Artículos de revisión
Conservation of natural enemies of Diaphorina citri and their impact on Huanglongbing: Analysis and perspectives

-
Publication dates-
February 24, 2025
May-Aug , 2024
- Article in PDF
- Article in XML
- Automatic translation
- Send this article by e-mail
- Share this article +
ABSTRACT
Diaphorina citri is the main vector of Candidatus Liberibacter asiaticus (CLas), the causative agent of HLB, the most devastating citrus disease. Although the importance of biological control is recognized, insecticides are the main control tool used. Its use in citrus should be cautious, as it could interfere with the biocontrol of exotic pests already present. Diaphorina citri has a wide range of natural enemies; however, only the parasitoid Tamarixia radiata and some species of entomopathogenic fungi are used inundatively. Although the main predators of the vector occur naturally, few studies address their conservation in situ. This review supports the idea that the conservation of natural enemies should be the basis of the integrated management of D. citri and CLas. The conservation of alternate hosts, the inclusion of nectar plants, in situ conservation of parasitoids, and the autodissemination of entomopathogenic fungi are proposed. Studies carried out on conservation of natural enemies of D. citri and related pests, their probable impact on the disease, and prospects for implementation in Mexico are analyzed and discussed. The proposed strategies could enhance not only the biological control of D. citri-CLas, but also the autoregulation of citrus pests in general.
Keywords:
Asian citrus psyllid, biocontrol, Tamarixia radiata, entomopathogenic fungi, conservation
Introduction
With over 600,000 hectares and a production exceeding 8 million tons in 2021, Mexico stands as one of the leading citrus producers. The economic contribution reached approximately 35 million pesos (SIAP, 2022), underscoring the significance of these crops within the country’s economic sector. Their production spans across 24 out of the 32 states comprising Mexico, from the Yucatan Peninsula to Tamaulipas along the Gulf of Mexico, and from Chiapas to Sonora and the Baja California Peninsula along the Pacific (Salcedo-Baca et al., 2010; García-Ávila et al., 2021).
-
SIAP, 2022Anuario estadístico de la producción agrícola, 2022
-
Salcedo-Baca et al., 2010Evaluación del Impacto Económico de la Enfermedad de los Cítricos Huanglongbing (HLB) en la Cadena Citrícola MexicanaInstituto Interamericano de Cooperación para la Agricultura (IICA), 2010
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
However, citrus crops face a plethora of phytosanitary challenges globally. Unlike other crops, successful cases of classical biological control occur within citrus cultivation. For instance, in Mexico, the cottony cushion scale (Icerya purchasi) is managed through the ladybird beetle Rodolia cardinalis and the citrus blackfly (Aleurocanthus woglumi), regulated by the parasitoid Encarsia perplexa, among others (Trevor et al., 2013).
This distinctive characteristic renders citrus cultivation unique. Unlike in other crops where chemical control forms the cornerstone of pest management, pesticides in citrus cultivation must be handled with greater caution. Their misuse could disrupt the achieved ecological balance and potentially lead to the resurgence of significant pests (Qureshi and Stansly, 2009), as mentioned above.
-
Qureshi and Stansly, 2009Incidence of invasive Diaphorina citri (Hemiptera: Psyllidae) and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrusJournal of Economic Entomology, 2009
In recent years, Mexico has witnessed new invasions of exotic pests, prominently the Asian citrus psyllid, Diaphorina citri (Hemiptera: Leviidae), which along with Trioza erytreae (Hemiptera: Triozidae), serves as a vector for Candidatus Liberibacter asiaticus (CLas), the causal agent of Huanglongbing (HLB). Following the psyllid’s introduction in 2002, HLB was first documented in 2009 in Yucatan. Since then, the disease has spread to various regions, encompassing 24 citrus- producing states (García-Ávila et al., 2021). The risks posed by the D. citri-CLas pathosystem in Mexican citriculture (Salcedo-Baca et al., 2010) prompted the development of integrated management strategies, highlighting chemical control alongside complementary biological approaches (García-Ávila et al., 2021).
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
-
Salcedo-Baca et al., 2010Evaluación del Impacto Económico de la Enfermedad de los Cítricos Huanglongbing (HLB) en la Cadena Citrícola MexicanaInstituto Interamericano de Cooperación para la Agricultura (IICA), 2010
In Mexico, the psyllid boasts a diverse array of natural enemies (NE), including one of its primary parasitoids, Tamarixia radiata (Hymenoptera: Eulophidae) (Cortez-Mondaca et al., 2010; Mora-Aguilera, 2013). Although widely distributed in Mexican citrus zones, levels of parasitism remain insufficient (3-26%) for the natural regulation of the pest and CLas (Halbert and Manjunath, 2004; García- Ávila et al., 2021). To enhance biological control (BC) of D. citri and mitigate CLas dispersal risks, the Mexican government has endeavored to establish a program of inundative biological control (IBC) utilizing the parasitoid T. radiata in citrus regions across the country (García-Ávila et al., 2021). Despite scientific literature stressing the importance of conserving NE in D. citri management, research in this realm remains scarce globally (Qureshi and Stansly, 2009; Irvin et al., 2021; Irvin and Hoddle, 2021), with documented studies in Mexico being nascent (Godoy-Ceja and Cortez-Madrigal, 2018).
-
Cortez-Mondaca et al., 2010Primer reporte de enemigos naturales y parasitismo sobre Diaphorina citri Kuwayama en Sinaloa, MéxicoSouthwestern Entomologist, 2010
-
Halbert and Manjunath, 2004Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in FloridaFlorida Entomologist, 2004
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
-
Qureshi and Stansly, 2009Incidence of invasive Diaphorina citri (Hemiptera: Psyllidae) and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrusJournal of Economic Entomology, 2009
-
Irvin et al., 2021Evaluating the potential of flowering plants for enhancing predatory hoverflies (Syrphidae) for biological control of Diaphorina citri (Liviidae) in CaliforniaBiological Control, 2021
-
Godoy-Ceja and Cortez-Madrigal, 2018Potencial de Asclepias curassavica L. (Apocynaceae) en el control biológico de plagasRevista Mexicana de Ciencias Agrícolas, 2018
Under the premise that integrated pest management (IPM) should rely on NE conservation, and insecticide application should occur only upon reaching an economic threshold (ET), this paper aims to analyze the status of D. citri-CLas pathosystem management globally, particularly in Mexico. Based on this, it proposes conservation strategies extending beyond D. citri biological control to encompass the regulation of other citrus phytophagous (including pests).
The citrus agroecosystem in Mexico
Cultivated citrus traces its origins back to Southeast Asia, where its evolutionary journey spans roughly 8 million years (Ibanez et al., 2015). Recognizing the geographical birthplace of a crop holds pivotal importance in combating exotic pests, often necessitating the exploration and importation of natural enemies from their native habitats. Over time, the citrus industry in Mexico has contended with a spectrum of exotic pests, some of which have been effectively managed through classical biological control methods (Trevor et al., 2013).
-
Ibanez et al., 2015El origen de las especies cultivadas de cítricosLevante Agrícola: Revista Internacional de Cítricos, 2015
In Mexico, citrus cultivation sprawls across 24 of the nation’s 32 states (García- Ávila et al., 2021). These regions boast diverse soil compositions, climatic variations, and cultural influences, all of which profoundly shape the design and management of the agroecosystem, including the dynamics of the D. citri-CLas pathosystem. Consequently, factors like wind patterns and orchard distribution have contributed to variations in the severity of Huanglongbing (HLB) outbreaks across different citrus regions, such as those along the Mexican Pacific versus the Yucatan Peninsula (Mora-Aguilera, 2013).
-
Mora-Aguilera, 2013Huanglongbing Candidatus Liberibacter spp. Ficha Ténica N° 78, 2013
Similar to their counterparts elsewhere, Mexican citrus growers exhibit distinct characteristics, necessitating consideration of their typology. This entails understanding their profiles based on factors like land holdings, technological adoption, and educational backgrounds (Rosales-Martínez et al., 2018). Such insights could facilitate tailored programs, including initiatives for conserving natural enemies. Consequently, the socioeconomic and cultural contexts of small- scale producers influence plantation features, ranging from mono-crops with weed tolerance (common in the Yucatan Peninsula) to intricate polycultures like the coffee-plantain-citrus system observed in certain regions of Veracruz (Aguilar et al., 2014). These agricultural models contrast starkly with extensive monocultures found in other parts of the country, such as Huimanguillo, Tabasco. Such variations extend to management practices, notably pesticide use, which is pivotal for implementing Conservation Biological Control (CBC) strategies (Gurr and You, 2016).
-
Rosales-Martínez et al., 2018Aspectos culturales, sociales y productivos para una tipología de cafeticultoresAgricultura, Sociedad y Desarrollo, 2018
-
Aguilar et al., 2014Evaluación socioeconómica del sistema agroforestal tradicional café-plátano-cítricos en el municipio de Tlapacoyan, Veracruz, MéxicoTropical and Subtropical Agroecosystems, 2014
-
Gurr and You, 2016Conservation biological control of pests in the molecular era: New opportunities to address old constraintsFrontiers in Plant Science, 2016
While some citrus plantations benefit from irrigation systems, the majority of Mexican citrus farming (63%) relies on rainfed agriculture (García-Ávila et al., 2021). Nonetheless, certain regions harbor conditions conducive to thriving populations of natural enemies. For instance, in Rovirosa, Quintana Roo, high relative humidity has fostered natural epizootics of the fungus Hirsutella citriformis in D. citri (Mora-Aguilera et al., 2017), underscoring the potential of entomopathogenic fungi (EPFs) as crucial tools for managing the D. citri-CLas complex in such areas.
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
The Citrus-Diaphorina citri-CLas pathosystem
In 2002, D. citri was introduced to Mexico and is now distributed across all citrus-growing areas of the country (García-Ávila et al., 2021). Apart from the direct damage it causes to citrus, D. citri serves as a vector for Gram-negative α-Proteobacteria of the genus Candidatus Liberibacter spp., the causal agent of HLB (Galdeano et al., 2020). Three species are involved: CL. africanus, CL. americanus, and CL. asiaticus, with the latter being the most prevalent worldwide (Thakuria et al., 2023). HLB damages and losses vary between countries, regions, and hosts. For example, in Brazil, losses in sweet orange fluctuated from 17.5-42%, while in Mexico, losses vary according to the crop and region, ranging from 17% for Persian lime in Yucatan to 62% for Mexican lime in Colima (Mora Aguilera et al., 2014). In a scenario of low HLB intensity in Mexico, losses of 1.84 million tons of citrus are estimated, representing 25% of the national production, with losses of 4 million workdays (Salcedo et al., 2010).
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
-
Galdeano et al., 2020Friend or foe? Relationship between ‘Candidatus Liberibacter asiaticus’ and Diaphorina citriTropical Plant Pathology, 2020
-
Thakuria et al., 2023Citrus Huanglongbing (HLB): Diagnostic and management optionsPhysiological and Molecular Plant Pathology, 2023
-
Mora Aguilera et al., 2014Situación actual y perspectivas del manejo del HLB de los cítricosRevista Mexicana de Fitopatología, 2014
The concept of an “epidemic” as an imbalance between populations can be applied to the citrus-D. citri-CLas pathosystem, where knowledge of its biology and ecology can make the difference between the success or failure of management programs. As a guide, the concept of an “epidemiological system” could be applied, conceived as an open system of n-factors operating in time and space to determine the incidence of an epidemic (Figure 1) (Mora-Aguilera et al., 2017).
For D. citri, the optimal temperature fluctuates between 25-28 °C, with development ceasing between 10 and 33°C (Liu and Tsai, 2000). Females cease oviposition when temperatures remain at 34 °C for five consecutive days; once temperatures decrease, adults resume oviposition (Skelley and Hoy, 2004). Conversely, CLas does not tolerate temperatures >35 °C (Thakuria et al., 2023). The number of eggs per D. citri female decreases when relative humidity remains below 40% (Skelley and Hoy, 2004).
-
Liu and Tsai, 2000Effects of temperature on biology and life table parameters of the Asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae)Annals of Applied Biology, 2000
-
Skelley and Hoy, 2004A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantineBiological Control, 2004
-
Thakuria et al., 2023Citrus Huanglongbing (HLB): Diagnostic and management optionsPhysiological and Molecular Plant Pathology, 2023
Moreover, humidity indirectly affects D. citri by favoring the prevalence of EPFs that reduce insect populations (Aubert, 1987). An interaction between light rainfall and high temperatures indirectly favored D. citri populations by promoting greater leaf flush in trees, a factor directly correlated with vector populations and HLB incidence (Leong et al., 2020). Wind is another key factor in D. citri and CLas dispersion. It has been documented that the psyllid can fly up to 2.4 km in the absence of wind. In the presence of wind, some estimates suggest the insect could disperse up to 4 km (Mora-Aguilera, 2013) or even 470 km (Stelinski, 2019).
-
Aubert, 1987Trioza erytreae Del Guercio and Diaphorina citri Kuwayama (Homoptera: Psylloidea), the two vectors of citrus greening disease: biological aspects and possible control strategiesFruits, 1987
-
Leong et al., 2020Incidence and spread of Huanglongbing (HLB) or citrus greening disease in relation to the distribution and fluctuation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) population in a citrus orchard in Sarawak, MalaysiaSerangga, 2020
-
Stelinski, 2019Ecological aspects of the vector-borne bacterial disease, citrus greening (Huanglongbing): dispersal and host use by Asian citrus psyllid, Diaphorina citri KuwayamaInsects, 2019
Thumbnail
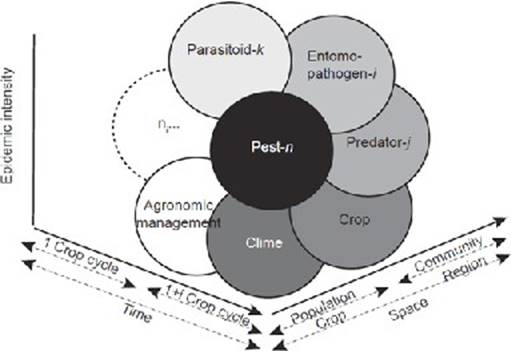
Figure 1
Epidemiological System defined by the interaction of factors that determine an epidemic process: pest species, entomopathogen, predator, and parasitoid, crop, agronomic management, climate, and any other specific factor (ni), all of which operate at different spatial-temporal levels (Adapted from Mora-Aguilera et al., 2017).
Epidemiological System defined by the interaction of factors that determine an epidemic process: pest species, entomopathogen, predator, and parasitoid, crop, agronomic management, climate, and any other specific factor (ni), all of which operate at different spatial-temporal levels (Adapted from Mora-Aguilera et al., 2017).
Under such circumstances, wind currents were one of the factors favoring the rapid spread of HLB in Mexico (Mora et al., 2013).
Overwintering adult D. citri, especially females, have a longer survival time (8-9 months) and congregate on the undersides of leaves. Nymphs are sedentary, but newly emerged individuals may migrate to neighboring shoots or move within the same shoot, but never to mature leaves (Yang et al., 2006). An aspect of ecological interest for vector and CLas management is that both males and females mate multiple times with different partners, and there is evidence that the female emits a sexual pheromone to attract her mate (Wenninger and Hall, 2008). Egg production in D. citri varies depending on the host. For example, Tsai and Liu (2000) obtained an average of 857.9 eggs on grapefruit, while Nava et al. (2007) recorded 348.4 eggs on Murraya paniculata.
-
Yang et al., 2006Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: a status report for ChinaInternational Journal of Pest Management, 2006
-
Wenninger and Hall, 2008Importance of multiple mating to female reproductive output in Diaphorina citriPhysiological Entomology, 2008
-
Tsai and Liu (2000Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plantsJournal of Economic Entomology, 2000
-
Nava et al. (2007Biology of Diaphorina citri (Hem., Psyllidae) on different hosts and at different temperaturesJournal of Applied Entomology, 2007
Diaphorina citri eggs are exclusively laid on vegetative shoots, and nymphs only develop on them; thus, their population fluctuation is positively correlated with these phenological periods (Leong et al., 2020). The greatest damage from D. citri usually occurs during autumn leaf flush (Yang et al., 2006; Godoy-Ceja and Cortez-Madrigal, 2018). Under tropical conditions such as those in India, up to nine generations can occur. Based on temperature, favorable psyllid days, and planted area, estimates for Mexico suggest anywhere from 15 to 34 annual generations (Díaz-Padilla et al., 2014). Diaphorina citri is strongly attracted to the color yellow (Patt et al., 2015).
-
Leong et al., 2020Incidence and spread of Huanglongbing (HLB) or citrus greening disease in relation to the distribution and fluctuation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) population in a citrus orchard in Sarawak, MalaysiaSerangga, 2020
-
Yang et al., 2006Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: a status report for ChinaInternational Journal of Pest Management, 2006
-
Godoy-Ceja and Cortez-Madrigal, 2018Potencial de Asclepias curassavica L. (Apocynaceae) en el control biológico de plagasRevista Mexicana de Ciencias Agrícolas, 2018
-
Díaz-Padilla et al., 2014Áreas de abundancia potencial en México del vector del huanglongbing, Diaphorina citri (Hemiptera: Liviidae)Revista Mexicana de Ciencias Agrícolas, 2014
-
Patt et al., 2015Efficacy of an autodisseminator of an entomopathogenic fungus, Isaria fumosorosea, to suppress Asian citrus psyllid, Diaphorina citri, under greenhouse conditionsBiological Control, 2015
The relationship between D. citri and CLas is of a propagative-circulative nature (Galdeano et al., 2020), and once the pathogen is acquired, the insect remains infective throughout its life; however, transovarial transmission has not been recorded (Halbert and Manjunath, 2004). According to Inoue et al. (2009), insects that acquire the bacterium in the adult stage show poor efficiency as vectors; conversely, those that acquire the pathogen in the nymphal stage are more efficient as vectors. The low concentration of bacteria in the adult insect led to low transmission to the plant; however, when the psyllid feeds in the nymphal stage on an infected plant, the bacteria concentration increases significantly, and transmission occurs in up to 67% of healthy plants. The incubation period of CLas in the host can be up to six months (Aleman et al., 2007).
-
Galdeano et al., 2020Friend or foe? Relationship between ‘Candidatus Liberibacter asiaticus’ and Diaphorina citriTropical Plant Pathology, 2020
-
Halbert and Manjunath, 2004Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in FloridaFlorida Entomologist, 2004
-
Inoue et al. (2009Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stageAnnals of Applied Biology, 2009
-
Aleman et al., 2007Diaphorina citri y la enfermedad huanglongbing: una combinación destructiva para la producción citrícolaRevista de Protección Vegetal, 2007
It has been proven that CLas requires amino acids obtained from the host plant and from D. citri. There is ample evidence that CLas can manipulate the behavior and performance of the vector, thus increasing the transmissibility and spread of the pathogen. This occurs both within the vector and through intermediation of the infected plant. Positive vectors exhibit higher fertility, fecundity, and developmental rates; however, survival is reduced. Nevertheless, CLas favors the increase in positive populations of D. citri (Yamada et al., 2019).
-
Yamada et al., 2019Diaphorin, a polyketide synthesized by an intracellular symbiont of the Asian citrus psyllid, is potentially harmful for biological control agentsPLoS One, 2019
Other intriguing aspects of the citrus-D. citri-CLas relationship include the fact that positive adult insects exhibit greater flight capability than negative psyllids. Their preference for volatiles in diseased plants is evident; however, once infected, the insects disperse towards healthy plants (Yamada et al., 2019). This, coupled with the females’ avoidance of infested shoots, facilitates disease spread. The psyllid is initially distributed in clusters, then randomly, and once again in clusters (Leong et al., 2020). CLas manipulation extends even to mating; for instance, D. citri males showed greater attraction to positive females than to healthy ones. Additionally, the parasitoid T. radiata is primarily attracted to diseased plants (Yamada et al., 2019). The psyllid’s immune system is also altered by CLas. In positive adults, the concentration of oxidases and esterases is significantly reduced. These enzymes are crucial for insecticide detoxification; hence, positive insects are more susceptible to insecticides, including entomopathogenic fungi (Yamada et al., 2019). This has practical implications for NE conservation; for example, using entomopathogenic fungi or minimal doses of insecticides targeted at positive insects would cause less harm to NE.
-
Yamada et al., 2019Diaphorin, a polyketide synthesized by an intracellular symbiont of the Asian citrus psyllid, is potentially harmful for biological control agentsPLoS One, 2019
-
Leong et al., 2020Incidence and spread of Huanglongbing (HLB) or citrus greening disease in relation to the distribution and fluctuation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) population in a citrus orchard in Sarawak, MalaysiaSerangga, 2020
Like other vectors, D. citri’s immune system is incomplete; thus, the insect’s associated endosymbionts contribute to the production of defense metabolites against invaders, such as CLas and NE (Yamada et al., 2019). In positive psyllids, the production of “diaphorin,” a polyketide with toxic effects on insects, increased; therefore, it could limit the activity of some psyllid NE, such as the coccinellid Harmonia axyridis (Galdeano et al., 2020). No research has been conducted to elucidate these interactions, such as levels of parasitism by T. radiata on positive psyllids.
-
Yamada et al., 2019Diaphorin, a polyketide synthesized by an intracellular symbiont of the Asian citrus psyllid, is potentially harmful for biological control agentsPLoS One, 2019
-
Galdeano et al., 2020Friend or foe? Relationship between ‘Candidatus Liberibacter asiaticus’ and Diaphorina citriTropical Plant Pathology, 2020
For D. citri, 59 host species from the Rutaceae family have been recorded (Halbert and Manjunath, 2004). Additionally, the psyllid can feed on Celtis spp. and Solanum tuberosum (Stelinski, 2019). Recently, Cordia myxa (Boraginaceae) was reported as a host for D. citri (Arshad et al., 2019), suggesting that the psyllid (and probably CLas) may have previously unrecorded hosts, particularly in regions where the D. citri-CLas complex has been recently introduced.
-
Halbert and Manjunath, 2004Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in FloridaFlorida Entomologist, 2004
-
Stelinski, 2019Ecological aspects of the vector-borne bacterial disease, citrus greening (Huanglongbing): dispersal and host use by Asian citrus psyllid, Diaphorina citri KuwayamaInsects, 2019
-
Arshad et al., 2019Cordia myxa L., a New Host Plant Record for Asian Citrus Psyllid, Diaphorina citri KuwayamaSouthwestern Entomologist, 2019
Grafting experiments have demonstrated the susceptibility of nearly all cultivated citrus to CLas. Citrus sinensis is typically the most susceptible species, while C. aurantiifolia is the most tolerant (Thakuria et al., 2023). Ornamental species of Murraya spp. are also hosts for CLas. Although M. paniculata can act as a bridge host in CLas dissemination, the significant decrease in bacterial titer after five months suggests that this species is of little importance as a CLas host (Damsteegt et al., 2010). In contrast, M. koenigii showed tolerance to CLas, suggesting its use as a trap plant (Beloti et al., 2018).
-
Thakuria et al., 2023Citrus Huanglongbing (HLB): Diagnostic and management optionsPhysiological and Molecular Plant Pathology, 2023
-
Damsteegt et al., 2010Murraya paniculata and related species as potential hosts and inoculum reservoirs of ‘Candidatus Liberibacter asiaticus’, causal agent of huanglongbingPlant Disease, 2010
Moreover, CLas was successfully transmitted via Cuscuta spp. to plants outside the Rutaceae family, such as Catharanthus roseus (Apocynaceae) and different solanaceous plantas. In Pithecillobium lucidum (Fabaceae) CLas was recorded even though no psyllids were recorded (Thakuria et al., 2023). Additionally, CLas was detected via PCR in Cleome rutidosperma (Capparaceae), Pisonia aculeata (Nyctaginaceae), and Trichostigma actandrum (Phytolaccaceae) without records of psyllids or HLB symptoms (Brown et al., 2014).
-
Thakuria et al., 2023Citrus Huanglongbing (HLB): Diagnostic and management optionsPhysiological and Molecular Plant Pathology, 2023
-
Brown et al., 2014First report of ‘Candidatus Liberibacter asiaticus’ associated with huanglongbing in the weeds Cleome rutidosperma, Pisonia aculeata and Trichostigma octandrum in JamaicaJournal of Citrus Pathology, 2014
In 2010, adult D. citri were recorded on glandular trichomes of Sicydium tamnifolium (Cucurbitaceae) on M. paniculata plants in Chetumal, Q.R., Mexico. This species is distributed only along the Gulf of Mexico and Chiapas (Lira, 2004). Although its role as a host for D. citri is unknown, these preliminary observations suggest that S. tamnifolium could be utilized as a trap plant for D. citri and other pests. However, it is necessary to elucidate the pros and cons of its use, such as interference with biological control, crop interference, potential pest reservoir, and economic considerations.
-
Lira, 2004El género Sycidium (Cucurbitaceae, Zanonioideae, Zanonieae, Sicydiinae) en MéxicoActa Botánica Mexicana, 2004
Management of Diaphorina citri-CLas
Once CLas infects its host, it is difficult to eradicate, despite numerous studies demonstrating control possibilities, such as through cultural, chemical, and biological control; thermotherapy, defense inducers, and genetic manipulation for resistance (Thakuria et al., 2023). However, in practice, HLB management includes preventive measures, including the production of healthy plants (Yang et al., 2006), eradication of diseased plants, and legal control (García-Ávila et al., 2021), but primarily through vector control (Yang et al., 2006; García-Ávila et al., 2021). Although integrated management of D. citri is proposed, greater attention is focused on chemical control (Qureshi and Stansly, 2007; Parra and Coelho, 2019), with biological control only being complementary (Etienne et al., 2001; Salcedo-Baca et al., 2010). However, when CLas is present in commercial orchards, chemical control is recommended (Parra and Coelho, 2019; García-Ávila et al., 2021).
-
Thakuria et al., 2023Citrus Huanglongbing (HLB): Diagnostic and management optionsPhysiological and Molecular Plant Pathology, 2023
-
Yang et al., 2006Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: a status report for ChinaInternational Journal of Pest Management, 2006
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
-
Qureshi and Stansly, 2007Integrated approaches for managing the Asian citrus psyllid Diaphorina citri (Homoptera: Psyllidae) in FloridaProceedings of the Florida State Horticultural Society, 2007
-
Parra and Coelho, 2019Applied biological control in Brazil: from laboratory assays to field applicationJournal of Insect Science, 2019
-
Etienne et al., 2001Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae)Fruits, 2001
-
Salcedo-Baca et al., 2010Evaluación del Impacto Económico de la Enfermedad de los Cítricos Huanglongbing (HLB) en la Cadena Citrícola MexicanaInstituto Interamericano de Cooperación para la Agricultura (IICA), 2010
Following the introduction of the D. citri-CLas complex in Brazil, insecticide applications increased by over 600% (Parra et al., 2016). Seen in this light, chemical control could have a detrimental impact on the environment and on natural enemy populations (Qureshi and Stansly, 2009; Monzó and Stansly, 2020). For disease vectors, the ET is practically the absence of pests. For instance, the transmission rate of CLas by adult D. citri to Citrus depressa plants was estimated at 8.5% when one adult was introduced per plant, with a two-week inoculation access period (Ukuda-Hosokawa et al., 2015). Only a few ET have been proposed for its control; for example, in Florida, five adults and three nymphs per flush are suggested (Halbert and Manjunath, 2004), or 0.2-0.7 adults per flush and 0.2 adults per trap (Monzo and Stansly, 2015). In Mexico, a proposed ET is one adult per trap (Yzquierdo-Álvarez, 2021).
-
Parra et al., 2016Tamarixia radiata (Hymenoptera: Eulophidae) × Diaphorina citri (Hemiptera: Liviidae): mass rearing and potential use of the parasitoid in BrazilJournal of Integrated Pest Management, 2016
-
Qureshi and Stansly, 2009Incidence of invasive Diaphorina citri (Hemiptera: Psyllidae) and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrusJournal of Economic Entomology, 2009
-
Monzó and Stansly, 2020Economic value of conservation biological control for management of the Asian citrus psyllid, vector of citrus Huanglongbing diseasePest Management Science, 2020
-
Ukuda-Hosokawa et al., 2015Infection density dynamics of the citrus greening bacterium “Candidatus Liberibacter asiaticus” in field populations of the psyllid Diaphorina citri and its relevance to the efficiency of pathogen transmission to citrus plantsApplied and Environmental Microbiology, 2015
-
Halbert and Manjunath, 2004Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in FloridaFlorida Entomologist, 2004
-
Yzquierdo-Álvarez, 2021Fluctuación espacio-temporal de Diaphorina citri Kuwayama (Hemiptera: Liviidae) en limón persa (Citrus latifolia) en la zona citrícola de Huimanguillo, TabascoActa Zoológica Mexicana, 2021
Diaphorina citri has a wide range of natural enemies, including predators, parasitoids, and entomopathogens (Yang et al., 2006; Qureshi and Stansly, 2007), which presumably should also reduce disease incidence and prevalence.
-
Yang et al., 2006Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: a status report for ChinaInternational Journal of Pest Management, 2006
-
Qureshi and Stansly, 2007Integrated approaches for managing the Asian citrus psyllid Diaphorina citri (Homoptera: Psyllidae) in FloridaProceedings of the Florida State Horticultural Society, 2007
Entomophagous. Records worldwide document 95 arthropod species (spiders and insects) across nine orders and 23 families (Kondo et al., 2015), with varying impacts on D. citri. For instance, in Saudi Arabia, the Order Araneae contributed up to 33.6% to vector reduction (Halbert and Manjunath, 2004). In Florida, predators contribute up to 100% to D. citri control (Qureshi et al., 2009). These include species of Araneae, Hemerobiidae, Syrphidae, and Anthocoridae, with lady beetles such as H. axyridis, Olla v-nigrum, and Cycloneda sanguinea being notable (Michaud, 2004). In Colombia, 16 natural enemy species from six families and five orders affecting D. citri have been recorded (Kondo et al., 2015). Conversely, extensive insecticide applications in Brazil limited the impact of predators on psyllid biological control (Parra et al., 2016). In Mexico, 20 species of entomophagous insects have been recorded, including families Coccinellidae, Chrysopidae, Anthocoridae, and Vespidae, with the first two being more prevalent. Additionally, entomopathogenic fungi such as Beauveria bassiana and Hirsutella citriformis have been isolated from D. citri (Table 1).
-
Kondo et al., 2015A checklist of natural enemies of Diaphorina citri Kuwayama (Hemiptera: Liviidae) in the department of Valle del Cauca, Colombia and the worldInsecta Mundi, 2015
-
Halbert and Manjunath, 2004Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in FloridaFlorida Entomologist, 2004
-
Qureshi et al., 2009Incidence of invasive Diaphorina citri (Hemiptera: Psyllidae) and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrusJournal of Economic Entomology, 2009
-
Michaud, 2004Natural mortality of Asian citrus psyllid (Homoptera: Psyllidae) in central FloridaBiological Control, 2004
-
Parra et al., 2016Tamarixia radiata (Hymenoptera: Eulophidae) × Diaphorina citri (Hemiptera: Liviidae): mass rearing and potential use of the parasitoid in BrazilJournal of Integrated Pest Management, 2016
Table 1
Natural enemies associated with D. citri in citrus orchards in Mexico.
Natural enemies associated with D. citri in citrus orchards in Mexico.
| Family/Species | Reference |
|---|---|
| Coccinellidae | |
| H. axyridis | Mora-Aguilera, 2013 |
| O. v-nigrum | |
| C. sanguinea | Cortez-Mondaca et al., 2010 |
| Chilocorus cacti | Mora-Aguilera, 2013 |
| Hippodamia convergens | |
| Scymnus sp. | |
| Arawana sp. | Lozano-Contreras y Jasso-Argumedo, 2012 |
| Azya orbigera | |
| Delphastus sp. | |
| Zagloba sp. | |
| Chrysopidae | |
| Ceraeocrhysa sp. | Lozano-Contreras y Jasso-Argumedo, 2012 |
| C. cubana | |
| C. claveri | |
| C. valida | |
| C. everes | |
| Chrysoperla rufrilabris | Cortez-Mondaca et al., 2010 |
| Ch. comanche | |
| Chrysoperla sp. | Mora-Aguilera, 2013 |
| Anthocoridae Orius spp. | Cortez-Mondaca et al., 2011 |
| Eulophidae T. radiata | |
| Encyrtidae Diaphorencyrtus spp. | |
| Globorotallidae H. citriformis | Mora-Aguilera et al., 2017 |
| Clavicitaceae B. bassiana |
The primary focus of biological control against D. citri has been the introduction of its parasitoids, T. radiata and Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) (Halbert and Manjunath, 2004), although the former has received more attention. However, parasitism levels have been variable, ranging from 28.9-36.4% in Taiwan (Chiu et al., 1988) to 100% in Reunion Island, France (Etienne et al., 2001).
-
Halbert and Manjunath, 2004Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in FloridaFlorida Entomologist, 2004
-
Chiu et al., 1988Attempts to establish Tetrastichus radiatus Waterson (Hymenoptera, Chalcidoidea), a primary parasite of Diaphorina citri Kuwayama in TaiwanInternational Organization of Citrus Virologists Conference Proceedings, 1988
-
Etienne et al., 2001Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae)Fruits, 2001
In Florida, T. radiata has been a relatively minor mortality factor for D. citri, with levels below 20% in spring-summer, but reaching 39-56% in autumn (Qureshi et al., 2009). In Puerto Rico, Pluke et al. (2008) reported parasitism levels of 70- 100%, while in Sinaloa, Mexico, natural parasitism rates of up to 59.6% were recorded (Cortez-Mondaca et al., 2010). The effectiveness of T. radiata is strongly limited by the presence of hyperparasitoids (Aubert, 1987), pesticide use (Diniz et al., 2020), absence of hosts in critical periods (Leong et al., 2020), and lack of nectar and pollen (Irvin and Hoddle, 2021).
-
Qureshi et al., 2009Incidence of invasive Diaphorina citri (Hemiptera: Psyllidae) and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrusJournal of Economic Entomology, 2009
-
Pluke et al. (2008Citrus flushing patterns, Diaphorina citri (Hemiptera: Psyllidae) populations and parasitism by Tamarixia radiata (Hymenoptera: Eulophidae) in Puerto RicoFlorida Entomologist, 2008
-
Cortez-Mondaca et al., 2010Primer reporte de enemigos naturales y parasitismo sobre Diaphorina citri Kuwayama en Sinaloa, MéxicoSouthwestern Entomologist, 2010
-
Aubert, 1987Trioza erytreae Del Guercio and Diaphorina citri Kuwayama (Homoptera: Psylloidea), the two vectors of citrus greening disease: biological aspects and possible control strategiesFruits, 1987
-
Diniz et al., 2020The enemy is outside: Releasing the parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in external sources of HLB inocula to control the Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae)Neotropical Entomology, 2020
-
Leong et al., 2020Incidence and spread of Huanglongbing (HLB) or citrus greening disease in relation to the distribution and fluctuation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) population in a citrus orchard in Sarawak, MalaysiaSerangga, 2020
-
Irvin and Hoddle, 2021Evaluating the potential of flowering plants for enhancing predatory hoverflies (Syrphidae) for biological control of Diaphorina citri (Liviidae) in CaliforniaBiological Control, 2021
Due to the variability in T. radiata parasitism, the greatest effort has been focused on mass rearing and release (Parra et al., 2016; Flores and Ciomperlik, 2017; García-Ávila et al., 2021). However, in commercial orchards, this strategy is incompatible with pesticides (Diniz et al., 2020). Thus, biological control is only practiced in backyard orchards, urban areas, and abandoned plantations, with scientific evidence of reducing D. citri and HLB incidence in commercial orchards (Parra et al., 2016; Diniz et al., 2020; García-Ávila et al., 2021). For example, in Texas, USA, after releasing T. radiata, vector reduction exceeded 91% (Flores and Ciomperlik, 2017). In countries like Brazil, parasitism levels reached 72.5% after parasitoid release (Parra et al., 2016). In Mexico, it is mentioned that natural parasitism by T. radiata (3-26%) increased to 85% after mass releases (García- Ávila et al., 2021).
-
Parra et al., 2016Tamarixia radiata (Hymenoptera: Eulophidae) × Diaphorina citri (Hemiptera: Liviidae): mass rearing and potential use of the parasitoid in BrazilJournal of Integrated Pest Management, 2016
-
Flores and Ciomperlik, 2017Biological control using the ectoparasitoid, Tamarixia radiata, against the Asian citrus psyllid, Diaphorina citri, in the Lower Rio Grande Valley of TexasSouthwestern Entomologist, 2017
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
-
Diniz et al., 2020The enemy is outside: Releasing the parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in external sources of HLB inocula to control the Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae)Neotropical Entomology, 2020
Entomopathogenic fungi. Prominent among these species are H. citriformis, B. bassiana (Padulla and Alves, 2021), Isaria fumosorosea (Gallou et al., 2016), and Metarhizium anisopliae (Ayala-Zermeño et al., 2015). However, only H. citriformis, B. bassiana, and I. fumosorosea (=Paecilomyces fumosoroseus) have been isolated from the insect (Halbert and Manjunath, 2004; Meyer et al., 2008; Mora-Aguilera et al., 2017). These entomopathogenic fungi have mainly been evaluated using inundative strategy (i.e., spraying); in Brazil, their field application achieved mortality levels of 57.8% to 96.1%, directly correlated with humidity and inversely with temperature (Saldarriaga-Ausique et al., 2017).
-
Padulla and Alves, 2021Suscetibilidade de ninfas de Diaphorina citri a fungos entomopatogênicosArquivos do Instituto Biológico, 2021
-
Gallou et al., 2016Species clarification of Isaria isolates used as biocontrol agents against Diaphorina citri (Hemiptera: Liviidae) in MexicoFungal Biology, 2016
-
Ayala-Zermeño et al., 2015Characterisation of entomopathogenic fungi used in the biological control programme of Diaphorina citri in MexicoBiocontrol Science and Technology, 2015
-
Halbert and Manjunath, 2004Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in FloridaFlorida Entomologist, 2004
-
Meyer et al., 2008Isolation and characterization of an Isaria fumosorosea isolate infecting the Asian citrus psyllid in FloridaJournal of Invertebrate Pathology, 2008
-
Saldarriaga-Ausique et al., 2017Efficacy of entomopathogenic fungi against adult Diaphorina citri from laboratory to field applicationsJournal of Pest Science, 2017
In Mexico, EPFs represent a significant tool against D. citri and CLas in commercial citrus orchards (García-Ávila et al., 2021). After assessing Cordyceps bassiana, M. anisopliae, and I. fumosorosea in Colima, Mexico, the highest mortality in nymphs (60%) and adults (50%) was achieved with C. bassiana (Lezama-Gutiérrez et al., 2012). Under field conditions, I. javanica (=fumosorosea) achieved mortality rates of 66-81.8% (Gallou et al., 2016).
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
-
Lezama-Gutiérrez et al., 2012Use of the entomopathogenic fungi Metarhizium anisopliae, Cordyceps bassiana and Isaria fumosorosea to control Diaphorina citri (Hemiptera: Psyllidae) in Persian lime under field conditionsInternational Journal of Tropical Insect Science, 2012
-
Gallou et al., 2016Species clarification of Isaria isolates used as biocontrol agents against Diaphorina citri (Hemiptera: Liviidae) in MexicoFungal Biology, 2016
EPFs can also colonize plant tissues. Bamisile et al. (2021) assessed the endophytic capability of B. bassiana and I. fumosorosea, finding that only the former colonized citrus tissues. Seven days after inoculation, mortality in D. citri adults ranged from 10% to 15%, and there was a significant reduction in egg deposition by females feeding on treated plants.
-
Bamisile et al. (2021General limitations to endophytic entomopathogenic fungi use as plant growth promoters, pests and pathogens biocontrol agentsPlants, 2021
Beyond biological control of the D. citri-HLB pathosystem: the conservation of natural enemies
There’s a prevailing notion that once CLas takes hold, the biological control (BC) of D. citri becomes inconsequential for managing HLB (Helbert and Manjunath, 2004; Yang et al., 2006). This might hold true if we consider BC as simply mirroring chemical control: aiming to drastically reduce pest populations. However, shifting away from the conventional and atomistic paradigm of pest management and its derived inundative biological control (IBC), we can discern a natural phenomenon of population self-regulation so potent that conservation biological control (CBC) should form the bedrock of IPM.
-
Yang et al., 2006Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: a status report for ChinaInternational Journal of Pest Management, 2006
CBC encompasses all measures aimed at environmental modification and pesticide management to favor the conservation and proliferation of beneficial organisms (Gurr and You, 2016). Despite being the linchpin of biological control, CBC garners minimal attention (Cortez-Madrigal and Gutiérrez-Cárdenas, 2023). This is largely due to modern agricultural practices being geared towards productivity rather than the preservation of beneficial organisms. Various strategies are proposed for their conservation, including pesticide management, devices for in situ conservation and augmentation of parasitoids (Kehrli et al., 2005), provision of alternate food sources such as nectar, pollen (Gurr and Wratten, 1999; Gurr and You, 2016) and alternate hosts (Cortez-Madrigal and Gutiérrez-Cárdenas, 2023).
-
Gurr and You, 2016Conservation biological control of pests in the molecular era: New opportunities to address old constraintsFrontiers in Plant Science, 2016
-
Cortez-Madrigal and Gutiérrez-Cárdenas, 2023Enhancing biological control: conservation of alternative hosts of natural enemiesEgyptian Journal of Biological Pest Control, 2023
-
Kehrli et al., 2005Mass-emergence devices: a biocontrol technique for conservation and augmentation of parasitoidsBiological Control, 2005
When beneficial organisms lack alternative food sources in a crop, they migrate to areas where sustenance is available, undercutting the presence of functional populations of beneficial organisms (Cortez-Madrigal and Gutiérrez-Cárdenas, 2023) or delaying their arrival to such an extent that BC becomes ineffective (Doutt and Nakata, 1973). This scenario can be altered by redesigning the agroecosystem to support the permanent establishment of parasitoids, predators, and entomopathogens before the pest appears, not only of insects and mites but of the entire spectrum of organisms involved in population self-regulation within ecosystems; a holistic view of CBC: “beyond biological control” (Cortez-Madrigal and Gutiérrez-Cárdenas, 2023).
-
Cortez-Madrigal and Gutiérrez-Cárdenas, 2023Enhancing biological control: conservation of alternative hosts of natural enemiesEgyptian Journal of Biological Pest Control, 2023
-
Doutt and Nakata, 1973The rubus leafhopper and its egg parasitoid: An endemic biotic system useful in grape-pest managementEnvironmental Entomology, 1973
Though the economic benefits of CBC in regulating D. citri populations have been validated (Monzó and Stansly, 2020), there’s a dearth of documented studies (Irvin et al., 2021; Irvin and Hoddle, 2021). Some conservation strategies for beneficial organisms evaluated in D. citri or related pests include nectar-rich flora, alternate hosts for beneficial organisms, in-situ conservation and augmentation devices for parasitoids, and the self-propagation of entomopathogens.
-
Monzó and Stansly, 2020Economic value of conservation biological control for management of the Asian citrus psyllid, vector of citrus Huanglongbing diseasePest Management Science, 2020
-
Irvin et al., 2021Evaluating the potential of flowering plants for enhancing predatory hoverflies (Syrphidae) for biological control of Diaphorina citri (Liviidae) in CaliforniaBiological Control, 2021
-
Irvin and Hoddle, 2021The effects of floral nectar, extrafloral nectar and hemipteran honeydew on the fitness of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of Diaphorina citriBiological Control, 2021
Unlike classical and inundative biological control, CBC is often cost-effective; it may even enhance the other two BC strategies. It’s been argued, for instance, that the failures of various classical BC programs stem from the absence of alternate hosts to sustain newly introduced beneficial organisms during specific periods (Gurr and Wratten, 1999). Could this be the case for T. radiata?
-
Gurr and Wratten, 1999Forum ‘Integrated biological control’: A proposal for enhancing success in biological controlInternational Journal of Pest Management, 1999
Should agroecosystem conditions favor the establishment of beneficial organisms, IBC could also be bolstered with fewer releases. The proposition integrating classical, inundative, and conservation biological control has been dubbed “integrated biological control” (Gurr and Wratten, 1999). Citrus cultivation in Mexico could serve as an initial model for integrated biological control.
-
Gurr and Wratten, 1999Forum ‘Integrated biological control’: A proposal for enhancing success in biological controlInternational Journal of Pest Management, 1999
Enhancing the biological control of D. citri
Inclusion and conservation of nectariferous plants. Due to prevailing crop conditions, the parasitoid T. radiata is necessarily absent from citrus for much of the year. One strategy for its conservation involves incorporating nectar-bearing plants to attract beneficial insect populations. However, not all nectar-bearing species are equally attractive, necessitating specific studies. For instance, Irvin et al. (2021), after evaluating four flowering plant species for the conservation of Allograpta obliqua (Diptera: Syrphidae), found that only two were attractive to the key predator of D. citri in California. Orchards with floral plants present recorded
-
Irvin et al. (2021Evaluating the potential of flowering plants for enhancing predatory hoverflies (Syrphidae) for biological control of Diaphorina citri (Liviidae) in CaliforniaBiological Control, 2021
2.5 times more predators and 3.5 times more oviposition than those without. When fed with nectar from Fagopyrum esculentum, the survival of the parasitoid T. radiata significantly increased, and females laid five times more eggs than those fed with other sugar sources. Thus, the addition of nectar-bearing plants in citrus orchards could enhance T. radiata efficiency (Irvin and Hoddle, 2021). In northwest Michoacán, Mexico, out of over 27 species of flowering plants evaluated, only 24 registered parasitoids. Annual plants like Stevia serrata, shrubs like Senecio salignus and Budleja sp., trees like Thouinia villosa, and climbers like Serjania racemosa stood out. Over 14 families of parasitic hymenoptera were recorded on the flowers of these plants (Unpublished data).
-
Irvin and Hoddle, 2021Evaluating the potential of flowering plants for enhancing predatory hoverflies (Syrphidae) for biological control of Diaphorina citri (Liviidae) in CaliforniaBiological Control, 2021
The addition of nectar and pollen alone does not guarantee the prevalence of beneficial insects in agroecosystems. In the absence of prey or hosts for beneficial insects, they, being entomophagous, will be unable to complete their life cycle or will venture away in search of food, particularly parasitoids like T. radiata. Thus, the conservation of alternate hosts is essential for the persistence of beneficial insects alongside agroecosystems.
Conservation of Alternate Hosts. An example similar to D. citri was the biological control of T. erytreae using the parasitoid T. dryi in Trinidad Island. Trioza eastopi feeds on the weed Litsea chinensis (Lauraceae) and serves as an alternate host for the parasitoid, which managed to increase its population and successfully control the pest (Duran-Vila and Bové, 2015).
-
Duran-Vila and Bové, 2015Citrus HLB is an emerging disease transmitted by psyllid vectors. Can it be prevented? If not, can it be managed?International Centre for Advanced Mediterranean Agronomic Studies. Watch Lett, 2015
The species Asclepias curassavica (Apocynaceae) hosts specialist phytophagous (>9), including orders Hemiptera, Lepidoptera, Diptera, Coleoptera, and subclass Acarina. It encompasses species such as Aphis nerii, Danaus plexippus (Lepidoptera: Nymphalidae), Euchaetes antica (Lepidoptera: Erebidae), Liriomyza asclepiadis (Diptera: Agromyzidae), and the mite Eotetranychus typhae (Tetranychidae), among others (Figure 2) (Cortez-Madrigal et al., 2016).
-
Cortez-Madrigal et al., 2016Conserving phytophagous arthropods to conserve natural enemies: Asclepias curassavica as the modelSouthwestern Entomologist, 2016
In field conditions at Cienega de Chapala, Michoacán, specialized phytophagous of A. curassavica recruited over 20 species of natural enemies in citrus orchards, including egg parasitoids, aphids, and leaf miners, as well as a wide diversity of predators, including families Coccinellidae, Syrphidae, Chamaemyiidae, Chrysopidae, Staphylinidae, Cecidomyiidae, and orders Thysanoptera, Hemiptera, and mites of the family Phytoseiidae, among others (Figure 2) (Cortez-Madrigal et al., 2016). Among the recorded ladybird beetles, O. v-nigrum, H. convergens, and sanguinea are significant predators of D. citri (Michaud, 2004; Cortez-Mondaca et al., 2010).
-
Cortez-Madrigal et al., 2016Conserving phytophagous arthropods to conserve natural enemies: Asclepias curassavica as the modelSouthwestern Entomologist, 2016
-
Michaud, 2004Natural mortality of Asian citrus psyllid (Homoptera: Psyllidae) in central FloridaBiological Control, 2004
Studies conducted in citrus orchards recorded a significant reduction in pests of Persian lime cultivation, Citrus x latifolia, when the plant A. curassavica was incorporated, including D. citri. One explanation provided was the abundance of natural enemies recruited by A. curassavica, primarily from the families Coccinellidae, Syrphidae, Chamaemyiidae, and Chrysopidae (Godoy-Ceja and Cortez-Madrigal, 2018).
-
Godoy-Ceja and Cortez-Madrigal, 2018Potencial de Asclepias curassavica L. (Apocynaceae) en el control biológico de plagasRevista Mexicana de Ciencias Agrícolas, 2018
Thumbnail

Figure 2
Aphis nerii on Asclepias curassavica and some associated natural enemies. A) Pseudodorus clavatus, B) Cycloneda sanguinea, C) Adult of Oligota sp., D) Adult of Chamaemyiidae, E) Mummies and adults of Lyciphlebus testaceipes.
Aphis nerii on Asclepias curassavica and some associated natural enemies. A) Pseudodorus clavatus, B) Cycloneda sanguinea, C) Adult of Oligota sp., D) Adult of Chamaemyiidae, E) Mummies and adults of Lyciphlebus testaceipes.
The monarch butterfly emerged as a significant host for the egg parasitoid Trichogramma pretiosum (Trichogrammatidae), exhibiting multiple egg parasitism. In the study region, T. pretiosum was present for much of the year (Cortez-Madrigal et al., 2016), which could explain the low incidence of lepidopteran larvae pests in citrus (Godoy-Ceja and Cortez-Madrigal, 2018).
-
Cortez-Madrigal et al., 2016Conserving phytophagous arthropods to conserve natural enemies: Asclepias curassavica as the modelSouthwestern Entomologist, 2016
-
Godoy-Ceja and Cortez-Madrigal, 2018Potencial de Asclepias curassavica L. (Apocynaceae) en el control biológico de plagasRevista Mexicana de Ciencias Agrícolas, 2018
Devices for preserving parasitoids in situ . The objective is to conserve in situ parasitoids of sessile or low-mobility pests. In containers with emergence openings and selective mesh, infested plant material is deposited, allowing only the parasitoid’s emergence (Kehrli et al., 2005). Another modification involves cages covering entire plants, adapted with emergence openings with selective mesh (Desurmont et al., 2022), as mentioned earlier.
-
Kehrli et al., 2005Mass-emergence devices: a biocontrol technique for conservation and augmentation of parasitoidsBiological Control, 2005
-
Desurmont et al., 2022Identifying an optimal screen mesh to enable augmentorium-based enhanced biological control of the olive fruit fly Bactrocera oleae (Diptera: Tephritidae) and the mediterranean fruit fly Ceratitis capitata (Diptera: Tephritidae)Journal of Insect Science, 2022
For the conservation of T. triozae (Hymenoptera: Eulophidae), a parasitoid of the tomato-potato psyllid (Bactericera cockerelli; Hemiptera: Triozidae), it was determined that a mesh opening of 500 µm allowed up to 89% parasitoid emergence and retained adult psyllids (Figure 3). In field conditions of tomato cultivation, the device promoted >80% parasitism of the psyllid (Unpublished data).
The same device with minor modifications could enhance the biological control of D. citri. For instance, through the design and use of cages similar to those described above, covering Murraya spp. plants with populations of D. citri and T. radiata. The vegetative characteristics of M. paniculata make it suitable for mass rearing programs of T. radiata for subsequent field release (Parra et al., 2016; García-Ávila et al., 2021). In addition to T. radiata, the use of these devices could also conserve natural enemies of the psyllid. For example, during dry periods and absence of citrus flush in Yucatán, Mexico, M. paniculata hedges were where prevalence of H. citriformis was recorded (Cortez-Madrigal et al., 2013).
-
Parra et al., 2016Tamarixia radiata (Hymenoptera: Eulophidae) × Diaphorina citri (Hemiptera: Liviidae): mass rearing and potential use of the parasitoid in BrazilJournal of Integrated Pest Management, 2016
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
-
Cortez-Madrigal et al., 2013Exploración y aislamiento de hongos entomopatógenos asociados a Diaphorina citri en la Península de YucatánTópicos Selectos de Agronomía Tropical 2, 2013
Thumbnail

Figure 3
Different types of mesh evaluated in the selective emergence of Tamarixia triozae, parasitoid of Bactericera cockerelli as a model to be implemented with D. citri and its parasitoid. A) 700 x 700 μm, B) 700 x 900 μm, and C) 500 μm mesh.
Different types of mesh evaluated in the selective emergence of Tamarixia triozae, parasitoid of Bactericera cockerelli as a model to be implemented with D. citri and its parasitoid. A) 700 x 700 μm, B) 700 x 900 μm, and C) 500 μm mesh.
Devices for the preservation and self-dissemination of EPFs. Some authors suggest the use of the self-dissemination technique of entomopathogenic fungi for managing D. citri (Avery et al., 2010). However, few studies have been carried out. For instance, Patt et al. (2015) evaluated yellow devices baited with I. fumosorosea spores under greenhouse conditions. After 24 hours, 55% of adults were infected with the entomopathogen; when the mycosed cadavers were exposed to healthy nymphs, over 90% of them were infected. The blastospores on the trap remained viable for up to 20 days, which decreased when traps were directly exposed to sunlight. In another study using yellow cards impregnated with spores of the same fungus, adult mortality was less than 40%; when citrus volatiles were added to the cards, mortality increased by 23% (Moran et al., 2011).
-
Avery et al., 2010Effect of photoperiod and host distribution on the horizontal transmission of Isaria fumosorosea (Hypocreales: Cordycipitaceae) in greenhouse whitefly assessed using a novel model bioassayBiocontrol Science and Technology, 2010
-
Patt et al. (2015Efficacy of an autodisseminator of an entomopathogenic fungus, Isaria fumosorosea, to suppress Asian citrus psyllid, Diaphorina citri, under greenhouse conditionsBiological Control, 2015
-
Moran et al., 2011Localized autoinoculation and dissemination of Isaria fumosorosea for control of the Asian citrus psyllid in South TexasSubtropical Plant Science, 2011
For fungi species like H. citriformis, with low potential for inundative use, self- dissemination could favor the conservation and early development of epizootics. Epizootics of H. citriformis occur in D. citri populations in different parts of the world, with variable mortalities ranging from 75% in Florida (Hall et al., 2012) to 92% in Rovirosa, Quintana Roo, Mexico (Mora-Aguilera et al., 2017).
-
Hall et al., 2012Observations on the entomopathogenic fungus Hirsutella citriformis attacking adult Diaphorina citri (Hemiptera: Psyllidae) in a managed citrus groveBioControl, 2012
The self-dissemination of natural enemies could be facilitated by the behavior of D. citri, including mating with multiple partners (Wenninger and Hall, 2008), males’ preference for mating with positive females, greater flight capacity of positive adults (Yamada et al., 2019), and attraction to colored traps (Moran et al., 2011), among others. Additionally, the potential development of a sex pheromone by the insect could enhance the self-dissemination technique (Wenninger et al., 2008).
-
Wenninger and Hall, 2008Importance of multiple mating to female reproductive output in Diaphorina citriPhysiological Entomology, 2008
-
Yamada et al., 2019Diaphorin, a polyketide synthesized by an intracellular symbiont of the Asian citrus psyllid, is potentially harmful for biological control agentsPLoS One, 2019
-
Moran et al., 2011Localized autoinoculation and dissemination of Isaria fumosorosea for control of the Asian citrus psyllid in South TexasSubtropical Plant Science, 2011
-
Wenninger et al., 2008Behavioral evidence for a female-produced sex attractant in Diaphorina citriEntomologia Experimentalis et Applicata, 2008
Analysis and Insights
Despite the catastrophic forecasts since the appearance of HLB in 2009 (Salcedo-Baca et al., 2010), efforts undertaken in Mexico have mitigated the worst impacts of the disease on Mexican citrus cultivation. However, eight years after its appearance, the disease continues to spread, and by 2017, records existed in all 24 citrus-growing states of the country (García-Ávila et al., 2021). This underscores that the national citrus industry will always be at constant risk due to the presence of the D. citri-CLas pathosystem. Recent studies indicate that some Mexican citrus growers have opted to switch crops, which will result in a significant reduction in national citrus production (Granados-Ramírez et al., 2018).
-
Salcedo-Baca et al., 2010Evaluación del Impacto Económico de la Enfermedad de los Cítricos Huanglongbing (HLB) en la Cadena Citrícola MexicanaInstituto Interamericano de Cooperación para la Agricultura (IICA), 2010
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
-
Granados-Ramírez et al., 2018Reconversión de cultivos como resultado de la presencia de huanglongbing en Colima, MéxicoRevista Geográfica de América Central, 2018
It is clear that research on the D. citri-CLas pathosystem is far from concluded. Strategies are needed to reduce phytosanitary costs, where CBC plays a crucial role. The present review highlights the absence of studies and programs focused on conserving natural enemies for D. citri, despite demonstrating their importance for its management (Qureshi and Stansly, 2009; Irvin et al., 2021). For instance, it has been estimated that in orchards without effective CBC programs, annual losses could reach up to 2,000 USD/ha (Monzó and Stansly, 2020). For reference, the cost of insecticide application in Florida citrus reached 3,000 USD/ha (García-Ávila et al., 2021). Therefore, the long-term viability of the strategies implemented so far for HLB management should be reconsidered (Monzó and Stansly, 2020).
-
Qureshi and Stansly, 2009Exclusion techniques reveal significant biotic mortality suffered by Asian citrus psyllid Diaphorina citri (Hemiptera: Psyllidae) populations in Florida citrusBiological Control, 2009
-
Irvin et al., 2021Evaluating the potential of flowering plants for enhancing predatory hoverflies (Syrphidae) for biological control of Diaphorina citri (Liviidae) in CaliforniaBiological Control, 2021
-
Monzó and Stansly, 2020Economic value of conservation biological control for management of the Asian citrus psyllid, vector of citrus Huanglongbing diseasePest Management Science, 2020
-
García-Ávila et al., 2021Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in MexicoArea-Wide Integrated Pest Management, 2021
The strategies proposed here for CBC programs in citrus are straightforward and would not disrupt measures already implemented for managing the D. citri- CLas pathosystem. A future task is to evaluate Murraya species in conserving T. radiata by designing and using selective cages during periods of absence of D. citri nymphs in citrus. Besides protecting the parasitoid from pesticide applications, their populations would be advanced to synchronize them with the onset of nymph populations in citrus. Other studies demonstrate the feasibility of modifying the sequence of pest-NE appearance in agroecosystems to NE-pest, which is known to enhance BC (Doutt and Nakata, 1973; Jacas and Urbaneja, 2010).
-
Doutt and Nakata, 1973The rubus leafhopper and its egg parasitoid: An endemic biotic system useful in grape-pest managementEnvironmental Entomology, 1973
-
Jacas and Urbaneja, 2010Biological control in citrus in Spain: from classical to conservation biological controlIntegrated Management of Arthropod Pests and Insect Borne Diseases, 2010
There are numerous plant species that harbor specialized phytophagous insects that could serve as alternate prey for predators in citrus; that is, with low risk of becoming pests; an example is the Apocynaceae family (= Asclepiadaceae). In Mexico, the family is widely distributed with >300 species in the country (Juárez- Jaimes et al., 2007), thus presenting high potential for inclusion in D. citri CBC programs. In addition to A. curassavica, species such as A. linaria, A. angustifolia, and Mandevilla foliosa have recorded populations of A. nerii, D. plexippus, and Lygaeidae bugs. Even CLas-tolerant plants like M. koenigii could function as D. citri trap plants and harbor NE; among them, T. radiata.
In conjunction with alternate hosts, the contribution of nectar and pollen is crucial for attracting and sustaining a diverse array of predators that prey on D. citri (Kondo et al., 2015). However, the inclusion of nectar-producing plants in citrus crops for conserving D. citri’s natural enemies remains an understudied aspect (Irvin et al., 2021; Irvin and Hoddle, 2021). Pioneering studies in Mexico, as described here, could serve as a foundational step for managing the pathosystem, especially in young citrus plantations where flowering is limited. Simultaneously, it’s imperative to ensure that companion plants do not serve as hosts for D. citri or other significant pests.
-
Kondo et al., 2015A checklist of natural enemies of Diaphorina citri Kuwayama (Hemiptera: Liviidae) in the department of Valle del Cauca, Colombia and the worldInsecta Mundi, 2015
-
Irvin et al., 2021Evaluating the potential of flowering plants for enhancing predatory hoverflies (Syrphidae) for biological control of Diaphorina citri (Liviidae) in CaliforniaBiological Control, 2021
-
Irvin and Hoddle, 2021The effects of floral nectar, extrafloral nectar and hemipteran honeydew on the fitness of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of Diaphorina citriBiological Control, 2021
citri’s attraction to the yellow color presents opportunities for employing traps baited with entomopathogens, capitalizing on the insect’s natural behavior for self-dissemination. The minimal inoculum required and the negligible impact on non-target organisms suggest that self-dissemination could serve as a valuable tool in CBC programs targeting the D. citri-CLas complex. One notable advantage of self-dissemination in vectors is its potential to trigger epizootics within populations (particularly in low-density populations), which could hypothetically slow down the spread and incidence of HLB. However, comprehensive field and laboratory epidemiological studies are needed to identify the biotic and abiotic factors conducive to epizootic development. This includes characterizing of EPFs, considering factors such as germination rates across various temperature and humidity conditions. Despite the common belief that high humidity is essential for the development of natural enemy pathogens (>90%; Tanada and Kaya, 1993), recent studies suggest that a humidity level as low as 53% could suffice for their germination and host mortality (Hastuti et al., 1999).
-
Tanada and Kaya, 1993Insect Pathology, 2012
-
Hastuti et al., 1999Effects of temperature and humidity on the susceptibility of Paropsis charybdis to Beauveria bassianaProceedings of the New Zealand Plant Protection Conference, 1999
These findings open up possibilities for identifying more virulent isolates in adverse environments. For instance, citrus regions characterized by low macroenvironmental humidity but favorable microenvironmental conditions for EPFs (e.g., due to irrigation practices) could be potential hotspots for identifying highly effective isolates. Surprisingly, among 28 articles related to EPFs in D. citri, none address the isolate’s response to humidity, indicating a significant gap in the existing research.
The assessment of other natural enemies, such as entomopathogenic nematodes (EPNs), remains a crucial area for exploration. Initial observations suggest that Heterorhabditis indica can infect and kill adult D. citri (unpublished data). However, the application of EPNs via foliage spraying has yielded limited results thus far. Hence, a pertinent question arises: how can EPNs be effectively applied for D. citri control? One plausible strategy could involve deploying attractive traps baited with juvenile EPNs. The swift mortality inflicted by these organisms on their hosts makes them an appealing tool for managing disease vectors like D. citri.
The vast diversity of the Mexican citrus agroecosystem poses a challenge for region-specific management of the D. citri-CLas pathosystem (Mora-Aguilera, 2013). Nevertheless, local management strategies applied to small and scattered citrus plantations, including ornamental Rutaceae, have demonstrated efficacy in disease control (Uechi et al., 2019). These successful practices should be considered for integration into CBC programs in Mexico, aiming to promote crop diversification. For instance, the utilization of windbreak barriers, already recommended for mitigating vector and HLB spread (Stelinski, 2019), could be enhanced by selecting barrier trees with nectar-producing potential. This approach would offer dual benefits: serving as physical barriers and attracting natural enemies and pollinators.
-
Uechi et al., 2019Management of huanglongbing (HLB) by an intensive vector and disease control in the surroundings of the orchard, in addition to planting HLB-free trees in Okinawa, JapanJapan Agricultural Research Quarterly: JARQ, 2019
-
Stelinski, 2019Ecological aspects of the vector-borne bacterial disease, citrus greening (Huanglongbing): dispersal and host use by Asian citrus psyllid, Diaphorina citri KuwayamaInsects, 2019
In summary, the effective and sustainable utilization of natural enemies in CBC programs necessitates robust ecological and epidemiological studies. It’s imperative to recognize natural enemies as integral components of complex ecological networks, contributing to intricate ecological-epidemiological interactions. The cultivation and spontaneous vegetation within agroecosystems play pivotal roles as the foundational trophic level, where various organisms, including entomophagous, entomopathogens, phytophagous, and phytopathogens, intersect. From these interactions emerge complex positive and negative feedback loops at each trophic level, ultimately influencing the health of citrus crops. For example, the predator-parasitoid-entomopathogen (P-P-E) complex associated with Toxoptera citricida indirectly promotes the survival of D. citri and thus CLas, by suppressing competition between the two pests for vegetative shoots. Ultimately, the entire D-P-E complex surrounding D. citri and T. citricida contributes positively to overall citrus health (Figure 4) (Mora et al., 2017).
Thumbnail

Figure 4
Phytosanitary trophic system in two citrus species (Persian lime and sweet orange) and the lemon grass M. paniculata, which are differentially infested with two pest-vector species (D. citri and T. citricida) and two pathogens (Citrus tristeza virus and Candidatus Liberibacter asiaticus). Taken from Mora et al. (2017).
Phytosanitary trophic system in two citrus species (Persian lime and sweet orange) and the lemon grass M. paniculata, which are differentially infested with two pest-vector species (D. citri and T. citricida) and two pathogens (Citrus tristeza virus and Candidatus Liberibacter asiaticus). Taken from Mora et al. (2017).
Conclusions
The primary control strategy for D. citri and CLas involves insecticides, along with inundative biological control using T. radiata and some species of EPFs in a complementary manner. Although natural epizootics of fungi in D. citri occur, there is a lack of ecological and epidemiological studies aiding in the rational selection of EPFs isolates. Few studies have been conducted to expand their use endophytically and through self-dissemination.
Predators of D. citri are recognized as one of its main regulators, sometimes even surpassing T. radiata in various parts of the world. However, their extensive diversity has been underutilized in managing the pathosystem, mainly due to chemical control interference and the ecological simplicity of cultivation.
The absence of studies on conservation biological control programs for D. citri is evident, despite repeated mentions of its importance. Implementation of such programs could enhance not only the biological control of D. citri-CLas but also the self-regulation of citrus pests in general. A greater effort and understanding are required from all involved sectors: institutions, government, researchers, technicians, producers, and society at large. The challenge is substantial, but the vast diversity of regions and citrus production systems in Mexico could facilitate what would be a first step towards the concept of integrated biological control: through conservation, classical, and inundative approaches.
Literatura Citada
- Aguilar, RC, Rodríguez, JAL, Gómez, MU y Medel, RR (2014). Evaluación socioeconómica del sistema agroforestal tradicional café-plátano-cítricos en el municipio de Tlapacoyan, Veracruz, México. Tropical and Subtropical Agroecosystems, 17: 315-319. http://www.redalyc.org/pdf/939/93931761024.pdf Links
- Alemán, J, Baños, H y Ravelo, J (2007). Diaphorina citri y la enfermedad huanglongbing: una combinación destructiva para la producción citrícola. Revista de Protección Vegetal, 22: 154-165. Links
- Arshad, M, Ullah, MI, Çağatay, NS, Dikmen, F, Abdullah, A y Afzal, M (2019). Cordia myxa L., a New Host Plant Record for Asian Citrus Psyllid, Diaphorina citri Kuwayama. Southwestern Entomologist, 44: 331-334. https://doi.org/10.3958/059.044.0137 Links
- Aubert, B (1987). Trioza erytreae Del Guercio and Diaphorina citri Kuwayama (Homoptera: Psylloidea), the two vectors of citrus greening disease: biological aspects and possible control strategies. Fruits, 42: 149-162. http://swfrec.ifas.ufl.edu/hlb/database/pdf/00000773.pdf Links
- Avery, PB, Queeley, GL, Faull, J y Simmonds, MSJ (2010). Effect of photoperiod and host distribution on the horizontal transmission of Isaria fumosorosea (Hypocreales: Cordycipitaceae) in greenhouse whitefly assessed using a novel model bioassay. Biocontrol Science and Technology, 20: 1097-1111. https://eprints.bbk.ac.uk/id/eprint/1258/1/1258-nicklin.pdf Links
- Ayala-Zermeño, MA, Gallou, A, Berlanga-Padilla, AM, Serna-Domínguez, MG, Arredondo-Bernal, HC y Montesinos-Matías, R (2015). Characterisation of entomopathogenic fungi used in the biological control programme of Diaphorina citri in Mexico. Biocontrol Science and Technology, 25: 1192-1207. https://doi.org/10.1080/09583157.2015.1041878 Links
- Bamisile, BS, Siddiqui, JA, Akutse, KS, Ramos Aguila, LC y Xu, Y (2021). General limitations to endophytic entomopathogenic fungi use as plant growth promoters, pests and pathogens biocontrol agents. Plants, 10: 1-23. https://doi.org/10.3390/plants10102119 Links
- Brown, SE, Oberheim, AP y Barrett, A (2014). First report of ‘Candidatus Liberibacter asiaticus’ associated with huanglongbing in the weeds Cleome rutidosperma, Pisonia aculeata and Trichostigma octandrum in Jamaica. Journal of Citrus Pathology, 1 (1): 257-258. https://escholarship.org/content/qt8dw9c1f5/qt8dw9c1f5.pdf Links
- Chiu, SC, Aubert, B y Chien, CC (1988). Attempts to establish Tetrastichus radiatus Waterson (Hymenoptera, Chalcidoidea), a primary parasite of Diaphorina citri Kuwayama in Taiwan. En: International Organization of Citrus Virologists Conference Proceedings, 10: 265-268. https://doi.org/10.5070/C520x931xp Links
- Cortez-Madrigal, H, García-González, F, Guzmán-Larralde, A, Acuña-Soto, JA y Otero-Colina, G (2016). Conserving phytophagous arthropods to conserve natural enemies: Asclepias curassavica as the model. Southwestern Entomologist, 41: 681-692. https://doi.org/10.3958/059.041.0311 Links
- Cortez-Madrigal, H y Gutiérrez-Cárdenas, OG (2023). Enhancing biological control: conservation of alternative hosts of natural enemies. Egyptian Journal of Biological Pest Control, 33: 1-13. https://doi.org/10.1186/s41938-023-00675-2 Links
- Cortez-Madrigal, H, Mora-Aguilera, G y Humber, RA (2013). Exploración y aislamiento de hongos entomopatógenos asociados a Diaphorina citri en la Península de Yucatán. En: Mendoza PJ de D, De la Cruz LE, Martínez ME, Osorio OR y Estrada BMA. Tópicos Selectos de Agronomía Tropical 2. Villahermosa, Tab., México: Universidad Juárez Autónoma de Tabasco, pp. 93-105. Links
- Cortez-Mondaca, E, Lugo-Angulo, NE, Pérez-Márquez, J y Apodaca-Sánchez, MÁ (2010). Primer reporte de enemigos naturales y parasitismo sobre Diaphorina citri Kuwayama en Sinaloa, México. Southwestern Entomologist, 35: 113-116. https://doi.org/10.3958/059.035.0113 Links
- Desurmont, GA, Tannières, M, Roche, M, Blanchet, A y Manoukis, NC (2022). Identifying an optimal screen mesh to enable augmentorium-based enhanced biological control of the olive fruit fly Bactrocera oleae (Diptera: Tephritidae) and the mediterranean fruit fly Ceratitis capitata (Diptera: Tephritidae). Journal of Insect Science, 22: 11. https://doi.org/10.1093/jisesa/ieac027 Links
- Damsteegt, VD, Postnikova, EN, Stone, AL, Kuhlmann, M, Wilson, C y Schneider, WL (2010). Murraya paniculata and related species as potential hosts and inoculum reservoirs of ‘Candidatus Liberibacter asiaticus’, causal agent of huanglongbing. Plant Disease, 94 (5): 528-533. https://doi.org/10.1094/PDIS-94-5-0528 Links
- Díaz-Padilla, G, López-Arroyo, JI, Sánchez-Cohen, I, Guajardo-Panes, RA, Mora-Aguilera, G y Quijano-Carranza, JÁ (2014). Áreas de abundancia potencial en México del vector del huanglongbing, Diaphorina citri (Hemiptera: Liviidae). Revista Mexicana de Ciencias Agrícolas, 5 (7): 1137-1153. https://www.scielo.org.mx/scielo.php?pid=S2007-09342014000700001&script=sci_arttext Links
- Diniz, AJF, Garcia, AG, Alves, GR, Reigada, C, Vieira, JM y Parra, JRP (2020). The enemy is outside: Releasing the parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in external sources of HLB inocula to control the Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae). Neotropical Entomology, 49: 250-257. https://doi.org/10.1007/s13744-019-00736-y Links
- Doutt, RL y Nakata, J (1973). The rubus leafhopper and its egg parasitoid: An endemic biotic system useful in grape-pest management. Environmental Entomology, 2: 381-386. https://doi.org/10.1093/ee/2.3.381 Links
- Duran-Vila, N y Bové, JM (2015). Citrus HLB is an emerging disease transmitted by psyllid vectors. Can it be prevented? If not, can it be managed? International Centre for Advanced Mediterranean Agronomic Studies. Watch Lett, 3: 1-9. Links
- Étienne, J, Quilici, S, Marival, D y Franck, A (2001). Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae). Fruits, 56: 307-315. https://fruits.edpsciences.org/articles/fruits/pdf/2001/05/quilici.pdf Links
- Flores, D y Ciomperlik, M (2017). Biological control using the ectoparasitoid, Tamarixia radiata, against the Asian citrus psyllid, Diaphorina citri, in the Lower Rio Grande Valley of Texas. Southwestern Entomologist, 42: 49-59. https://doi.org/10.3958/059.042.0105 Links
- Galdeano, DM, De Souza Pacheco, I, Alves, GR, Granato, LM, Rashidi, M y Machado, MA (2020). Friend or foe? Relationship between ‘Candidatus Liberibacter asiaticus’ and Diaphorina citri. Tropical Plant Pathology, 45: 559-571. https://doi.org/10.1007/s40858-020-00375-4 Links
- Gallou, A, Serna-Domínguez, MG, Berlanga-Padilla, AM, Ayala-Zermeño, MA, Mellín-Rosas, MA y Arredondo-Bernal, HC (2016). Species clarification of Isaria isolates used as biocontrol agents against Diaphorina citri (Hemiptera: Liviidae) in Mexico. Fungal Biology, 120: 414-423. https://doi.org/10.1016/j.funbio.2015.11.009 Links
- García-Ávila, CJ, Trujillo-Arriaga, FJ, Quezada-Salinas, A, Ruiz-Galván, I, Bravo-Pérez, D y Robles-García, PL (2021). Holistic area-wide approach for successfully managing citrus greening (Huanglongbing) in Mexico. En: Hendrichs J, Pereira R y Vreysen MJB. Area-Wide Integrated Pest Management. Boca Ratón, FL: CRC Press, pp. 33-49. Links
- Godoy-Ceja, CA y Cortez-Madrigal, H (2018). Potencial de Asclepias curassavica L. (Apocynaceae) en el control biológico de plagas. Revista Mexicana de Ciencias Agrícolas, 9: 303-315. https://doi.org/10.29312/remexca.v9i2.1073 Links
- Granados-Ramírez, GR y Hernández-Hernández, R (2018). Reconversión de cultivos como resultado de la presencia de huanglongbing en Colima, México. Revista Geográfica de América Central, 3: 413-431. http://dx.doi.org/10.15359/rgac.61-3.21 Links
- Gurr, GM y Wratten, SD (1999). Forum ‘Integrated biological control’: A proposal for enhancing success in biological control. International Journal of Pest Management, 45: 81-84. https://doi.org/10.1080/096708799227851 Links
- Gurr, GM y You, M (2016). Conservation biological control of pests in the molecular era: New opportunities to address old constraints. Frontiers in Plant Science, 6: 1255. https://doi.org/10.3389/fpls.2015.01255 Links
- Hall, DG, Hentz, MG, Meyer, JM, Kriss, AB, Gottwald, TR y Boucias, DG (2012). Observations on the entomopathogenic fungus Hirsutella citriformis attacking adult Diaphorina citri (Hemiptera: Psyllidae) in a managed citrus grove. BioControl, 57: 663-675. https://doi.org/10.1007/s10526-012-9448-0 Links
- Halbert, SE y Manjunath, KL (2004). Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in Florida. Florida Entomologist, 87: 330-353. https://doi.org/10.1653/0015-4040(2004)0870330:ACPSPA2.0.CO;2 Links
- Hastuti, BS, Glare, TR y Chapman, RB (1999). Effects of temperature and humidity on the susceptibility of Paropsis charybdis to Beauveria bassiana. En: Proceedings of the New Zealand Plant Protection Conference, 52: 103-107. Links
- Ibanez, V, López-García, A, Carbonell Caballero, J, Alonso, R, Terol, J, Dopazo, J y Talón, M (2015). El origen de las especies cultivadas de cítricos. Levante Agrícola: Revista Internacional de Cítricos, 426: 74-79. http://redivia.gva.es/bitstream/handle/20.500.11939/4176/2015_Iba%C3%B1ez_El%20Origen.pdf?sequence=1&isAllowed=y Links
- Inoue, H, Ohnishi, J, Ito, T, Tomimura, K, Miyata, S, Iwanami, T y Ashihara, W (2009). Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Annals of Applied Biology, 155: 29-36. https://doi.org/10.1111/j.1744-7348.2009.00317.x Links
- Irvin, NA y Hoddle, MS (2021). The effects of floral nectar, extrafloral nectar and hemipteran honeydew on the fitness of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of Diaphorina citri. Biological Control, 163: 104753. https://doi.org/10.1016/j.biocontrol.2021.104753 Links
- Irvin, NA, Pierce, C y Hoddle, MS (2021). Evaluating the potential of flowering plants for enhancing predatory hoverflies (Syrphidae) for biological control of Diaphorina citri (Liviidae) in California. Biological Control, 157: 104574. https://doi.org/10.1016/j.biocontrol.2021.104574 Links
- Jacas, JA y Urbaneja, A (2010). Biological control in citrus in Spain: from classical to conservation biological control. En: Ciancio A y Mukerji KG. Integrated Management of Arthropod Pests and Insect Borne Diseases. London: Springer, pp. 61-70. Links
- Juárez-Jaimes, V, Alvarado-Cárdenas, LO y Villaseñor, JL (2007). La familia Apocynaceae sensu lato en México: diversidad y distribución. Revista Mexicana de Biodiversidad, 78: 459-482. https://www.scielo.org.mx/pdf/rmbiodiv/v78n2/v78n2a20.pdf Links
- Kehrli, P, Lehmann, M y Bacher, S (2005). Mass-emergence devices: a biocontrol technique for conservation and augmentation of parasitoids. Biological Control, 32: 191-199. https://doi.org/10.1016/j.biocontrol.2004.09.012 Links
- Kondo, T, González, G, Tauber, C, Sarmiento, YCG, Mondragon, AFV y Forero, D (2015). A checklist of natural enemies of Diaphorina citri Kuwayama (Hemiptera: Liviidae) in the department of Valle del Cauca, Colombia and the world. Insecta Mundi, 457: 1-14. https://journals.flvc.org/mundi/article/view/0457 Links
- Leong, SS, Leong, SCT y Beattie, GAC (2020). Incidence and spread of Huanglongbing (HLB) or citrus greening disease in relation to the distribution and fluctuation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) population in a citrus orchard in Sarawak, Malaysia. Serangga, 24-42. http://journalarticle.ukm.my/16586/1/37916-134204-1-PB.pdf Links
- Lezama-Gutiérrez, R, Molina-Ochoa, J, Chávez-Flores, O, Ángel-Sahagún, CA, Skoda, SR, Reyes-Martínez, G y Foster, JE (2012). Use of the entomopathogenic fungi Metarhizium anisopliae, Cordyceps bassiana and Isaria fumosorosea to control Diaphorina citri (Hemiptera: Psyllidae) in Persian lime under field conditions. International Journal of Tropical Insect Science, 32: 39-44. http://digitalcommons.unl.edu/entomologyfacpub/362 Links
- Lira, R (2004). El género Sycidium (Cucurbitaceae, Zanonioideae, Zanonieae, Sicydiinae) en México. Acta Botánica Mexicana, 68: 39-64. https://inecol.repositorioinstitucional.mx/jspui/bitstream/1005/387/1/978-Texto%20del%20art%C3%ADculo-2771-1-10-20160628.pdf Links
- Liu, YH y Tsai, JH (2000). Effects of temperature on biology and life table parameters of the Asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Annals of Applied Biology, 137: 201-206. https://swfrec.ifas.ufl.edu/hlb/database/pdf/Liu%20and%20Tsai%202000.pdf Links
- Lozano-Contreras, MGL y Jasso-Argumedo, J (2012). Identificación de enemigos naturales de Diaphorina citri Kuwayama (Hemiptera: Psyllidae) en el estado de Yucatán, México. Fitosanidad, 16: 5-11. http://www.redalyc.org/articulo.oa?id=209125190004 Links
- Meyer, JM, Hoy, MA y Boucias, DG (2008). Isolation and characterization of an Isaria fumosorosea isolate infecting the Asian citrus psyllid in Florida. Journal of Invertebrate Pathology, 99(1): 96-102. https://doi.org/10.101016/j.jip.2008.03.007 Links
- Michaud, JP (2004). Natural mortality of Asian citrus psyllid (Homoptera: Psyllidae) in central Florida. Biological Control, 29: 260-269. https://doi.org/10.1016/S1049-9644(03)00161-0 Links
- Monzó, C y Stansly, PA (2015). Thresholds for vector control and compatibility with beneficial fauna in citrus with high incidence of Huanglongbing. Acta Hortic, 1065: 1137-1143. https://swfrec.ifas.ufl.edu/hlb/database/pdf/24_Monzo_15.pdf Links
- Monzó, C y Stansly, PA (2020). Economic value of conservation biological control for management of the Asian citrus psyllid, vector of citrus Huanglongbing disease. Pest Management Science, 76: 1691-1698. https://doi.org/10.1002/ps.5691 Links
- Mora-Aguilera, G (2013). Huanglongbing Candidatus Liberibacter spp. Ficha Ténica N° 78. SENASICA, SADER, México, 28p. Links
- Mora-Aguilera, G, Robles-García, P, López-Arroyo, JI, Flores-Sánchez, J, Acevedo-Sánchez, G, Domínguez-Monge, S y González-Gómez, R (2014). Situación actual y perspectivas del manejo del HLB de los cítricos. Revista Mexicana de Fitopatología, 32(2): 108-119. https://www.scielo.org.mx/pdf/rmfi/v32n2/2007-8080-rmfi-32-02-00108.pdf Links
- Moran, PJ, Patt, JM, Cabanillas, HE, Adamczyk, JL, Jackson, MA, Dunlap, CA y Avery, PB (2011). Localized autoinoculation and dissemination of Isaria fumosorosea for control of the Asian citrus psyllid in South Texas. Subtropical Plant Science, 63: 23-35. https://pubag.nal.usda.gov/download/54210/pd Links
- Nava, DE, Torres, MLG, Rodrigues, MDL, Bento, JMS y Parra, JRP (2007). Biology of Diaphorina citri (Hem., Psyllidae) on different hosts and at different temperatures. Journal of Applied Entomology, 131(9-10): 709-715. https://doi.org/10.1111/j.1439-0418.2007.01230.x Links
- Padulla, LFL y Alves, SB (2021). Suscetibilidade de ninfas de Diaphorina citri a fungos entomopatogênicos. Arquivos do Instituto Biológico, 76: 297-302. https://doi.org/10.1590/1808-1657v76p2972009 Links
- Parra, JRP, Alves, GR, Diniz, AJF y Vieira, JM (2016). Tamarixia radiata (Hymenoptera: Eulophidae) × Diaphorina citri (Hemiptera: Liviidae): mass rearing and potential use of the parasitoid in Brazil. Journal of Integrated Pest Management, 7: 1-11. https://doi.org/10.1093/jipm/pmw003 Links
- Parra, JRP y Coelho, A (2019). Applied biological control in Brazil: from laboratory assays to field application. Journal of Insect Science, 19: 1-6. https://doi.org/10.1093/jisesa/iey112 Links
- Patt, JM, Chow, A, Meikle, WG, Gracia, C, Jackson, MA, Flores, D y Damczyk, JJ (2015). Efficacy of an autodisseminator of an entomopathogenic fungus, Isaria fumosorosea, to suppress Asian citrus psyllid, Diaphorina citri, under greenhouse conditions. Biological Control, 88: 37-45. http://dx.doi.org/10.1016/j.biocontrol.2015.04.014 Links
- Pluke, RW, Qureshi, JA y Stansly, PA (2008). Citrus flushing patterns, Diaphorina citri (Hemiptera: Psyllidae) populations and parasitism by Tamarixia radiata (Hymenoptera: Eulophidae) in Puerto Rico. Florida Entomologist, 91: 36-42. https://doi.org/10.1653/0015-4040(2008)0910036:CFPDCH2.0.CO;2 Links
- Qureshi, JA y Stansly, PA (2007). Integrated approaches for managing the Asian citrus psyllid Diaphorina citri (Homoptera: Psyllidae) in Florida. En: Proceedings of the Florida State Horticultural Society 120: 110-115. https://swfrec.ifas.ufl.edu/docs/pdf/entomology/publications/Stansly_Non_Qureshi07.pdf Links
- Qureshi, JA y Stansly, PA (2009). Exclusion techniques reveal significant biotic mortality suffered by Asian citrus psyllid Diaphorina citri (Hemiptera: Psyllidae) populations in Florida citrus. Biological Control, 50: 129-136. https://doi.org/10.1016/j.biocontrol.2009.04.001 Links
- Qureshi, JA, Rogers, ME, Hall, DG y Stansly, PA (2009). Incidence of invasive Diaphorina citri (Hemiptera: Psyllidae) and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrus. Journal of Economic Entomology, 102: 247-256. https://pubag.nal.usda.gov/download/28837/pdf Links
- Rosales-Martínez, V, Martínez-Dávila, JP, Osorio-Acosta, F, López-Romero, G, Asiaín-Hoyos, A y Estrella-Chulím, N (2018). Aspectos culturales, sociales y productivos para una tipología de cafeticultores. Agricultura, Sociedad y Desarrollo, 15: 47-61. https://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1870-54722018000100047 Links
- Salcedo-Baca, D, Hinojosa, RA, Mora-Aguilera, G, Covarrubias-Gutiérrez, I, DePaolis, JR, Mora-Flores, JS y Cintora-González, CL (2010). Evaluación del Impacto Económico de la Enfermedad de los Cítricos Huanglongbing (HLB) en la Cadena Citrícola Mexicana. Instituto Interamericano de Cooperación para la Agricultura (IICA). 141p. http://cef-profit.com/Resumen%20ejecutivo%20impactos%20HLB.pdf Links
- Saldarriaga-Ausique, JJ, D’Alessandro, CP, Conceschi, MR, Mascarin, GM y Delalibera Júnior, I (2017). Efficacy of entomopathogenic fungi against adult Diaphorina citri from laboratory to field applications. Journal of Pest Science, 90: 947-960. https://doi.org/10.1007/s10340-017-0846-z Links
- SIAP (2022). Anuario estadístico de la producción agrícola. https://nube.siap.gob.mx/cierreagricola/ (consultado 18/08/23). Links
- Skelley, LH y Hoy, MA (2004). A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantine. Biological Control, 29(1): 14-23. https://doi.org/10.1016/S1049-9644(03)00129-4 Links
- Stelinski, LL (2019). Ecological aspects of the vector-borne bacterial disease, citrus greening (Huanglongbing): dispersal and host use by Asian citrus psyllid, Diaphorina citri Kuwayama. Insects, 10: 1-11. http://dx.doi.org/10.3390/insects10070208 Links
- Tanada, Y y Kaya, HK (2012). Insect Pathology. Academic Press. 666p. Links
- Thakuria, D, Chaliha, C, Dutta, P, Sinha, S, Uzir, P, Singh, SB y Singh, D (2023). Citrus Huanglongbing (HLB): Diagnostic and management options. Physiological and Molecular Plant Pathology, 125: 102016. https://doi.org/10.1016/j.pmpp.2023.102016 Links
- Tsai, JH y Liu, YH (2000). Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. Journal of Economic Entomology, 93(6): 1721-1725. https://doi.org/10.1603/0022-0493-93.6.1721 Links
- Uechi, N, Katoh, H, Iwanami, T, Fujikawa, T, Miyata, SI, Yasuda, K y Uesato, T (2019). Management of huanglongbing (HLB) by an intensive vector and disease control in the surroundings of the orchard, in addition to planting HLB-free trees in Okinawa, Japan. Japan Agricultural Research Quarterly: JARQ, 53: 103-108. https://www.jstage.jst.go.jp/article/jarq/53/2/53_103/_pdf Links
- Ukuda-Hosokawa, R, Sadoyama, Y, Kishaba, M, Kuriwada, T, Anbutsu, H y Fukatsu, T (2015). Infection density dynamics of the citrus greening bacterium “Candidatus Liberibacter asiaticus” in field populations of the psyllid Diaphorina citri and its relevance to the efficiency of pathogen transmission to citrus plants. Applied and Environmental Microbiology, 81: 3728-3736. https://doi.org/10.1128/AEM.00707-15 Links
- Wenninger, EJ y Hall, DG (2008). Importance of multiple mating to female reproductive output in Diaphorina citri. Physiological Entomology, 33: 316-321. https://doi.org/10.1111/j.1365-3032.2008.00633.x Links
- Wenninger, EJ, Stelinski, LL y Hall, DG (2008). Behavioral evidence for a female-produced sex attractant in Diaphorina citri. Entomologia Experimentalis et Applicata, 128: 450-459. https://doi.org/10.1111/j.1570-7458.2008.00738.x Links
- Yamada, T, Hamada, M, Floreancig, P y Nakabachi, A (2019). Diaphorin, a polyketide synthesized by an intracellular symbiont of the Asian citrus psyllid, is potentially harmful for biological control agents. PLoS One, 14: e0216319. https://doi.org/10.1371/journal.pone.0216319 Links
- Yang, Y, Huang, M, Beattie, GA, Xia, Y, Ouyang, G y Xiong, J (2006). Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: a status report for China. International Journal of Pest Management, 52: 343-352. https://doi.org/10.1080/09670870600872994 Links
- Yzquierdo-Alvarez, ME, Rincón-Ramírez, JA, Loeza-Kuk, E, López-Olguín, JF, Aceves-Navarro, LA y Ortiz-García, CF (2021). Fluctuación espacio-temporal de Diaphorina citri Kuwayama (Hemiptera: Liviidae) en limón persa (Citrus latifolia) en la zona citrícola de Huimanguillo, Tabasco. Acta Zoológica Mexicana, 37: 1-14. https://doi.org/10.21829/azm.2021.371237 Links