










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)  Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Accesos
Accesos
Links relacionados
 Similares en SciELO
Similares en SciELO
Compartir

 Permalink
PermalinkCiencia forestal en México
versión impresa ISSN 1405-3586
Rev. Cien. For. Mex vol.34 no.106 México jul./dic. 2009
Artículo científico
Productividad foliar y radicular en ecosistemas forestales del Noreste de México
José de Jesús Návar–Cháidez1 y Enrique Jurado–Ybarra2
1 Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional (CIIDIR)–IPN, Unidad Durango. Correo–e: jnavar@ipn.mx.
2 Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León.
Fecha de recepción: 12 de marzo de 2008.
Fecha de aceptación: 19 de octubre de 2009.
RESUMEN
La productividad primaria es clave en el entendimiento de los almacenes y flujos de varios elementos biogeoquímicos. En 16 comunidades forestales del noreste de México se midió la caída de hojas y ramas desde enero a junio de 2004; para ello se utilizaron 75 trampas de 1 × 1 m suspendidas sobre el suelo, con tres repeticiones al menos. Se midió la producción de raíces por medio de excavaciones con cilindros, de donde se extrajeron las raíces del suelo. Los resultados mostraron una gran variación temporal y espacial en los parámetros de interés. La productividad de hojarasca promedio (± intervalo de confianza α = 0.05) fue de 3.79 Mg ha–1 a–1 (± 1.43); destacan las comunidades de Cupressus spp. (9.25 ± 2.58), el matorral espinoso tamaulipeco en el derramadero (7.23 ± 2.09) y una plantación de Pinus pseudostrobus (5.27 ± 0.93), mientras que el resto de los bosques tuvieron valores inferiores a 4.05 Mg ha–1 año–1. Las variaciones registradas se asocian con la cantidad de biomasa aérea presente. Los valores correspondientes a la hojarasca siguieron una tendencia estacional contrastante entre comunidades subtropicales y templadas; se definió que, cuantitativamente, es tan importante como la productividad maderable de muchos ecosistemas, por lo que no se debe obviar en los balances de masas y energía.
Palabras clave: Balance de masas, biomasa aérea, comunidades forestales templadas, matorral espinoso tamaulipeco, raíces finas, variaciones espacio–temporales.
ABSTRACT
Primary productivity is a key to understand stocks and fluxes of several biogeochemical elements. The aim of this research project was to measure leaf and branch fall in 75 1 × 1m traps, suspended by ropes with at least three replicates in each of 16 plant communities of northeastern Mexico. Root production was measured during excavations using metal cylinders where roots were extracted. The results showed a great deal of variation on the measured parameters within and between plant communities. Average branch and leaf fall (± confidence intervals α = 0.05) was 3.79 Mg ha–1 a–1 (± 1.43), where highest productivity was recorded on sites reforested with Cupressus spp (9.25 ± 2.58), Tamaulipan thornscrub located in an intermittent wetlands (7.23 ± 2.09) and a Pinus pseudostrobus reforestation site (5.27 ± 0.93) and the remaining of the plant communities recorded productivities of less than 4.05 Mg ha–1 a–1. There was some evidence that the variation in productivity was associated to total standing biomass. Branch and leaf fall followed a seasonal tendency with contrasting tendencies in subtropical and temperate plant communities. Finally leaf and branch fall are an important productivity component to be considered in any mass or energy balance of ecosystems of northeastern Mexico.
Key words: Mass balance, aerial biomass, temperate forest communities, Tamaulipan thornscrub, fine roots, seasonal spatial variations.
INTRODUCCIÓN
Aproximadamente 90% de la biomasa acumulada en la tierra se localiza en los bosques en forma de fustes, ramas, hojas, raíces y materia orgánica (Raeb et al., 1996). Se estima que su producción anual a nivel mundial oscila de 50 a 75 × 109 Mg. Los componentes de productividad se clasifican en materia orgánica seca sobre el suelo (follaje, ramas, fustes, etc.) y la que se localiza por debajo de éste (raíces gruesas y finas, etc.).
Los estudios de hojarasca, generalmente, comprenden a las hojas, ramas, flores y frutos que caen de manera natural y se han utilizado como un índice que permite calcular la productividad primaria neta de una comunidad forestal. Dicho material es un componente cuantitativo y funcional del ecosistema que desempeña un papel importante en el ambiente físico y químico; su descomposición integra elementos de fertilidad y toxicidad al sustrato; intercepta la luz y sirve para regular los gradientes térmicos entre el suelo y la atmósfera; aumenta la captación de la lluvia, pero reduce su evaporación, por lo tanto, sirve de barrera física para muchos procesos y elementos (Facelli y Pickett, 2008), los cuales varían en el espacio y en el tiempo, porque la cantidad presente difiere en función de los sitios y momentos evaluados; ya que es determinada por la tasa de producción, acumulación y descomposición (Couteaux et al., 1995).
El aporte de nutrimentos de la hojarasca de las especies estructurales es muy relevante, como la principal fuente de recursos vitales para el suelo forestal (Vitousek y Sanford, 1986) y representa alrededor del 80% del total de elementos nutritivos que regresan al suelo por los detritos del árbol. La cantidad y naturaleza de la hojarasca tienen una importante relación con la formación del suelo y el mantenimiento de su fertilidad, de ahí que tanto la cuantificación de su producción como su composición sean importantes para comprender los ciclos de elementos en los bosques (Rai y Proctor, 1986; Couteaux et al., 1995; Facelly y Pickett, 2008).
La productividad de la hojarasca es dinámica, pues los cambios climáticos, sucesionales, la edad de las plantas y las características del micrositio, son algunos de los factores que juegan un papel importante en esas variaciones espacio – temporales. Durante las sucesiones terrestres, el incremento en biomasa no fotosintética es muy significativo, por lo que, la caída de hojarasca no refleja la productividad; pero el proceso se invierte, y entonces, llega a constituir hasta un tercio del total de ésta (Puig y Bracho, 1987), a medida que avanzan y la comunidad llega a un estado de equilibrio o estabilidad.
En México se han realizado trabajos de producción de hojarasca, principalmente, en bosques tropicales húmedos y en bosques tropicales deciduos, como el de Álvarez et al. (1992). En este contexto, controles espacio – temporales también han sido estudiados de manera parcial. La producción primaria neta es menor cuando la altitud es mayor en bosques subtropicales deciduos (Martínez–Yrízar et al., 1996). Pavón y Briones (2000) indican que la producción de hojarasca depende de la cantidad de lluvia que reciben las plantas durante la fase de crecimiento. Si se considera que la materia orgánica y los nutrimentos de la vegetación se reincorporan al suelo por el aporte de material foliar, sus resultados apoyan la teoría de que los desiertos son ecosistemas controlados por el agua. De acuerdo con los datos obtenidos en el trabajo de los huertos familiares de Sahcabá y Hocabá, Yucatán (Montañez, 1998), hay relación entre las especies que proporcionan más cantidad de biomasa, vía hojarasca, con aquellas que determinan la estructura del sistema.
Existen experiencias de investigación sobre productividad radicular llevados a cabo con excavaciones, minirrizotrones y observaciones directas (Vogt et al., 1997). La controversia para estimar la dinámica de las raíces finas se asocia con la de la producción y cómo diferentes recursos abióticos cambian dichos parámetros. Una de las razones es que los árboles tienen asignaciones muy variables de la fotosíntesis para estas estructuras (con una variación del 4 – 69% del total del carbono de la planta fijado por año), lo que puede afectar significativamente a los procesos ecológicos. En contraste, con la variabilidad más o menos baja de la productividad de las raíces bajo diferentes condiciones, su producción y turno pueden ser muy sensibles al ambiente y es una forma de respuesta de las plantas que se ajustan a un ambiente cambiante.
En México, y en particular en el noreste del país, existe poca información sobre la productividad foliar y radicular de los diferentes tipos de vegetación que existen. Razón por la cual, el estudio se enfoca en:
a) Medir la cantidad de hojarasca y de raíces finas que producen 16 diferentes comunidades forestales
b) Entender como la biomasa aérea total contribuye con la productividad de hojarasca
c) Explicar las variaciones espacio – temporales en la productividad de hojarasca y de raíces.
Las hipótesis planteadas son las siguientes:
1) Las comunidades forestales producen cantidades similares de hojarasca y biomasa radicular
2) La biomasa aérea total no está relacionada con la productividad de hojarasca
3) No existen variaciones espacio – temporales en su productividad.
MATERIALES Y MÉTODOS
Localización del área de estudio
El área de estudio se ubica en el matorral y bosque escuela de la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León (UANL), en los municipios de Linares e Iturbide, respectivamente, del estado de Nuevo León, México.
El matorral – escuela se localiza en las cercanías de la ciudad de Linares, N. L., dentro del ecosistema denominado matorral espinoso tamaulipeco entre los 24° 47' 00" latitud norte y 99° 32' 00" longitud oeste, el cual se distribuye en la Planicie Costera del Golfo Norte. El bioma cubre una superficie de 200,000 km2 en el sur de Estados Unidos y noreste de México; aquí se ve afectado, negativamente, por el cambio de uso del suelo de forma importante por la práctica ancestral roza–tumba–quema (Treviño et al., 1996; Návar–Chaidez, 2008). El lugar se sitúa a 360 msnm y posee un clima cálido, subhúmedo con una precipitación media anual de 745 mm y una desviación estándar de 250 mm, caracterizado en promedio por 80 eventos lluviosos, de los cuales sólo 5% sobrepasan 20 mm de lámina de agua. La temperatura media anual es de 22.3°C (Návar et al., 1999) (Figura 1).

El bosque – escuela se ubica cerca del pueblo de Santa Rosa en el municipio de Iturbide, N. L. a 40 km al suroeste de Linares a 24° 43' 00'' latitud norte y 99° 52' 00" longitud oeste. Tiene una superficie de 550 ha con un intervalo altitudinal de los 1,200 hasta 1,900 m, enclavado en la Sierra Madre Oriental. La precipitación promedio anual del área es de 600 mm con una desviación estándar de 240 mm y se presenta principalmente durante dos periodos (mayo – junio y julio – septiembre), con una estación interestival seca, denominada canícula. Los inviernos son secos, con la presencia de tormentas y ondas frías ocasionales procedentes del norte. La temperatura promedio anual es de 17°C, con valores que pueden llegar hasta 30°C en el verano y bajo 0°C en el invierno (Figura 1).
Se definieron cuatro áreas de estudio, una y tres en el matorral espinoso tamaulipeco (MET) y en la Sierra Madre Oriental, respectivamente. En el MET se establecieron tres sitios: 1) derramadero (matorral alto), 2) valle (matorral mediano) y 3) loma (matorral bajo). En el bosque – escuela otros tres, los cuales se denominaron: a) La Loma (en reforestaciones de Pinus pseudostrobus Lindl. 1, P. pseudostrobus 2, Cupressus spp., P. pinceana Gordon, P. cembroides Zucc. y P. nelsonii Shaw.), b) Quercus (Quercus canbyi Trel. en dos sitios), c) P. pseudostrobus 1 (bosque nativo de P. pseudostrobus con dos sitios en el paraje La Palapa) y d) Las Adjuntas (en reforestaciones con P. cembroides, P. pinceana y P. nelsonii). En total se midió la hojarasca en 16 comunidades forestales, tres en el matorral espinoso tamaulipeco y 13 en la Sierra Madre Oriental.
La biomasa radicular se determinó sólo en 10 de estas comunidades; las tres del MET, las tres del sitio Las Adjuntas (P. pinceana, P. cembroides y P. nelsonii) y cuatro del sitio La Loma (P. pseudostrobus, P. pinceana, P. cembroides y P. nelsonii).
Recolección de hojarasca
En total se colocaron 75 trampas de 1m2, construidas con un marco de madera y malla de plástico, a razón de tres repeticiones en cada parcela de 20 m2; fueron suspendidas en el aire con lazo para evitar el contacto con el suelo y la pérdida de hojarasca por diferentes procesos. Se establecieron en su mayoría entre enero y marzo de 2004 y las recolectas de hojarasca se realizaron, en promedio, cada 15 días.
Todo el material de la trampa se embolsó y etiquetó para trasladarse al laboratorio; donde se separaron las ramas y hojas del resto de los elementos orgánicos compuestos también por flores y frutos. Se pesó la muestra en verde, misma que se secó en la estufa por un periodo de 48 h a 90°C, finalmente, se registró el peso seco.
Biomasa radicular
En la recolecta de las raíces se utilizó un cilindro de 40 cm de largo y 23 cm de diámetro, que se insertó en el suelo empujándolo firmemente con un mazo hasta alcanzar la roca madre, a una profundidad que varió de 20 a 30 cm. A continuación, el suelo se extrajo del cilindro, se pesó y colocó en la estufa a una temperatura de 90°C durante 48 h, se registró el peso seco total. Se separaron las raíces supriores a 2 mm de diámetro y también se pesaron. Las raíces finas se obtuvieron pulverizando el suelo y enseguida se colocó en agua para separarlas por diferencia de densidades con el uso de un tamiz.
Análisis estadístico de los datos
El análisis de varianza completamente al azar se utilizó cuando los tratamientos considerados correspondieron a las diferentes comunidades, con r repeticiones. La biomasa arbórea total aérea se estimó mediante el ajuste de las ecuaciones alométricas citadas por Návar et al. (2004) y Návar (2009). Se ajustó una relación lineal a los promedios de hojarasca y la biomasa aérea total calculada con las ecuaciones alométricas antes mencionadas.
RESULTADOS
La productividad de hojarasca del matorral espinoso tamaulipeco del noreste de México se ilustra en la Figura 2. En algunas fechas, para diversos sitios se presentó una alta variación en la productividad de hojarasca. En general, ésta fue superior entre los sitios que la observada para las fechas de muestreo; sin embargo, en el Derramadero se registró la mayor productividad, le sigue la del Valle y finalmente La Loma. La disponibilidad de agua del suelo y el aporte de la vegetación muestran la misma tendencia. La productividad promedio en estos lugares fue de (± intervalos de confianza, α = 0.05) de 4.34 (± 1.37) Mg ha–1 año–1.

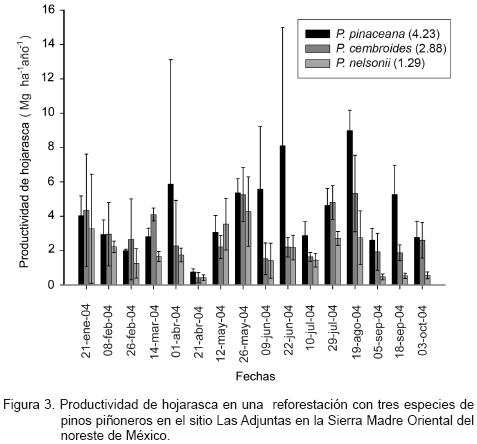
La productividad de hojarasca de las reforestaciones con piñoneros en el sitio Las Adjuntas se muestra en la Figura 3. Los valores más altos se detectaron en los sitios de reforestación con P. pinceana, seguido por el de P. cembroides y por el de P. nelsonii. En ocasiones la productividad fue superior en sitios con P. cembroides, pero su diferencia no es estadísticamente significativa. Los sitios de Las Adjuntas registraron una productividad promedio (± intervalos de confianza, α = 0.05) de 2.85 (± 1.58) Mg ha–1 año–1.
La productividad de hojarasca de las diferentes reforestaciones en La Loma, se presenta en la Figura 4.
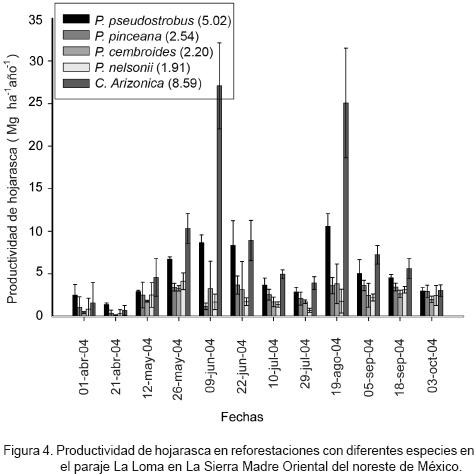
Se observó una distribución más amplia de la productividad de hojarasca, que se explica por las variaciones tan notorias que aportan las reforestaciones de Cupressus spp., que sobresale en algunos periodos. En orden descendente se identificaron las correspondientes a P. pseudostrobus, P. pinceana, P. cembroides y P. nelsonii. Estos sitios de La Loma del bosque–escuela tuvieron valores promedio (± intervalos de confianza, α = 0.05) de 4.12 (± 1.56) Mg ha–1 año–1; aunque Cupressus spp. alcanzó una productividad media de 9.25 (± 2.58) Mg ha–1 año–1.
La productividad de hojarasca varió entre los sitios. Fue menor que lo obtenido cuando se consideraron las diferentes fechas de muestreo, esto indica la importancia del clima en el proceso. Las áreas de bosque nativo de P. pseudostrobus del bosque–escuela tuvieron una productividad promedio (± intervalos de confianza, α = 0.05) de 3.42 (± 0.92) Mg ha–1 año–1.

La variación tuvo un comportamiento similar al bosque nativo de pino (Figura 6). Los sitios de bosque nativo de Quercus spp. del bosque – escuela presentaron una productividad promedio (± intervalos de confianza, α = 0.05) de 3.01 (± 1.39) Mg ha–1 año–1.

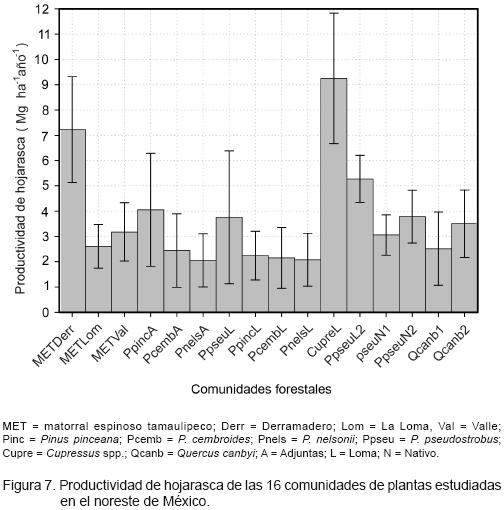
La productividad de hojarasca para cada una de las comunidades se muestra en la Figura 7. Se observa que las reforestaciones de Cupressus spp. resultaron superiores, con excepción del Derramadero en el matorral espinoso tamaulipeco; éste, a su vez, registró un valor más alto que las de pinos piñoneros o que la correspondiente a los bosques nativos de pino o encino. La mayoría de los tipos de vegetación tuvieron valores promedio menores a 4.0 Mg ha–1 año–1, salvo las reforestaciones de Cupressus spp., el Derramadero y el Valle del matorral espinoso tamaulipeco y la de P. pseudostrobus de La Loma. En general, la productividad promedio (± intervalos de confianza, α = 0.05) fue de 3.70 (± 1.43) Mg ha–1 año–1.
Las variaciones de la productividad al nivel de la comunidad forestal fueron explicadas parcialmente por la biomasa aérea total (Figura 8). Se advierte una relación proporcional con una pendiente de 0.064. Es decir, a mayor cantidad de biomasa aérea las comunidades, en promedio, producen más hojarasca. La tasa es de 0.064 Mg ha–1 año–1 por cada Mg ha–1 que existe en el sitio; el valor es muy bajo porque se contrasta con la cantidad total, ante la ausencia de datos precisos de biomasa foliar. Mediciones de compartimentos de biomasa en reforestaciones de Durango (Návar et al., 2004) indican que la parte foliar constituye 15% del total. Es claro que este índice está en función de la edad del árbol. Con dicho valor, un estimador de la caída de hojarasca que dependa de la biomasa foliar sería cercano a 42%. Una relación logarítmica que nace del origen es más aceptable desde el punto de vista fisiológico. Sin embargo, con el modelo lineal se obtuvo un mejor coeficiente de determinación.

Mientras la productividad foliar en el matorral espinoso tamaulipeco de Linares, N. L. llega a su máximo durante la primavera, alcanza su mínimo en las comunidades templadas de piñoneros ubicadas en Iturbide, N. L. Esto se explica, parcialmente, por la interacción fenología de las especies y el clima. Así, la mayoría de las especies del matorral florecen y producen hojas en las etapas primarias de la primavera y otras esperan a las lluvias; en los piñoneros parece que la formación de hojas ocurre en invierno y verano.
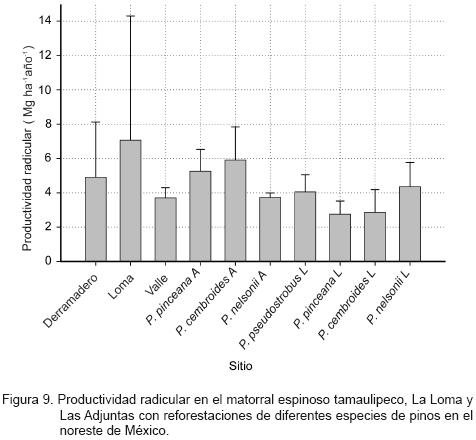
La productividad radicular del matorral espinoso tamaulipeco y en las reforestaciones de Las Adjuntas se presenta en la Figura 9. Se determinó una variación muy amplia entre los sitios y en las comunidades, pero no diferencias estadísticamente significativas. La productividad radicular promedio (± intervalos de confianza, α = 0.05) fue de 5.22 (± 0.99) Mg ha–1 año–1. El paraje La Loma registró un comportamiento similar; aunque sin diferencias estadísticamente significativas, para el caso de las comunidades. Su productividad radicular promedio (± intervalos de confianza, α = 0.05) se calculó en 3.51 (± 1.12) Mg ha–1 año–1. La del paraje Las Adjuntas mostró diferencia estadísticamente significativa en que P. nelsonii tuvo la menor productividad radicular. En esa localidad la productividad radicular promedio (± intervalos de confianza, α = 0.05) fue de 4.96 (± 1.16) Mg ha–1 año–1.
DISCUSIÓN
Los datos de productividad de hojarasca para el matorral que se distribuye en el Derramadero indican un promedio de 7.77 Mg ha–1 año–1 y son similares a la caída de material foliar registrada en un bosque semi deciduo de Brasil, donde se midieron 8.7 Mg ha–1 año–1 (Bretasso–Borges et al., 2003). En los sitios de La Loma del bosque – escuela, localizado en Iturbide, N. L., se determinó una productividad promedio (± intervalos de confianza, α = 0.05) de 4.12 (± 1.56) Mg ha–1 año–1, aunque para Cupressus spp. se estimó un valor promedio de 9.25 (± 2.58) Mg ha–1 año–1, el cual es mayor que la productividad foliar citada para Cupressus spp. en Colombia con 3.7 Mg ha–1 año–1 (Ramírez–Correa et al., 2007). Los sitios de bosque nativo de Quercus spp. del bosque – escuela, Iturbide, N. L. tuvieron una productividad promedio (± intervalos de confianza, α = 0.05) de 3.01 (± 1.39) Mg ha–1 año–1, menor que las 7.9 Mg ha–1 año–1 que se obtuvieron en Colombia para Quercus humboldtii Bonpl. (Ramírez–Correa et al., 2007).
En general las comunidades presentaron una productividad foliar promedio (± intervalos de confianza, α = 0.05) de 3.70 (± 1.43) Mg ha–1 año–1, valor reducido si se le compara con la productividad de hojarasca en bosques tropicales de Brasil que asciende a 10.5 Mg ha–1 año–1 (Figueira et al., 2000). Estas observaciones se ubican en el intervalo consignado para bosques venezolanos, cuyos valores son del orden de 3.9 Mg ha–1 año–1 (Dezzeo y Chacón, 2006).
La productividad radicular promedio (± intervalos de confianza, α = 0.05) de todas las comunidades forestales fue de 4.46 (± 0.84) Mg ha–1 año–1. Estadística que parece alta en contraste con las cifras disponibles en la literatura. Así, en suelos más o menos drenados en Carolina del Sur, la productividad de raíces finas (<3 mm) es de 1.8 Mg ha–1 año–1 (Baker et al., 2001). En bosques de pino y encino, el mismo indicador se calcula en 2.51 y 3.63 Mg ha–1 año–1, respectivamente (Usman et al., 1999).
La productividad de hojarasca y radicular parecen ser muy altas en comparación con la fustal y ramal de las comunidades forestales analizadas. Estas últimas, para el matorral espinoso tamaulipeco varían desde 1.50 Mg ha–1 año–1 (Návar et al., 2001) hasta 2.39 Mg ha–1 año–1 y en el caso de reforestaciones de pinos piñoneros, es menor a 1.00 Mg ha–1 año–1 a una edad de entre 15 y 20 años. Los valores tan contrastantes podrían ser el resultado de varios factores, dentro de los cuales destacan las dimensiones de las plantas, los estadíos sucesionales y la respuesta a los cambios ambientales, entre otros.
La productividad de hojarasca no debe de obviarse en los balances de masas y de energía en comunidades forestales como las aquí estudiadas; ya que es cerca de 2 a 4 veces mayor que la productividad fustal y ramal, lo que probablemente obedece a que las plantas están en las fases intermedias del establecimiento del matorral espinoso tamaulipeco, el cual se dejó de trabajar de forma convencional (roza–tumba–quema–abandono–enmontamiento) a finales de los años 70s. Las reforestaciones del bosque – escuela, por otra parte, cuentan con tan solo 20 años de edad. Es decir, a medida que la comunidad está en la etapa de regeneración, es factible que las productividades se equilibren y, conforme envejecen, la correspondiente a las raíces y de hojarasca aumenta en contraste con la productividad fustal y ramal, para regresar posiblemente al equilibrio, en comunidades maduras.
El medio ambiente desempeña un papel importante en la diferencia de productividades aérea y subterránea. El índice de precipitación anual y evapotranspiración anual va desde 0.62 hasta 0.75, del matorral espinoso tamaulipeco a los bosques templados y reforestaciones de la Sierra Madre Oriental. En este contexto, las plantas utilizan estrategias o adaptaciones para sobrevivir en ambientes deficitarios de agua, en respuesta a los cambios ambientales. Por ejemplo, los índices de biomasa radicular / biomasa aérea son altos, como ha sido notado para chaparrales de California (Kummerow et al., 2004).
La profundidad del suelo que ocupan los sistemas radiculares es variable también entre comunidades áridas del Desierto Chihuahuense y es una consecuencia de las condiciones del ambiente (Zou et al., 2005). Altos potenciales del agua en las plantas (–4.0 MPa) se observan durante sequías severas en especies del matorral espinoso tamaulipeco (González–Rodríguez y Cantú–Silva, 2001). Návar y Bryan (1990) propusieron que la escorrentía fustal podría ser un mecanismo de adaptación de los arbustos a los ambientes áridos.
Es notoria la falta de información para entender las variaciones existentes tanto en las comunidades forestales estudiadas, como en los diferentes estadíos sucesionales, preguntas a resolver con trabajos específicos posteriores en estos ecosistemas del noreste de México.
CONCLUSIONES
La productividad de hojarasca y radicular en 16 comunidades forestales del noreste de México presentaron mucha variación espacio – temporal, la cual se explica, parcialmente, por las irregularidades en los valores de la biomasa aérea y, tal vez, por los diferentes ritmos fenológicos que pudieron haber contribuido a entender parte de la dispersión remanente.
Respecto a las fechas de muestreo la variación es originada, al menos en parte, por la interacción clima y fenología de las especies, ya que se observan patrones o tendencias contrastantes en comunidades semi–áridas subtropicales y en comunidades templadas.
REFERENCIAS
Álvarez S., F. J., G. R. Sánchez, I. Sánchez–Gallén y J. A. González–Iturbe. 1992. Métodos para el estudio de la productividad primaria y la descomposición en comunidades terrestres. Cuadernos de Ecología, Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D. F. México. 52 p. [ Links ]
Baker, T. T., W. H. Conner, B. G. Lockaby, J. A. Stanturf and M. K. Burke. 2001. Fine root productivity and dynamics on a forested floodplain in South Carolina. Soil Science Society of America 65: 545–556. [ Links ]
Bretasso–Borges, M. S., D. Prieto–Trueba, F. C. Peral, C. A. R. Tamburi, R. Caseri y R. Berzaín–Iturralde. 2003. Producción de hojarasca em um bosque semideciduo estacional en São Pedro, Potriendaba, São Paulo, Brazil. Revista del Jardín Botânico Nacional 24: 173–176. [ Links ]
Couteaux, M. M, P. Bottner and B. Berg. 1995. Litter decomposition, climate and litter quality. Trends in Ecology and Evolution 10: 63–66. [ Links ]
Dezzeo, N. and N. Chacón. 2006. Litterfall and nutrient input in undisturbed and adjacent fire disturbed forests of the gran sabana, southern Venezuela. Interciencia 31: 894–899. [ Links ]
Facelli, J. M. and S. T. A. Pickett. 2008. Plant litter: its dynamics and effects on plant community. Botanical Review 57: 1–32. [ Links ]
Figueira, A. M. S., C. A. Souza, A. Maia, H. Rocha, H. Freitas, M. Goulden, M. Menton and S. D. Miller. 2000. Litterfall and leaf area measurements before and after logging in Tapajós National Forest, Santarem–Pará, Brasil. AGU Chapman Conference. San Diego, CA. USA. 12 p. [ Links ]
González–Rodríguez, H. e I. Cantú–Silva. 2001. Adaptación a la sequía de plantas arbustivas de matorral espinoso tamaulipeco. Ciencia UANL 4: 454–461. [ Links ]
Kummerow, J., D. Krause and W. Jow 2004. Root systems of chaparral shrubs. Oecologia 29: 163–177. [ Links ]
Martínez–Yrízar, A., J. M. Mass, L. A. Pérez–Jiménez and J. Sarukhán K. 1996. Net primary productivity of a tropical deciduous forest ecosystem in western Mexico. Journal of Tropical Ecology 12: 169–175. [ Links ]
Montañez E., P. 1998. Producción de hojarasca y aporte de nutrimentos en los huertos familiares de Hocabá y Sahcabá, Yucatán, México. Tesis de Maestría. Facultad de Medicina Veterinaria y Zootecnia. Universidad Autónoma de Yucatán. Mérida, Yuc., México. 110 p. [ Links ]
Návar, J. and R. Bryan. 1990. Interception loss and rainfall redistribution by three semi–arid growing shrubs in northeastern Mexico. Journal of Hydrology 115: 51–63. [ Links ]
Návar J., F. Charles and E. Jurado. 1999. Spatial variations of interception losscomponents by Tamaulipan thornscrub in northeastern Mexico. Forest Ecology and Management 124: 231–239. [ Links ]
Návar, J., J. Nájera and E. Jurado. 2001. Preliminary estimates of biomass growth in the Tamaulipan thornscrub of northeastern Mexico. Journal of Arid Environments 47(3): 281–290. [ Links ]
Návar, J., E. Mendez, J. Graciano, V. Dale and B. Parresol. 2004. Biomass equations for shrub species of Tamaulipan thornscrub of northeastern Mexico. Journal of Arid Environments 59(4): 657–674. [ Links ]
Návar–Chaidez, J. J. 2008. Carbon fluxes resulting from land use changes in the Tamaulipan thornscrub of northeastern Mexico. Carbon Balance and Management 3:6 doi:10.1186/1750–0680–3–6. [ Links ]
Návar, J. 2009. Biomass component equations for Latin American species and groups of species. Annals of Forest Science 66: 208–216. [ Links ]
Pavón, N. P. and O. Briones. 2000. Root distribution, standing crop biomass and belowground productivity in a semidesert in Mexico. Plant Ecology 146: 131–136. [ Links ]
Puig, H. y R. Bracho. 1987. El bosque mesófilo de montaña de Tamaulipas. Instituto de Ecología, A. C. México, D. F. México. 186 p. [ Links ]
Ramírez–Correa, J. A., C. M. Zapata–Duque, J. D. León–Peláez y M. I. González–Hernández. 2007. Caída de hojarasca y retorno de nutrientes en bosques montanos andinos de Piedras Blancas, Antioquia, Colombia. Interciencia 32: 303–311. [ Links ]
Raeb, I., U. Asan y O. Grozeb. 1996. Acumulación de CO2 en la parte aérea de la biomasa de los bosques de Turquía y Bulgaria, en las últimas décadas. In: Memorias XII Congreso Forestal Mundial, Natalia, Turquía. pp. 121–128. [ Links ]
Rai, S. N. and J. Proctor. 1986. Ecological studies on four rainforests in Karnataka, India. II. Litterfall. Journal of Ecology 74: 455–463. [ Links ]
Treviño, E., A. Akca, J. Návar, J. Jiménez and O. Aguirre. 1996. Detection of land use change by satellite imagery in the municipality of Linares, Nuevo León, México. Proceedings of the Fifth International Conference on Desert Development. Vol I. Texas Tech University, August 12–17. pp. 236–240. [ Links ]
Usman, S., S. P. Singh and Y. S. Rawat. 1999. Fine root productivity and turnover in two evergreen Central Himalayan Forests. Annals of Botany 84: 87–94. [ Links ]
Vitousek, P. M. and R. L. Sanford. 1986. Nutrient cycling in moist tropical forest. Annual Review Ecology and Systematics 17: 137–167. [ Links ]
Vogt, K., D. Vogt and J. Bloomfield. 1997. Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level. Plant and Soil 200: 71–89. [ Links ]
Zou, C., S. R. Archer, P. Barnes and C. McMurty. 2005. Soil moisture redistribution as a mechanism of facilitation in savanna tree–shrub clusters. Oecologia 145: 32–40. [ Links ]

