThe Atlantic Barrier Ecosystem at Carrie Bow Cay, Belize I. Structure and Communities, 1982
-
Klaus et al.
2007Environmental controls on corallite morphology in the reef coral Montastraea annularisBull Mar Sci, 2007
-
Kramer et al. 2021Characterization of morpho-functional traits in mesophotic corals reveals optimized light capture and photosynthesisbioRxiv, 2021
Orbicella is one of the most important hermatypic coral genera in the Caribbean region and one of the most threatened by environmental changes, such as pollution or increased nutrients in seawater due to wastewater discharge (Rico-Ensenaro et al. 2023). Species of the genus Orbicella obtain energy from heterotrophic assimilation (15-35%) by feeding on zooplankton and from autotrophic assimilation (70-90%) through the translocation of nutrients from endosymbiotic algae (Houlbrèque and Ferrier-Pagès 2009, Yranzo et al. 2020). Symbiont photosynthetic products contribute the most to the energy balance needed for the survival and growth of Orbicella colonies (Teece et al. 2011). However, faced with changes in seawater quality (e.g., changes in optical or chemical properties), it is important to understand the effect that light environments have on the growth of corals and the phenotypic capacity of corals to acclimate to different environmental gradients (Merks et al. 2004).
-
Rico-Ensenaro et al. 2023The δ15N in Orbicella faveolata organic matter reveals anthropogenic impact by sewage inputs in a Mexican Caribbean coral reef lagoonEnviron Sci Pollut Res, 2023
-
Houlbrèque and Ferrier-Pagès 2009Heterotrophy in tropical scleractinian coralsBiol Rev, 2009
-
Yranzo et al. 2020Coral pilar estrella y coral estrella montañoso Orbicella annularis y Orbicella faveolata, Venezuela, 2020
-
Teece
et al. 2011Heterotrophic y autotrophic assimilation of fatty acids by two scleractinian corals, Montastraea faveolata and Porites astreoidesLimnol Oceanogr, 2011
-
Merks et al. 2004Polyp oriented modelling of coral growthJ Theor Biol, 2004
Despite the important effect that light has on the morphology and phenotypic plasticity of corals, the information on the growth mechanisms associated with changes in light conditions is scarce (Todd 2008). In particular, how changes in light conditions affect coral growth has scarcely been explored. Orbicella faveolata is distributed over a wide depth gradient (1-50 m); thus, we believe it has great potential to acclimate physiologically to new light environments. Therefore, the objective of the present study was to evaluate the growth response of O. faveolata colonies transplanted between different light habitats (i.e., deep to shallow) and assess their acclimation capacity and potential use in restoration interventions.
-
Todd
2008Morphological plasticity in scleractinian coralsBiolo Rev, 2008
MATERIALS AND METHODS
Study area
The study was performed in the Jardines reef of the Arrecife de Puerto Morelos National Park (PNAPM, for its acronym in Spanish). This protected natural area was established in 1998 and is located in front of the town of Puerto Morelos in the state of Quintana Roo (21°00′00″ to 20°48′33″ N, 86°53′14.94″ to 86°46′38.94″ W; Fig. 1). The Puerto Morelos reef is a fringing coastal reef that forms an internal lagoon, with a depth of 3-5 m, characterized by calcareous sand stabilized by seagrass meadows (CONANP 2000). The area has a well-developed back reef and reef crest, a relatively flat fore reef with some massive coral sections, and several channels that gradually descend to 20-25 m toward an extensive sand platform (Rodríguez-Martínez et al. 2010). The reef zone is composed by 41 species of scleractinian corals and 32 gorgonian corals. The most abundant hermatypic coral species in the region are O. faveolata, Orbicella annularis, Acropora palmata, Acropora cervicorni, Millepora complanata, Pseudodiploria strigosa, Montastraea cavernosa, Dichocoenia stokesii, and Agaricia tenuifolia (Ruíz-Rentería et al. 1998, Alvarez-Filip et al. 2019, Caballero-Aragón 2020, Molina-Ramos 2020). However, in the last 5 years, live coral cover has decreased (>60%) due to the effects of thermal anomalies, hurricanes, and diseases that cause changes in species composition and dominance (Alvarez-Filip et al. 2022).
-
CONANP 2000Programa de Manejo del Parque Nacional Arrecife de Puerto Morelos, 2000
-
Rodríguez-Martínez et al. 2010Environmental state and tendencies of the Puerto Morelos CARICOMP site, MéxicoRev Biol Trop, 2010
-
Ruíz-Rentería et al. 1998Puerto Morelos, Quintana Roo, MéxicoCaribbean Coral Reef, Seagrass and Mangrove Sites, 1998
-
Alvarez-Filip et al. 2019A rapid spread of the stony coral tissue loss disease outbreak in the Mexican CaribbeanPeerJ Preprints, 2019
-
Caballero-Aragón 2020Puerto Morelos coral reefs, their current state and classification by a scoring systemDiversity, 2020
-
Molina-Ramos 2020Estado de conservación y cambios en el arrecife del Parque Nacional Arrecife de Puerto Morelos ante las amenazas antropogénicas [dissertation], 2020
-
Alvarez-Filip et al. 2022Stony coral tissue loss disease decimated Caribbean coral populations and reshaped reef functionalityCommun Biol, 2022
Thumbnail
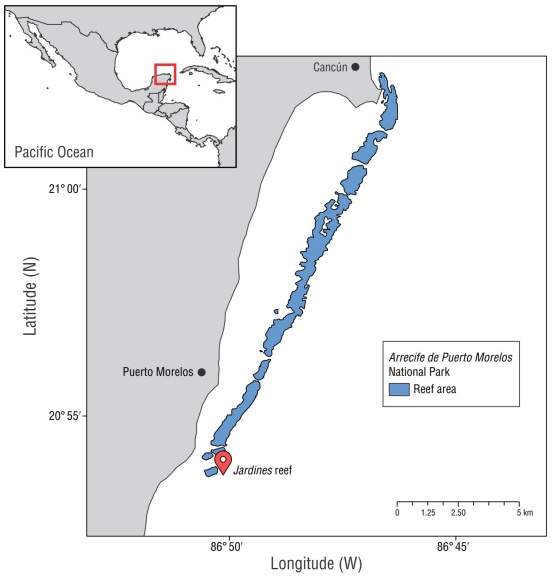
Figure 1
Map of Arrecife de Puerto Morelos National Park (APMNP), Mexico. The marker indicates the study site.
Map of Arrecife de Puerto Morelos National Park (APMNP), Mexico. The marker indicates the study site.
Obtaining growth parameters
For the transplantation treatment, 16 fragments of O. faveolata were obtained from 4 adult donor colonies of similar size (hemispherical, 40-cm height) that appeared healthy and were in the same light gradient (9 m). For the control treatment, the same procedure was done at a 3-m depth light gradient. All fragments (3 cm2) were extracted using a submersible hydraulic electric drill (Nemo Power Tools, Hong Kong, China). The fragments obtained at 9 m (transplantation treatment) were transplanted to the depth of the control fragments (3 m); all fragments were installed on 2 fiberglass beds. Each fragment was fixed to an acrylic plate using epoxy plasticine. After installation, the fragments were allowed to acclimate for 2 months to reduce the effect of handling stress (Fig. 2).
Thumbnail
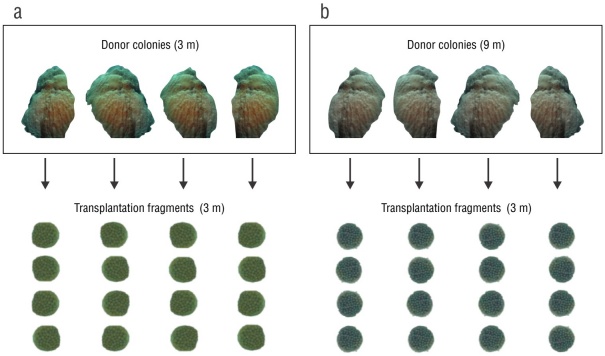
Figure 2
Experimental design. Donor coral fragments were collected at 2 depths: (a) 3 m (control treatment) and (b) 9 m (transplantation treatment). Four fragments were obtained from each donor colony, which were transplanted to a depth of 3 m.
Experimental design. Donor coral fragments were collected at 2 depths: (a) 3 m (control treatment) and (b) 9 m (transplantation treatment). Four fragments were obtained from each donor colony, which were transplanted to a depth of 3 m.
At the end of the acclimation period, during a period of 9 months, the following variables were measured monthly in each fragment: linear extension (cm), defined as the longitudinal distance from the base of the acrylic to the maximum height of the fragment, and the apical diameter (cm), defined as the maximum diameter from an aerial perspective. All measurements were estimated from digital images (25 MP) obtained with a GoPro Hero 9 camera (San Mateo, USA) (in linear function) using a plastic vernier (accuracy 0.05 cm) as a reference scale. The images from each monthly sampling were processed for each fragment for both treatments using the ImageJ program (Schneider et al. 2012).
-
Schneider
et al. 2012NIH Image to ImageJ: 25 years of image analysisNat Methods, 2012
This study considered the environmental variables seawater temperature (°C) and photosynthetically active radiation (PAR) to describe their influence on the growth of the massive coral O. faveolata. Daily values (May 2021 to February 2022) were obtained for both variables from the database of the Sistema Académico de Monitoreo Meteorológico y Oceanográfico (https://sammo.icmyl.unam.mx); these were averaged to obtain a monthly value for each variable.
Statistical analysis
Descriptive statistics were obtained for all monthly growth parameters (i.e., the average, standard deviation, ranges, and maximum and minimum values). The annual growth rate was obtained by extrapolating data from the 9 months of the study; the average value of each month was used to avoid overestimating the growth. A general monthly value was obtained, which was added to each of the 3 missing months to complete the annual period. After testing the normality and homoscedasticity of the data, various one-way analyses of variance (ANOVA) were performed, using generalized linear models (GLM), to evaluate the differences in growth parameters at the treatment and time level (monthly), including their interaction. Finally, simple linear regression models (r 2) were used to evaluate the relationship between growth characteristics and environmental factors (temperature and PAR). All statistical analyses were performed in the SigmaPlot program v. 11 (Graffiti LLC, Palo Alto, USA), using a 95% confidence interval (α = 0.05).
RESULTS
Linear extension
At the end of the experimental period, no signs of bleaching, loss, or death of tissue were observed in any of the fragments in the 2 treatments. Growth in linear extension showed a cumulative increase of 0.74 cm for the control treatment after a 9-month period, showing a monthly growth of 0.09 ± 0.02 cm (range: 0.04-0.26) (Table 1). The transplantation treatment showed a total accumulated growth of 0.70 cm, with a monthly average of 0.08 ± 0.01 cm (range: 0.06-0.64). The annual growth of the linear extension was 1.04 ± 0.18 cm·y-1 and 1.11 ± 0.23 cm·y-1 for the control treatment and the transplanted fragments, respectively. The results obtained for the linear extension showed no significant differences between treatments (F 1,69 = 0.050, P > 0.823), over time (months; F 6,69 = 2.154, P > 0.061), or in interactions between both variables (F 6,69 = 0.745, P > 0.616) (Fig. 3). The relative growth of the linear extension showed a similar increase (control: 16%; transplant: 15%) (Table 2).
Table 1
Growth variables (cm·month-1) measured in Orbicella faveolata fragments for the transplantation and control treatments at 3-m depth and environmental variables measured in the Arrecife de Puerto Morelos National Park (PNANP, for its acronym in Spanish).
Growth variables (cm·month-1) measured in Orbicella faveolata fragments for the transplantation and control treatments at 3-m depth and environmental variables measured in the Arrecife de Puerto Morelos National Park (PNANP, for its acronym in Spanish).
| Trasplantation | Control | |||||
| Month | Linear extension (cm) | Apical diameter (cm) | Linear extension (cm) | Apical diameter (cm) | Temperature (°C) | PAR (µmol·quanta·m-2·d-1) |
| June | 0.64 ± 0.67 | 0.15 ± 0.15 | 0.04 ± 0.05 | 0.06 ± 0.11 | 29.61 ± 1.09 | 52,539 ± 39,084 |
| August | 0.11 ± 0.09 | 0.18 ± 0.16 | 0.13 ± 0.10 | 0.12 ± 0.17 | 30.56 ± 1.10 | 60,112 ± 40,072 |
| September | 0.06 ± 0.07 | 0.09 ± 0.07 | 0.07 ± 0.05 | 0.17 ± 0.18 | 30.09 ± 0.97 | 54,275 ± 38,510 |
| October | 0.10 ± 0.07 | 0.17 ± 0.08 | 0.08 ± 0.07 | 0.14 ± 0.12 | 29.94 ± 0.88 | 54,513 ± 34,755 |
| November | 0.13 ± 0.14 | 0.09 ± 0.04 | 0.26 ± 0.24 | 0.14 ± 0.11 | 27.21 ± 0.87 | 42,145 ± 28,672 |
| January | 0.08 ± 0.10 | 0.10 ± 0.06 | 0.04 ± 0.01 | 0.16 ± 0.21 | 25.95 ± 1.15 | 42,612 ± 30,077 |
| February | 0.13 ± 0.09 | 0.22 ± 0.19 | 0.09 ± 0.06 | 0.15 ± 0.21 | 26.48 ± 1.10 | 52,144 ± 34,325 |
Table 2
Increase in monthly growth percentage (%) for the linear extension and apical diameter variables in Orbicella faveolata fragments for the control and transplantation treatments at 3-m depth.
Increase in monthly growth percentage (%) for the linear extension and apical diameter variables in Orbicella faveolata fragments for the control and transplantation treatments at 3-m depth.
| Month | Trasplantation | Control | ||
| Linear extension (cm) | Apical diameter (cm) | Linear extension (cm) | Apical diameter (cm) | |
| June | 1.37 ± 1.34 | 3.92 ± 3.92 | 1.38 ± 1.88 | 1.67 ± 2.84 |
| August | 2.44 ± 2.44 | 4.78 ± 4.70 | 3.50 ± 3.70 | 3.05 ± 4.36 |
| September | 1.16 ± 1.32 | 2.03 ± 1.82 | 1.78 ± 1.56 | 4.11 ± 4.33 |
| October | 2.34 ± 2.05 | 3.86 ± 1.90 | 2.08 ± 2.23 | 3.55 ± 3.62 |
| November | 2.54 ± 2.69 | 1.96 ± 0.99 | 4.70 ± 4.36 | 3.27 ± 2.72 |
| January | 2.08 ± 3.14 | 2.31 ± 1.60 | 0.88 ± 0.21 | 3.70 ± 5.07 |
| February | 3.07 ± 2.70 | 4.83 ± 4.38 | 1.81 ± 1.31 | 3.15 ± 4.23 |
Thumbnail
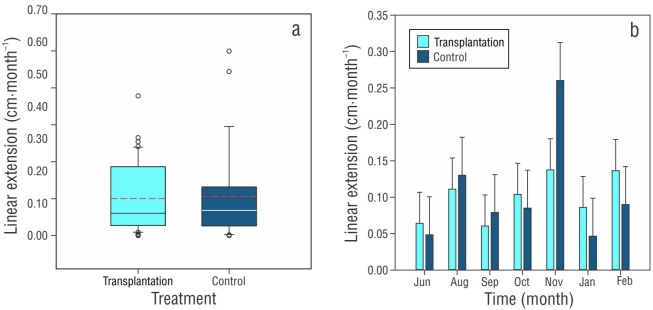
Figure 3
Boxplots of the average monthly growth of the linear extension (± SD) of Orbicella faveolata at 3-m depth over 9 months by (a) treatment (the black line represents the median, and the dashed red line denotes the mean) and (b) over time (months).
Boxplots of the average monthly growth of the linear extension (± SD) of Orbicella faveolata at 3-m depth over 9 months by (a) treatment (the black line represents the median, and the dashed red line denotes the mean) and (b) over time (months).
Apical diameter
The total cumulative apical diameter growth for the control treatment after the study period was 0.96 cm, with a monthly average of 0.12 ± 0.02 cm (range: 0.06-0.17) (Table 1). The total cumulative growth of the transplantation treatment was 1.03 cm, with a monthly average of 0.13 ± 0.03 cm (range: 0.09-0.22). The annual growth of the apical diameter was 1.45 ± 0.67 cm·y-1 for the control treatment and 1.54 ± 0.40 cm·y-1 for the transplantation treatment.
The results of the apical diameter showed no significant differences between treatments (F 1,69 = 0.0641, P > 0.801), over time (months; F 6,69 = 0.365, P > 0.898), or in the interaction between variables (F 6,69 = 0.599, P > 0.730) (Fig. 4). Apical diameter growth increased 23% for the control treatment and 24% for the transplantation treatment (Table 2).
Thumbnail
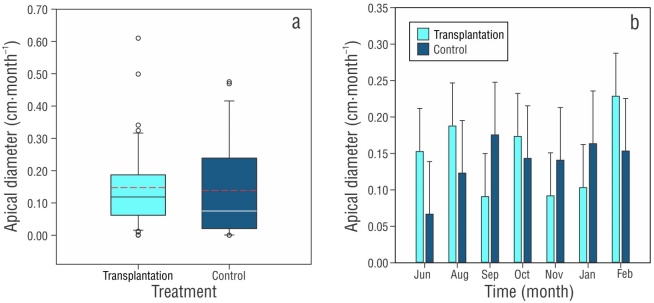
Figure 4
Boxplots of the average monthly growth of the apical diameter (± SD) of Orbicella faveolata at 3-m depth over 9 months by (a) treatment (the black line represents the median, and the dashed red line represents the mean) and (b) over time (months).
Boxplots of the average monthly growth of the apical diameter (± SD) of Orbicella faveolata at 3-m depth over 9 months by (a) treatment (the black line represents the median, and the dashed red line represents the mean) and (b) over time (months).
Environmental variables
The monthly average of the sea surface temperature for PNAPM during the study period was 28.62 °C (25.95-30.56 °C) (Table 1), with the highest values during the months of August and September (~30.32 °C) and the lowest values in January and February (~26.21 °C). Irradiance remained at an average of 52,286 µmol·quanta·m-2·d-1, with the lowest values in November (42,145 µmol· quanta·m-2·d-1) and maximum values in August (60,112 µmol· quanta·m-2·d-1) (Table 1). August showed the highest values for temperature and light incidence; however, the results of the linear regression models showed no deterministic relationship (r 2) between the environmental variables, or with the linear extension (P > 0.05), or with the apical diameter (P > 0.05).
DISCUSSION
In corals, skeletal growth is controlled by various factors, such as salinity, nutrients, exposure to waves, sedimentation, aragonite saturation state, temperature, and light; the last 2 factors have the most influence (Nybakken 2001, Sheppard et al. 2009, Veron 2010, Calderón-Aguilera et al. 2017). In the case of light, it has been proposed that the average calcification rate and extension rate decrease proportionally as irradiance decreases with increasing depth (Dustan 1975, Bosscher 1993). Conversely, in shallow waters, irradiance can reach excessive energy levels and cause photodamage due to oxidative stress (increase in reactive oxygen species) that reduces the photosynthetic capacity of the symbiont algae (Todd et al. 2004, Rodríguez-Troncoso et al. 2014).
-
Nybakken 2001Marine Biology: an ecological approach, 2001
-
Sheppard et al. 2009The Biology of Coral Reefs, 2009
-
Veron 2010A Reef in Time, 2010
-
Calderón-Aguilera et al. 2017Los arrecifes coralinos de México: Servicios ambientales y secuestro de carbonoElem Polít Públ, 2017
-
Dustan 1975Growth and form in the reef-building coral Montastrea annularisMar Biol, 1975
-
Bosscher 1993Computarized tomography and skeletal density of coral skeletonsCoral Reefs, 1993
-
Todd et al. 2004Genotype x environment interactions in transplanted clones of the massive corals Favia speciosa and Diploastrea helioporaMar Ecol Prog Ser, 2004
-
Rodríguez-Troncoso et al. 2014Corales: organismos base constructores de los ecosistemas arrecifalesTemas sobre Investigaciones Costeras, 2014
Previous studies focused on the relationship between coral morphology and light show that colonies modify their morphology to flatter shapes with increasing depth to increase their ability to capture light (Graus and Macintyre 1982, Willis 1987, Gutiérrez-Estrada 2017, Prada et al. 2022). Ow and Todd (2010) suggested that these morphological changes provide a long-term solution that requires less energy investment compared to the energy required for physiological adjustments.
-
Graus and
Macintyre 1982Variation in forms of the Reef Coral Montastraea annularis (Ellis and Solander): A quantitative evaluation of growth response to light distribution using computer simulationThe Atlantic Barrier Ecosystem at Carrie Bow Cay, Belize I. Structure and Communities, 1982
-
Willis 1987Phenotypic plasticity versus phenotypic stability in the reef corals Turbinaria mesenterina and Pavona cactus. In: Morphological Variation in the reef corals Turbinaria esenterina and Pavona cactus: synthesis of transplant, histocompatibility, electrophoresis, growth, and reproduction studies [dissertation], 1987
-
Gutiérrez-Estrada 2017Relación entre las características de crecimiento y los disepimentos en Orbicella faveolata creciendo en un gradiente lumínico [dissertation], 2017
-
Prada et al. 2022Linking photoacclimation responses and microbiome shifts between depth-segregated sibling species of reef coralsR Soc Open Sci, 2022
-
Ow and Todd
(2010)Light-induced morphological plasticity in the scleractinian coral Goniastrea pectinata and its functional significanceCoral Reefs, 2010
The effect of light intensity on the characteristics and morphology of skeletal growth in hermatypic corals has been studied previously (Graus and Macintyre 1982, Hubbard and Scature 1985, Carricart-Ganivet et al. 2007, Todd 2008, Gutiérrez-Estrada 2017, Mallon et al. 2022). However, the effect of coral transplantation between depths with different environmental light conditions has been unexplored. This study shows a rapid response in the skeletal growth of the coral O. faveolata and its short-term physiological plasticity (1-9 months), which could be a mechanism to compensate the effects of transplantation between sites with different light gradients. Therefore, the coral O. faveolata shows physiological plasticity in the short-term that possibly entails morphotypic modification in the long-term (>1 year), which allows it to acclimate to new light environments.
-
Graus and Macintyre 1982Variation in forms of the Reef Coral Montastraea annularis (Ellis and Solander): A quantitative evaluation of growth response to light distribution using computer simulationThe Atlantic Barrier Ecosystem at Carrie Bow Cay, Belize I. Structure and Communities, 1982
-
Hubbard and Scature 1985Growth rates of seven species of scleractinean corals from Cane Bay and Salt River, St. Croix, USVIBull Mar Sci, 1985
-
Carricart-Ganivet et al. 2007Growth and luminescence characteristics in skeletons of massive Porites from a depth gradient in the central Great Barrier ReefJ Exp Mar Biol Ecol, 2007
-
Todd
2008Morphological plasticity in scleractinian coralsBiolo Rev, 2008
-
Gutiérrez-Estrada 2017Relación entre las características de crecimiento y los disepimentos en Orbicella faveolata creciendo en un gradiente lumínico [dissertation], 2017
-
Mallon et al. 2022Light-driven dynamics between calcification and production in functionally diverse coral reef calcifiersLimnol Oceanogr, 2022
Reports indicate that, in the Caribbean Sea, corals of the genus Orbicella are physiologically capable of living in shallow and deep habitats for at least 6 months after being transplanted (transplantation from shallow habitat to deep habitat and from deep habitat to shallow habitat) (Prada et al. 2022). This finding coincides with the results obtained in this study; O. faveolata showed physiological plasticity (in terms of growth) in response to changes in light and depth conditions. This could show that corals of the genus Orbicella have adapted to different light environments through an energy balance via autotrophy and heterotrophy in relation to light availability (Prada et al. 2022).
-
Prada et al. 2022Linking photoacclimation responses and microbiome shifts between depth-segregated sibling species of reef coralsR Soc Open Sci, 2022
-
Prada et al. 2022Linking photoacclimation responses and microbiome shifts between depth-segregated sibling species of reef coralsR Soc Open Sci, 2022
Our results demonstrated that the transplanted colonies quickly reached the growth rate of the control colonies and showed similar monthly growth during the 9-month period of the study. At 3 m depth, the annual linear extension for the control treatment and the transplantation treatment was 1.04 cm·y-1 and 1.11 cm·y-1, respectively. These results contrast with the extension rate (0.80 cm·y-1) reported for O. faveolata at 9 m depth in the Mexican Caribbean (Gutiérrez-Estrada 2017). A similar pattern has been reported in Florida (USA), where the linear extension of colonies in the shallow zone was higher than that of colonies in the deep zone (>6 m) during the same time period (Manzello et al. 2015). Gutiérrez-Estrada (2017) suggested that this could be due to the fact that skeletal extension depends on the number of exothecal dissepiments deposited per year, whereas density depends on their thickness, which are directly influenced by blue (lunar) light and conditions detected by corals at relatively shallow depths.
-
Gutiérrez-Estrada
2017Relación entre las características de crecimiento y los disepimentos en Orbicella faveolata creciendo en un gradiente lumínico [dissertation], 2017
-
Manzello et al. 2015Recent decade of growth and calcification of Orbicella faveolata in the Florida Keys: an inshore-offshore comparisonMar Ecol Prog Ser, 2015
-
Gutiérrez-Estrada
(2017)Relación entre las características de crecimiento y los disepimentos en Orbicella faveolata creciendo en un gradiente lumínico [dissertation], 2017
In light environments that facilitate optimal growth, massive colonies of Porites have been observed to exhibit higher growth in linear extension in contrast to colonies located in environments with high turbidity (Lough et al. 1999). Furthermore, reports indicate that colonies transplanted to shallower environments show increased lateral extension (diameter), which coincides with the results of the present study (Table 1). This could be explained by the size of the coral fragments (3-3.5 cm²), because, when a sexually mature colony fragments, the coral resources are allocated to growth and calcification processes instead of gametogenesis (Forsman et al. 2015). In this context, the transplant site and genotype of origin can influence calcification rates, resulting in an interaction effect between environmental and parental factors on the skeletal characteristics of massive corals (Smith et al. 2007). This could suggest that colony morphology varies depending on habitat, which results in high phenotypic plasticity in the skeletal characteristics of corals (Smith et al. 2007).
-
Lough et al. 1999Variability in growth characteristics of massive Porites on the Great Barrier ReefCRC Reef Res Cen Tech Rep, 1999
-
Forsman et al.
2015Growing coral larger and faster: micro-colony-fusion as a strategy for accelerating coral coverPeerJ, 2015
-
Smith et al. 2007Phenotypic plasticity for skeletal growth, density and calcification of Porites lobata in response to habitat typeCoral Reefs, 2007
-
Smith et al. 2007Phenotypic plasticity for skeletal growth, density and calcification of Porites lobata in response to habitat typeCoral Reefs, 2007
Previous reports with similar methodologies have found that coral species, such as Turbinaria mesenterina, found at depths of 1 m (shallow) and 4 m (deep) were able to adapt to the environmental conditions of both depths after transplantation; nevertheless, their growth was lower in the deep zone (Willis 1987). Other species of massive corals have shown similar patterns when transplanted to different environments. For example, Ow and Todd (2010) found higher growth in Goniastrea pectinata colonies transplanted to a depth of 7 m in contrast to the growth of colonies transplanted to a depth of 3 m after a short period (163 days). This could be explained by the high phenotypic plasticity and photo adaptive response to light irradiance of some genera of massive corals, such as Porites and Orbicella, which allow them to distribute over a depth light gradient (Smith et al. 2007, Todd 2008).
-
Willis 1987Phenotypic plasticity versus phenotypic stability in the reef corals Turbinaria mesenterina and Pavona cactus. In: Morphological Variation in the reef corals Turbinaria esenterina and Pavona cactus: synthesis of transplant, histocompatibility, electrophoresis, growth, and reproduction studies [dissertation], 1987
-
Ow and Todd (2010)Light-induced morphological plasticity in the scleractinian coral Goniastrea pectinata and its functional significanceCoral Reefs, 2010
-
Smith et al.
2007Phenotypic plasticity for skeletal growth, density and calcification of Porites lobata in response to habitat typeCoral Reefs, 2007
-
Todd 2008Morphological plasticity in scleractinian coralsBiolo Rev, 2008
CONCLUSIONS
The results indicate that there were no significant differences in the linear extension or apical diameter between treatments and time periods (months). Therefore, the coral O. faveolata has the potential to acclimate to new environmental light conditions after being transplanted from a deep environment to a shallow environment in a short period of time. Furthermore, we suggest that transplanted colonies can achieve growth rates similar to those of control colonies relatively fast (1-9 months), which demonstrates phenotypic plasticity to changes in their light environment. Focusing on the lateral extension of the colonies, we observed a higher increase in diameter compared to the linear extension in both treatments; therefore, we can conclude that the colonies of both treatments used the available resources to repair the periphery damage caused by fragmentation.
However, the study period was relatively short to determine if the short-term response will be beneficial in the long term or if the survival and health of the colony will be compromised in the future. Therefore, we suggest conducting similar studies that consider long-term time periods (>9 months). The information from this study reveals that O. faveolata colonies transplanted from a deep to a shallow environment have the potential to acclimate to new light conditions in a short period of time.
Acknowledgments
We thank the Mexican authorities of the Arrecife de Puerto Morelos National Park (SEMARNAT/CONANP) for providing the collecting permit (log number 23/LW-0103/04/21) and facilitations granted. We also thank Miguel I. Gómez Reali, Edgar Escalante Mancera, and Fernando Negrete Soto for their assistance during field work.
REFERENCES
- Allemand D, Tambutte E, Zoccola D, Tambutte S. 2011. Coral calcification, cells to reefs. In: Dubinsky Z, Stambler N (eds.), Coral reefs: an ecosystem in transition. New York (USA): Springer. p. 119-150. Links
- Alvarez-Filip L, Estrada-Saldívar N, Pérez-Cervantes E, Molina-Hernández A, González-Barrios FJ. 2019. A rapid spread of the stony coral tissue loss disease outbreak in the Mexican Caribbean. PeerJ Preprints. 7:e27893v1. https://doi.org/10.7287/peerj.preprints.27893v1 Links
- Alvarez-Filip L, González-Barrios FJ, Pérez-Cervantes E, Molina-Hernández AM, Estrada-Saldívar N. 2022. Stony coral tissue loss disease decimated Caribbean coral populations and reshaped reef functionality. Commun Biol. 5:440. https://doi.org/10.1038/s42003-022-03398-6 Links
- Bosscher H. 1993. Computarized tomography and skeletal density of coral skeletons. Coral Reefs. 12:97-103. https://doi.org/10.1007/BF00302109 Links
- Caballero-Aragón H, Perera-Valderrama S, Cerdeira-Estrada S, Martell-Dubois R, Rosique-de la Cruz L, Álvarez-Filip L, Pérez-Cervantes E, Estrada-Saldívar N, Ressl R. 2020. Puerto Morelos coral reefs, their current state and classification by a scoring system. Diversity. 12(7):272. https://doi.org/10.3390/d12070272 Links
- Calderón-Aguilera LE, Reyes-Bonilla H, Norzagaray-López CO, López-Pérez RA. 2017. Los arrecifes coralinos de México: Servicios ambientales y secuestro de carbono. Elem Polít Públ. 1:53-62. Links
- Carricart-Ganivet JP, Lough JM, Barnes DJ. 2007. Growth and luminescence characteristics in skeletons of massive Porites from a depth gradient in the central Great Barrier Reef. J Exp Mar Biol Ecol. 351(1-2):27-36. https://doi.org/10.1016/j.jembe.2007.05.038 Links
- Chave KE, Smith SV, Roy KJ. 1972. Carbonate production by coral reefs. Mar Geol. 12(2):123-140. https://doi.org/10.1016/0025-3227(72)90024-2 Links
- Colombo-Pallotta MF, Rodríguez-Román A, Iglesias-Prieto R. 2010. Calcification in bleached and unbleached Montastraea faveolata: evaluating the role of oxygen and glycerol. Coral Reefs. 29:899-907. https://doi.org/10.1007/s00338-010-0638-x Links
- [CONANP] Comisión Nacional de Áreas Naturales Protegidas. 2000. Programa de Manejo del Parque Nacional Arrecife de Puerto Morelos. Puerto Morelos (Mexico): CONANP. Management program. 225 p. Links
- Dustan P. 1975. Growth and form in the reef-building coral Montastrea annularis. Mar Biol. 33:101-107. http://dx.doi.org/10.1007/BF00390714 Links
- Enríquez S, Méndez ER, Iglesias-Prieto R. 2005. Multiple scattering on coral skeletons enhances light absorption by symbiotic algae. Limnol Oceanogr. 50(4):1025-1032. https://doi.org/10.4319/lo.2005.50.4.1025 Links
- Enríquez S, Méndez ER, Hoegh-Guldberg O, Iglesias-Prieto R. 2017. Key functional role of the optical properties of coral skeletons in coral ecology and evolution. Proc R Soc B. 284:1-9. https://doi.org/10.1098/rspb.2016.1667 Links
- Fitt WK, McFarland FK, Warner ME, Chilcoat GC. 2000. Seasonal patterns of tissue biomass and densities of symbiotic dinoflagellates in reef corals and relation to coral bleaching. Limnol Oceanogr. 45(3):677-685. https://doi.org/10.4319/lo.2000.45.3.0677 Links
- Foster AB. 1979. Phenotypic plasticity in the reef corals Montastraea annularis (Ellis & Solander) and Siderastrea siderea (Ellis & Solander). J Exp Mar Biol Ecol. 39(1):25-54. https://doi.org/10.1016/0022-0981(79)90003-0 Links
- Forsman ZH, Page CA, Toonen RJ, Vaughan D. 2015. Growing coral larger and faster: micro-colony-fusion as a strategy for accelerating coral cover. PeerJ. 16:e1313. https://doi.org/10.7717/peerj.1313 Links
- Graus RR, Macintyre IG. 1976. Light control of growth form in colonial reef corals: a computer simulation. Science. 193(4256):895-897. https://doi.org/10.1126/science.193.4256.895 Links
- Graus RR, Macintyre IG. 1982. Variation in forms of the Reef Coral Montastraea annularis (Ellis and Solander): A quantitative evaluation of growth response to light distribution using computer simulation. In. Klaus R, Macintyre IG (eds.), The Atlantic Barrier Ecosystem at Carrie Bow Cay, Belize I. Structure and Communities. Washington (USA): Smithsonian Institution Press. p. 441-464. Links
- Gutiérrez-Estrada G. 2017. Relación entre las características de crecimiento y los disepimentos en Orbicella faveolata creciendo en un gradiente lumínico [dissertation]. [Mexico]: Universidad Nacional Autónoma de México. 39 p. Links
- Houlbrèque F, Ferrier‐Pagès C. 2009. Heterotrophy in tropical scleractinian corals. Biol Rev. 84(1):1-17 Links
- Kaniewska P, Magnusson SH, Anthony KRN, Reef R, Kühl M, Hoegh-Guldberg O. 2011. Importance of macro-versus microstructure in modulating light levels inside coral colonies. J Phycol. 47(4):846-860. https://doi.org/10.1111/j.1529-8817.2011.01021.x Links
- Kramer N, Guan J, Chen S, Wangpraseurt D, Loya Y. 2021. Characterization of morpho-functional traits in mesophotic corals reveals optimized light capture and photosynthesis. bioRxiv. 2021(09). https://doi.org/10.1101/2021.09.29.462347 Links
- Hubbard DK, Scaturo D. 1985. Growth rates of seven species of scleractinean corals from Cane Bay and Salt River, St. Croix, USVI. Bull Mar Sci. 36:325-338. Links
- Klaus J, Budd AF, Heikoop JM, Fouke BW. 2007. Environmental controls on corallite morphology in the reef coral Montastraea annularis. Bull Mar Sci. 80:233-260. Links
- Kleypas JA, Buddemeirer RW, Archer D, Gattuso JP, Langdon C, Opdyke BN. 1999. Geochemical consequences of increased atmospheric carbon dioxide on coral reefs. Science. 284(5411):118-120. https://doi.org/10.1126/science.284.5411.118 Links
- Lough JM, Barnes DJ, Devereux MJ, Tobin BJ, Tobin S. 1999. Variability in growth characteristics of massive Porites on the Great Barrier Reef. CRC Reef Res Cen Tech Rep. 28:95. Links
- Manzello DP, Enochs IC, Kolodziej G, Carlton R. 2015. Recent decade of growth and calcification of Orbicella faveolata in the Florida Keys: an inshore-offshore comparison. Mar Ecol Prog Ser. 521:81-89. https://doi.org/10.3354/meps11085 Links
- Mallon J, Cyronak T, Hall ER, Banaszak AT, Exton DA, Bass AM. 2022. Light-driven dynamics between calcification and production in functionally diverse coral reef calcifiers. Limnol Oceanogr. 67(2):434-449. https://doi.org/10.1002/lno.12002 Links
- Merks MH, Hoekstra AG, Kaandorp JA, Sloot PMA. 2004. Polyp oriented modelling of coral growth. J Theor Biol. 228(4):559-576. https://doi.org/10.1016/j.jtbi.2004.02.020 Links
- Molina-Ramos SI. 2020. Estado de conservación y cambios en el arrecife del Parque Nacional Arrecife de Puerto Morelos ante las amenazas antropogénicas [dissertation]. [Mexico]: Universidad Iberoamericana Puebla. 55 p. Links
- Nybakken JW. 2001. Marine Biology: an ecological approach. 5th ed. San Francisco (USA): Benjamin Cummings. 516 p. Links
- Ow YX, Todd PA. 2010. Light-induced morphological plasticity in the scleractinian coral Goniastrea pectinata and its functional significance. Coral Reefs. 29:797-808. https://doi.org/10.1007/s00338-010-0631-4 Links
- Page CA. 2013. Reskinning a reef: Mote marine lab scientists explore a new approach to reef restoration. Reef Mar Aqua Mag. 72-8. https://doi.org/10.13140/RG.2.1.4281.0967 Links
- Perry CT, Alvarez-Filip L. 2018. Changing geo-ecological functions of coral reefs in the Anthropocene. Funct Ecol. 33(6):976-988. https://doi.org/10.1111/1365-2435.13247 Links
- Prada C, López-Londoño T, Pollock FJ, Roitman S, Ritchie KB, Levitan DR, Knowlton N, Woodley C, Iglesias-Prieto R, Medina M. 2022. Linking photoacclimation responses and microbiome shifts between depth-segregated sibling species of reef corals. R Soc Open Sci .9:14. https://doi.org/10.1098/rsos.211591 Links
- Rico-Esenaro SD, Tortolero-Langarica JJA, Iglesias-Prieto R, Carricart-Ganivet JP. 2023. The δ15N in Orbicella faveolata organic matter reveals anthropogenic impact by sewage inputs in a Mexican Caribbean coral reef lagoon. Environ Sci Pollut Res. 30:118872-118880. https://doi.org/10.1007/s11356-023-30476-x Links
- Rodríguez-Martínez RE, Ruíz-Rentería F, Tussenbroek B, Barba-Santos G, Escalante-Mancera E, Jordán-Garza G, Jordán-Dahlgren E. 2010. Environmental state and tendencies of the Puerto Morelos CARICOMP site, México. Rev Biol Trop. 58(supp 3):23-43. Links
- Rodríguez-Troncoso AP, Tortolero-Langarica JJA. 2014. Corales: organismos base constructores de los ecosistemas arrecifales. In: Cifuentes-Lemus JL, Cupul-Magaña FG (eds.), Temas sobre Investigaciones Costeras. Guadalajara (Mexico): Universidad de Guadalajara. p. 33-55. Links
- Ruíz-Rentería F, Tussenbroek B, Jordán-Dahlgren E. 1998. Puerto Morelos, Quintana Roo, México. In: Björn Kjerfve (ed.), Caribbean Coral Reef, Seagrass and Mangrove Sites. Puerto Morelos (Mexico): UNESCO. p. 57-66. Links
- [SAMMO] Sistema Académico de Monitoreo Meteorológico y Oceanográfico. 2002. Datos Actuales Estación Puerto Morelos, Quintana Roo, México. Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México; [15 March 2022]. https://sammo.icmyl.unam.mx . Links
- Scheufen T, Iglesias-Prieto R, Enríquez S. 2017. Changes in the number of symbionts and Symbiodinium cell pigmentation modulate differentially coral light absorption and photosynthetic performance. Front Mar Sci. 4:1-16. https://doi.org/10.3389/fmars.2017.00309 Links
- Schneider C, Rasband W, Eliceiri K. 2012. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 9:671-675. https://doi.org/10.1038/nmeth.2089 Links
- Sheppard CRC, Davy SK, Pilling GM. 2009. The Biology of Coral Reefs. New York (USA): OXFORD University Press. 339 p. Links
- Smith LW, Barshis D, Birkeland C. 2007. Phenotypic plasticity for skeletal growth, density and calcification of Porites lobata in response to habitat type. Coral Reefs. 26:559-667. https://doi.org/10.1007/s00338-007-0216-z Links
- Teece MA, Estes B, Gelsleichter E, Lirman D. 2011. Heterotrophic y autotrophic assimilation of fatty acids by two scleractinian corals, Montastraea faveolata and Porites astreoides. Limnol Oceanogr. 56(4):1285-1296. https://doi.org/10.4319/lo.2011.56.4.1285 Links
- Todd PA, Ladle RJ, Lewin-Koh NIJ, Chou LM. 2004. Genotype x environment interactions in transplanted clones of the massive corals Favia speciosa and Diploastrea heliopora. Mar Ecol Prog Ser. 271:167-182. Links
- Todd PA. 2008. Morphological plasticity in scleractinian corals. Biolo Rev. 83(3):315-337. https://doi.org/10.1111/j.1469-185X.2008.00045.x Links
- Veron JEN. 2010. A Reef in Time. Australia: Harvard University Press. 304 p. Links
- Willis BL. 1987. Phenotypic plasticity versus phenotypic stability in the reef corals Turbinaria mesenterina and Pavona cactus. In: Morphological Variation in the reef corals Turbinaria esenterina and Pavona cactus: synthesis of transplant, histocompatibility, electrophoresis, growth, and reproduction studies [dissertation]. [Australia]: University of North Queensland. 53-87 p. Links
- Yranzo A, Villamizar E, Herrera-Reveles AT, Pérez J, Boadas H, Pereira C, Rodríguez JG, Narciso S, Bustillos F, Cavada-Blanco F. 2020. Coral pilar estrella y coral estrella montañoso Orbicella annularis y Orbicella faveolata, Venezuela. Venezuela: Instituto de Zoología y Ecología Tropical, EDGE of Existence, Zoological Society of London. Technical report. 38 p. https://doi.org/10.13140/RG.2.2.35996.31361/1 Links