










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)  Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Accesos
Accesos
Links relacionados
 Similares en SciELO
Similares en SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.29 no.2 Ciudad de México ago. 2012
Late Pleistocene mammals from Chivacabé, Huehuetenango, Guatemala
Mamíferos del Pleistoceno tardío de Chivacabé, Huehuetenango, Guatemala
Jim I. Mead1*, Arturo Baez2, Sandra L. Swift1, Jon Lohse3, and Lorena Paiz4
1 Department of Geosciences, and Don L. Sunquist Center of Excellence in Paleontology, East Tennessee State University, Johnson City, TN 37604, USA.
2 Green Houses, College of Agriculture and Life Sciences, University of Arizona, Tucson, AZ 85721, USA.
3 Center for Archaeological Studies, Texas State University–San Marcos, San Marcos, Texas 78666, USA.
4 Universidad del Valle de Guatemala. *mead@etsu.edu
Manuscript received: July 15, 2011.
Corrected manuscript received: December 6, 2011.
Manuscript accepted: December 8, 2011.
ABSTRACT
Few Pleistocene paleontological faunas are published in detail for most of Central America. Probably the richest locality of vertebrates in Central America is at Tomayate, El Salvador, and dates to the early–middle Pleistocene. Literature about late Pleistocene vertebrate species from Guatemala is especially scarce. The purpose of the present paper is to introduce the late Pleistocene mammalian remains from Chivacabé, in the western highlands of Guatemala. The Chivacabé fauna radiocarbon dates to between 15,700 and 12,920 calendar years ago. The specimens recovered from excavations between 1977 and1992 are probably only a small portion of the entire fauna likely to exist under 4 to 5 m of redeposited tephra and valley alluvium. Recovered specimens include at least one individual of Glyptotherium sp., three individuals of Cuvieronius cf. C. hyodon, one individual of Equus sp., and two individuals of Odocoileus cf. O. virginianus. One specimen that originally was thought to represent a deer antler is in fact a hyoid bone of Cuvieronius. Previous reports of 'peccary ' and Eremotherium from Chivacabé are not supported by archived fossils. No faunal specimens exhibit supposed human modification marks purported by previous investigators; all aberrations observed on the bones and teeth can be explained by other taphonomicprocesses. The Chivacabé fauna represents one of the very few late Pleistocene faunas from Guatemala described thus far. A preliminary list of late Pleistocene localities known in Guatemala suggests that detailed studies of these faunas are warranted.
Key words: Mammals, Pleistocene, Central America, Guatemala.
RESUMEN
Se ha publicado poca información paleontológica detallada de la fauna del Pleistoceno para gran parte de Centroamérica. Probablemente la localidad más rica de vertebrados en Centroamérica es Tomayate, en El Salvador, que data del Pleistoceno temprano a medio. La literatura de especies de vertebrados delPleistoceno tardío de Guatemala es especialmente escasa. El propósito de este trabajo es presentar los restos de mamíferos del Pleistoceno tardío de Chivacabé, en las tierras altas occidentales de Guatemala. La edad de radiocarbono de la fauna de Chivacabé data de entre 15,700 y 12,920 años calendario. Los especímenes recobrados de excavaciones entre 1977 y 1992 son probablemente una pequeña porción de toda la fauna que existe bajo 4 a 5 m de tefra redepositada y aluvión. Los especímenes recobrados incluyen un individuo de Glyptotherium sp., tres de Cuvieronius cf. C. hyodon, uno de Equus sp. y dos de Odocoileus cf. O. virginianus. Un individuo que originalmente se pensaba que representaba una cornamenta de venado es de hecho un hueso flotante de Cuvieronius. Reportes previos de "pecaríes" y de Eremotherium de Chivacabé no son sustentados por fósiles archivados. Ningún espécimen de fauna exhibe las supuestas marcas humanas de modificación dadas a entender por investigadores previos; todas las aberraciones observadas en los huesos y en los dientes pueden ser explicadas por otros procesos tafonómicos. La fauna de Chivacabé representa una de las muy pocas faunas del Pleistoceno tardío de Guatemala descritas hasta ahora. Una lista preliminar de localidades del Pleistoceno tardío de Guatemala sugiere que son necesarios estudios detallados de estas faunas.
Palabras clave: mamíferos, Pleistoceno, Centroamérica, Guatemala.
INTRODUCTION
Pleistocene vertebrates of Central America are extremely important as they often represent extinct species that only recently occupied the Panamanian land bridge connection between North America and South America. This land bridge was the avenue and filter for north and south dispersals during the Great American Biotic Interchange (Webb, 1991; Morgan, 2008; Woodburne, 2010). Elevations along this corridor range from sea level up to over 4,000 m in the Sierra de Cuchumantanes in Guatemala. As climate changed through glacial and interglacial regimes where ice sheets waxed and waned at high latitudes, plants and animals of the Central American land bridge were affected.
Many late Pleistocene vertebrate faunas in Central America have been discovered, yet most are biased toward the larger mammalian species, with small mammal, bird, amphibians, and reptiles rarely recovered. Biochronological studies provide a framework in which to document the sequential changes that occurred in both North America and South America (see Woodburne, 2004, 2010) and permit some organization for fossil localities that lack external isotopic age control. As expected, Central American faunas are a mixture of species from both north and south, and it is often difficult to assign these with confidence to a particular biochronological unit.
In North America the arrival of Bison south of 55°N signifies the end to the Irvingtonian and the beginning of the Rancholabrean North American Land Mammal Ages (Bell et al., 2004); the Lujanian is the corresponding latest Pleistocene land mammal age in South America. Both the Rancholabrean and Lujanian are utilized to represent late Pleistocene faunas of Central America. Many diagnostic taxa of the North American late Pleistocene, such as Bison, Mammuthus, and Euceratherium, occur no further south than Central America; while some species of rodents, opossums, armadillos, tree sloths, and toxondonts, for example, do not occur significantly north of Central America (Marshall et al., 1982; Webb, 1991; Ferrusquía–Villafranca et al., 2010). To date little is understood about the Central American Pleistocene, yet it is clear that the region holds great importance to the overall understanding of changing faunal communities in the Western Hemisphere.
Overviews and discussions of important sites and specimens exist of the late Pleistocene faunas of Central America and southernmost Mexico: Costa Rica (Gómez, 1986; Lucas et al., 1997; Laurito–Mora, 2003), El Salvador (Webb and Perrigo, 1984; Cisneros, 2005, 2008), Honduras (Jackson and Fernandez, 2005; Lucas, 2008b), Mexico (Arroyo–Cabrales et al., 1997; Arroyo–Cabrales and Polaco, 2003; Ceballos et al., 2010; Ferrusquía–Villafranca et al., 2010), Nicaragua (Lucas et al., 2008), and Panama (Gazin, 1956). Records of late Pleistocene species from Guatemala are scarce in the literature (Mook, 1959; Woodburne, 1968; Ericastilla–Godoy and García, 1994). Probably the richest locality of vertebrates in Central America is at Tomayate, El Salvador, and dates to the early–middle Pleistocene (Cisneros, 2005). With the exception of southernmost Mexico (see references in Ferrusquía–Villafranca et al., 2010), few details about Pleistocene vertebrates are published for most of the rest of Central America. The purpose of the present paper is to document the mammalian remains from the late Pleistocene locality at Chivacabé (originally labeled the Mastodonte de Villatoro), located in the western highlands of Guatemala (Figure 1).
CHIVACABÉ LOCALITY
Chivacabé is located on the farm of Octavio Villatoro (Aldea de Chivacabé) approximately 6 km west of the town of Huehuetenango (N15° 18' 46.3", W 91° 32' 50.1") at 1896 m (6,220 ft) elevation (Figure 1). The site is situated on and in a deep alluvial terrace of a small stream in Cañon de Chivacabé that drains into the Río Selegua of the Huehuetenango Basin. Six kilometers north of the locality is the high Sierra de Cuchumantanes.
Chivacabé is a previously–recorded faunal locality that contains latest Pleistocene megafaunal remains, which was investigated in January 2009. A fluted lanceolate projectile point was recovered from the surface at this locality (Hayden, 1980), and previous archaeological research (Hayden and Cox, 1978; Ericastilla–Godoy, 1992) sought, with mixed results, to determine whether cultural materials were deposited in association with the faunal remains. Considering the importance of a Paleoindian encampment with associated late Pleistocene fauna in Central America, additional multidisciplinary research was undertaken in 2009 (Lohse and Paiz, 2010). The prime objective of the 2009 research was to determine whether the faunal remains and cultural artifacts present at the on–site museum were associated, or if the two components were deposited independently. Investigations included detailed archaeological excavation of exposed deposits, a geoarchaeological assessment of the depositional context and history, and an appraisal of the taxa present. Radiocarbon dates were determined on sediment samples collected from documented paleosols. Details about the cultural artifacts, geological history, and geochronology are presented in Lohse and Paiz (2010). Conclusions from the 2009 investigations indicate that the faunal deposit (presented here) is not associated with cultural artifacts. Nevertheless, the paleontological assemblage at Chivacabé represents one of the few fully documented late Pleistocene deposits in a poorly known region, and helps to fill in the overall picture for Central American faunal communities during the period immediately preceding the Holocene.
Previous Excavations
The faunal deposit was discovered in 1976 when the landowner Octavio Alvarado Villatoro and his brother dug a water well. From 1977–1978 Herb Alexander and Brian Hayden (from Simon Fraser University, British Columbia, Canada) excavated a block of sediments adjacent to the well and noted the presence of what they at the time identified as mastodon, horse, glyptodont, peccary, and deer. In 1979 John Cox (Simon Fraser University), who had been a participant in earlier field seasons, conducted limited testing in other areas across the alluvial terrace on which the site is located. Many of the faunal elements previously found (most of which were left in situ) and the cultural artifacts appear to have been preliminarily identified and cataloged during the 1979 season. They display catalog markings that include '1979', as do specimens that are known to have been exposed in prior seasons. The in situ faunal elements, located approximately 4.5 m below present surface, have been the focus of local and national interest and are part of a covered display adjacent to a small museum that also exhibits removed skeletal remains. According to Hayden (1980) and Hayden and Cox (1978) it was during these early excavations that a glyptodont scapula was recovered, which showed an apparent puncture hole thought to be caused by human action (refuted below). Hayden and Cox (1978) thought that the sediments were deposited in a lacustrine environment; however, geomorphic studies in 2009 indicate that the skeletal elements were part of an alluvial deposit (Frederick and Yelacic, 2010; Yelacic, 2010). In 1992, Guatemalan archaeologist Sergio A. Ericastilla–Godoy produced a small extension of the excavation at Hayden's site; a few additional faunal remains in the same faunal layer were recovered and are also on display in situ (Ericastilla–Godoy, 1992, 1996). He also reported additional evidence that was interpreted to represent human modification of skeletal remains, such as cut marks on a deer antler, modified proboscidean molars, and various bone fractures that did not appear to be natural (Ericastilla–Godoy, 1996).
Chronology
The actual age of the bone bed was not understood in detail until conclusion of the 2009 geomorphological work by Frederick and Yelacic (2010). Originally it was thought that the faunal unit included human artifacts in association with extinct species at 4.5 m depth. However, recent analyses indicate that these remains were deposited during a period of general environmental instability following a volcanic event and during which large amounts of poorly consolidated tephra were washed down hillsides and accumulated on terraces and in topographic depressions. On the basis of radiocarbon and stratigraphic data recorded by Frederick and Yelacic (2010), the faunal deposit reported here dates at two standard deviations between 15,700–15,100 calendar years before present (Beta–257570; 13,020±60 radiocarbon years BP) and 13,210–12,920 calendar years before present (Beta–257568; 11,160±60 radiocarbon years BP). These dates are derived from bulk soil humates taken from buried paleosols that bracket what Frederick and Yelacic (2010) label Depositional Unit 1, a layer of reworked tephra that contains the faunal remains. This unit is well over two meters in thickness in the excavation block, and extends considerably deeper across much of the Cañon de Chivacabé. The youngest of these dates corresponds with the North American Clovis interval (Waters and Stafford, 2007), and these ages correspond to the Rancholabrean Land Mammal Age of North America (Bell et al., 2004).
CHIVACABÉ LOCAL FAUNA
The Chivacabé local fauna is represented only by large mammals. All specimens are curated by IDAEH (Departamento de Monumentos Prehispánicos y Coloniales del Instituto de Antropología e Historia de Guatemala) and are archived at the locality. All numbers refer to field specimen numbers assigned by H. Alexander and B. Hayden, and possibly J. Cox. The list of vertebrates recovered varies from annual report to annual report, and no publication could be identified specifically about the fossils and how they were identified. Ericastilla–Godoy (1992, 1996) provides the most recent compilation of species from Chivacabé and includes Odocoileus sp. (deer), 'gliptodonte' (glyptodont), Equus (horse), 'mastodonte' (mastodon, referring to a gomphoth–ere), Tayassu tajuco (peccary), and Eremotherium (giant ground sloth). During the 2009 field season, all specimens that remain exposed in the block, as well as a number that had been removed and stored either in the on–site museum or in the Villatoro home were reexamined. Those fossils in situ were left in place and documented as completely as possible. All of the available faunal specimens discovered during the various excavations discussed above were examined.
Many of the skeletal remains were left in situ, and we observed in 2009 that many of these fossils were highly degraded and falling apart. Polyethylene glycol 1500 was used at some point to preserve the bones during one of the previous field seasons. This preservative was decaying and peeling off some of the bones. Apparently the late Dr. Oscar Polaco (part of the 1991 field crew from Mexico who came to assist S. Ericastilla–Godoy) did extensive repair on the bones using a dental plaster. Earlier photos taken by the excavation crews from Simon Fraser University (still available on the internet; http://www.sfu.museum/hola/en/slides/) show that the bones were originally in a good state of preservation. However, many of the same elements exhibit degradation by 2009. As these are an important component to the on–site display, current plans are for the entire in situ assemblage to remain in place. During the course of our field season, we attempted to improve the conserved condition of the assemblage by hand–cleaning the fossils and then applying PaleoBond (Penetrant, 40, 100), Paleo–Poxy, or Butvar 76. Because many of the skeletal elements had to be left in situ, the certainty of identification was greatly reduced.
Most specimens have a 3–part field number assigned by the various researchers. This sequence is made up of a two–digit number that seems to correspond with a field season during which they were exposed or recorded, a unit designation (1 through 12 for the Simon Fraser University excavations or '92' for the Ericastilla–Godoy field season), and what appears to be a serial specimen number within each unit designation. We observed many specimens that had no apparent field number assigned to them, or such numbers have since degraded, and many specimens had '79' as well as either '77' or '78' suggesting that they had been exposed in an earlier field season and then re–cataloged later. When these specimens were critical for our study we would assign them our own field number, first labeling it by the skeletal element (e.g., molar No Number 5).
Glyptodont
North American glyptodonts are distinct and placed in their own genus Glyptotherium, which is closely related to the South American Glyptodon (Gillette and Ray, 1981). Glyptotherium sp. has been recorded from Pleistocene localities in El Salvador, Honduras, and Panama (Gazin, 1956; Webb and Perrigo, 1984). Glyptodon sp. is reported from Guatemala and Costa Rica (Gómez, 1986). Some of the Costa Rican osteoderms, according to Lucas et al. (1997:421), are heavily abraded yet they "clearly do not belong to Glyptotherium"; however, no defining characters were provided for identification by either set of authors. Jackson and Fernandez (2005) and Lucas (2008b) recovered osteoderms from Honduras that are reported as Glyptotherium floridanum. Until a detailed analysis is conducted on glyptodont osteoderms with associated cranial remains that permit accurate identifications, it is not certain that Glyptodon is known from Central America. We assume that the osteoderms recovered from Chivacabé represent Glyptotherium because the genus is found immediately north in Mexico and south in Honduras (Arroyo–Cabrales and Polaco, 2003; Ferrusquía–Villafranca et al., 2010).
The dermal armor of glyptodonts is their most recognizable characteristic. Consequently, the morphology of the osteoderms that make up the carapace has long been recognized as taxonomically significant (Gillette and Ray, 1981). Each osteoderm is polygonal in shape (six–or four–sided) and sutured to its neighbors (see Holmes and Simpson, 1931). The typical osteoderm has a central figure and several peripheral figures symmetrically arranged surrounding the center which creates a rosette pattern (see Hill, 2006). Diagnostic characters used here are slightly modified from Gillette and Ray (1981) and as in Mead et al. (2007). In Glyptotherium texanum, the central rosette is larger than the peripherals, and is convex and slightly raised above the level of the flattened peripherals. In. G arizonae and G. cyclindricum the central figures are relatively smaller than the peripherals, less than 50% of the entire osteoderm diameter, and are generally flat to weakly convex. The central figure in G. floridanum is approximately equal to and larger in size than the peripherals, and is typically raised and weakly concave. The central figure of the osteoderm of G. mexicanum is generally large, never smaller than 50% of the diameter. Although seemingly distinct for all recognized species of Glyptotherium, we acknowledge that there is variation in the rosette size and pattern within and among species. Complicating the issue is the fact that G. cyclindricum and G. mexicanum are less than adequately understood, being known mainly from the type localities (McDonald, 2002). Glyptotherium arizonae and G. texanum are known only from early Pleistocene and earlier faunas, and are not known from late Pleistocene localities (Gillette and Ray, 1981). Clearly much remains to be understood about Central American glyptodonts.
Glyptodont skeletal remains from Chivacabé represent at least one individual of Glyptotherium sp. The following specimens are identified: osteoderm (78–5–A, 78–5–3, 785–10, 78–5–11, 78–6–12, 78–6–13, 78–6–26, 78–7–4, 78–10–1, 78– 10–3, 78–11–2, 78–11–5, 78–11–9, 78–11–10, 78–11–17, 79– 11–39, 79–11–41, 79–11–43, 79–11–66), long bone fragment (79–11–196), ilium fragment (No Number; on display in museum as 'mastodonte'). The greatest width of the individual osteoderms varies from 41.0 to 53.9 mm (n=10) (Figure 2). Width of the rosettes varies from 17.4 to 29.5 mm. Ratios of these two measurements are expressed as the rosette percentage of the entire width; these vary from 35.6% to 59.3%. None of the central figures are raised or convex; this omits G. texanum (known in North America only from pre–late Pleistocene localities) from consideration. The central figures on osteoderms from Chivacabé all have flat to concave structures. These characters would seem to indicate that some of the osteoderms might be identified as G.flori–danum, whereas other specimens would appear to be from G. cyclindricum or G. mexicanum. Because we feel there is only a single individual glyptodont preserved at Chivacabé, we believe that the species cannot be adequately determined with the recovered specimens. The postcranial remains are fragmentary and too poorly preserved to permit adequate identification. Hayden (1980) and Ericastilla–Godoy (1992) state that specimen 78–8–1c/d is a scapula of a glyptodont; however, we identify this specimen as an ilium belonging to Equus (see below).

Gomphothere
Proboscideans known to have inhabited Central America from southern Mexico through to South America during the Pleistocene include elephants (Mammuthus, Elephantidae), gomphotheres (Cuvieronius, Haplomastodon [including Stegomastodon] Gomphotheriidae), and mast–odonts (Mammut, Mammutidae; see among others, Lucas et al. 1997; Arroyo–Cabrales et al., 2002; Montellano–Ballesteros, 2002; Cisneros, 2005, 2008; Lucas 2008c; Lucas et al., 2008; Lucas and Alvarado, 2010). Ferretti (2008) and Prado et al. (2005) reviewed the gomphotheres of South America. Tooth and tusk morphology of skeletal elements in the Chivacabé local fauna indicates that a few individuals of a gomphothere were recovered and left largely in situ. The systematics and phylogeny of gomphotheres is still not fully resolved (e.g., Shoshani, 1996; Ferretti, 2008; Lucas, 2008c; Lucas and Alvarado, 2010).
The following remains were recovered: tusk fragments (78–5–15 [only tip exposed]; 78–6–8, 78–8–15, 78–10–15); mandible fragment (78–4–11); dp/3? (77–5–4); M3 (77–6–13, 77– 6–15, 78–4–12, 78–5–13, 79–11–2, No Number 5); M1–2s (77–5–5, 77–6–18, 77–3–19?, 78–4–13 [in mandible fragment], 78– 10–16, 78–10–17, 78–11–23, 78–11–24, 78–11–26, 79–11–4, 79– 11–152, No Number 1–4); stylohyoidium (78–2–6); atlas (78–5–18); axis (78–6–3); cervical vertebra (78–2–8, 78–6–7); thoracic vertebra (78–3–4, 78–3–13); vertebral fragment (782–8); patella (78–7–9); rib (78–5–26, 78–6–5, 78–6–6, 78–6–19, 78– 6–20, 78–6–21, 79–11–218, 79–11–219); left humerus (78–2–9); right humerus (78–2–4 [sub–adult], 79–11–200, 79– 11–238); ulna (78–2–1, 79–11–170); femur (78–3–1); left femur (78–5–16); right tibia (78–2–5).
The morphology of the juvenile mandible (78–411) fragment illustrates that the lower jaw does not have a sharply downturned symphysis and therefore does not belong to Rhyncotherium (see Lucas and Morgan, 2008). Preservation is not complete but the existing fragment does not show evidence of lower tusk alveoli that would be present in Rhyncotherium, but not in Cuvieronius (Shoshani, 1996; Lucas, 2008a). This observation implies that 78–4–11 belongs to either Cuvieronius or Haplomastodon (see Lucas and Alvarado, 2010).
Four tusk fragments (78–5–15, 78–6–8, 78–8–15, 7810–15) are well preserved. Tusk 78–5–15 had its base broken during hand–excavation of the well that resulted in the initial discovery of the locality. The tip of the tusk has an enamel band that continues the 100 cm length; the broken base has a diameter of 169.3 mm. Tusk fragment 78–10–15 has been repaired (apparently by Dr. Oscar Polaco; Figure 3a). Tusk fragment 78–8–15 has an enamel band (66.7 mm wide at base of tusk) that spirals from the tip down the length of the straight tusk; length of tusk is 680 mm and base diameter is 177.4 mm (Figure 3b). An enamel band (greatest width= 23.7 mm) begins at the tip of the tusk and spirals the length of the fragment for 320 mm, ending at the broken base, which has a diameter of 40.7 mm. Specimen 78–6–8 is too damaged and highly repaired to permit description.

Haplomastodon does not have an enamel band. Shoshani (1996) and Lucas (2008c) state that the enamel band is usually absent on Haplomastodon, but when present, it is highly reduced. Thus, we have identified the three best–preserved tusks containing prominent enamel bands as belonging to Cuvieronius. Based on preserved tusks, at least two individuals of Cuvieronius cf. C. hyodon are present at Chivacabé. Until there is a detailed study of the Pleistocene gomphotheres, we will follow Lucas (2008a) in using the species name C. hyodon (Fischer, 1814) instead of utilizing C. tropicus (Cope, 1884).
Most teeth are isolated specimens; only the M/2, 784–13, is in a fragment of a mandible (Figures 3c, 3d). An alveolus for the M/3 is apparent and suggests that the tooth was in situ but subsequent to or during excavation the molar became isolated. The M/2 is 120.7 mm long, with the width of the protolophid 70.9 mm and the width of the tritolophid 82.7 mm. There is occlusal wear on the first two lophids and slightly apparent on the tritolophid. Specimen 78–4–12 is an unerupted M/3 (no wear on the lophids; Figure 4b) and may well belong adjacent to specimen 78–4–13. The length is 182.5 mm with the width of the tritolophid 83.7 mm; 4.5 lophids are present. The assumption with these two specimens is that they belong to one individual Cuvieronius with a lower M/2 almost in full wear and an M/3 that is unerupted; this would correspond to a subadult individual.
We are not sure which of the many isolated molars might represent an M/1 that could be associated with 784–13. However, there is a heavily worn, small molariform tooth (77–5–4; Figures 4c, 4d) that appears to be deciduous premolar. According to Shoshani (1996, features 27, 23), Cuvieronius does not have premolars P2/2–P4/4 but it does have a dp3. Figures 4c and 4d show what we believe is a dp3 in an extreme state of occlusal wear. The greatest preserved length is 61.8 mm with a width of the metalophid of 38.0 mm.
Five additional M3s were recovered (77–6–13, 776–15, 78–5–13, 79–11–2, No Number 5). All M3s have 4.5 lophs (4 plus a 5th as a buttress; 77–6–15: Figure 4a); four are in a state of full occlusal wear on all lophs, and one has only slight wear on the buttress loph. We assume that the four M3s with full wear are from one fully–adult individual. Specimen No Number 5 must represent a second adult individual in the deposit.
Specimen 79–11–152 is a fragmentary molar of Cuvieronius that was suggested by Hayden and Cox (1978) to possibly show evidence indicating modification by humans. Although the molar is heavily battered, nothing is evident to us to represent human modifications or pathological abnormality. The tooth roots exhibit normal taphonomic rounding and battering as a result of transport in an active alluvial depositional environment.
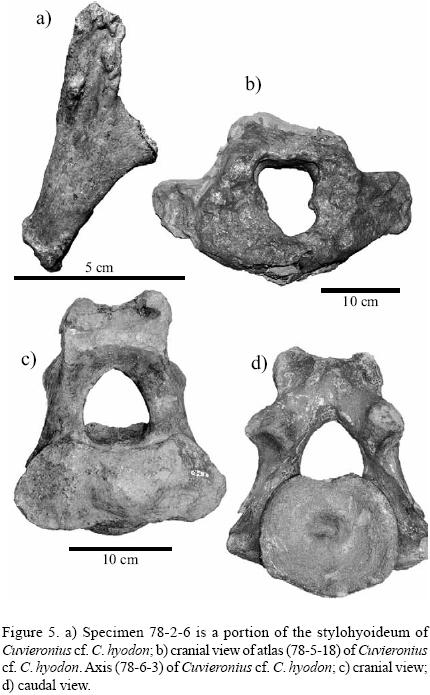
Hayden and Cox (1978) discussed a specimen that they felt represented a deer antler fragment (78–2–6; Figure 5a). The interior structure is not that found in antlers. The cortex is thin and the cross–section is ovoid, not circular as found with antlers at the burr. We have determined that the bone is instead a fragment of a hyoid. Based on its large size it is probably from a gomphothere and may represent Cuvieronius. The hyoid apparatus supports the base of the tongue, pharynx, and larynx, and is constructed of three components: two thyrohyoidea, one basihyoideum, and two stylohyoidea (see Graham, 1986, Shoshani and Marchant, 2001). We believe that specimen 78–2–6 is a portion of the stylohyoideum (Figure 5a). The posterior ramus illustrates the surface that attaches to muscle tissue. The articular surface of the superior ramus articulates with the styloid process of the temporal (squamosal) bone. The ventral shaft of the inferior ramus is absent. Hyoid bones from gomphotheres are rare and have received little attention (Shoshani and Marchant, 2001).
Most postcranial skeletal elements are too poorly preserved to adequately identify. Identification is hampered because most of the bones are in situ and exposed only one or two sides for examination. All postcranial elements are identified as Cuvieronius cf. C. hydon as they are proboscidean elements associated with tusks and molars identified to that taxon.
The atlas (78–5–18; Figure 5b) is complete and remains in situ, which hampers analysis. The greatest dorso–ventral length is approximately 210 mm and the greatest medial width is approximately 360 mm. The axis (78–6–3; Figures 5c, 5d) was complete and in situ but we were allowed to remove the bone for detailed preservation and analysis. The greatest width across the cranial articular processes on either side of the odontoid process is 198.8 mm; greatest dorso–ventral height is 204.5 mm; greatest width across centrum caudal surface is 126.6 mm; and greatest width across the posterior articular processes is 145.5 mm (measurements as in Göhlich, 1998).
Most of the limb elements were either highly degraded at the articular ends or had repairs made with plaster making accurate measurements unattainable. The greatest dorso–ventral length of the patella (78–7–9) is 129.9 mm and the greatest width is 89.7 mm (measurements as in Göhlich, 1998).
Equid
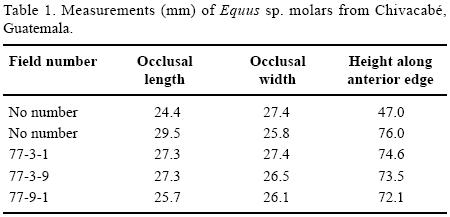
At least one individual horse was recovered (Equus sp.). The following remains were identified: molars (77–3–1, 77–3–9, 77–9–1; two with no numbers; Figure 6a), scapula (79–11–158), fragment of ilium (78–7–9; 78–8–1c/d; Figure 6c), acetabulum and pubis (78–8–2), and tibia (no number). The molars are large (Table 1) and likely represent a larger species of Equus. The scapula is nearly complete with the epiphysis fused representing an adult individual. The length is 27.8 cm from greatest projection of the glenoid fossa to the dorsal edge of the bone; this area is slightly degraded by exposure post–excavation or carnivore gnawing. Based on comparisons with modern Equus skeletons, the scapula 79–11–158 is from the same sized animal as the acetabulum and pubis (78–8–2). Measurement across the acetabulum is 63.7 mm. The acetabulum exhibits a slight puncture mark that is consistent with one made by a carnivore on green bone. The tibia is fragmentary, with heavily worn breakage at the mid–diaphysis. The bone was repaired with plaster by Dr. Oscar Polaco.
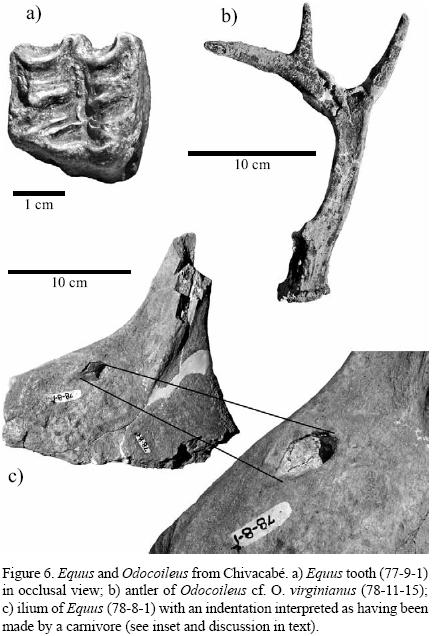
A fragmentary ilium is identified here as belonging to Equus (78–8–1c/d; Figure 6c). Hayden and Cox (1978) and Ericastilla–Godoy (1992) both determined that this bone was a scapula of a glyptodont, 'escapula de gliptodont'. An indentifying mark is found on the bone that Hayden and Cox (1978) and Ericastilla–Godoy (1992) refer to as evidence of an impact depression from a wooden projectile point. Scapulas on live glyptodonts are located beneath (interior to) the dermal armor of the carapace. We believe that the bone is actually an ilium of Equus. The ilial shaft is broken and preserves a green–bone breakage pattern. Close examination of the indentation (Figure 6c), agrees with an indentation made by a canine from a large carnivore. We assume that a carnivore scavenged the carcass. A canine will sometimes leave not only a deep, circular puncture but the curved base of a lower canine will create a lateral, oblique impression adjacent to the depression caused by the posterior portion of the tooth. A series of additional small indentations occur across the iliac flange (lateral face) and may represent tooth 'chatter marks' that indented the bone but did not penetrate the cortex.
Cervid
Two individual–deer (Odocoileus cf. O. virginianus) are documented on the basis of the recovery of two right antlers (78–10–7/8, 78–11–15; Figure 6b). Odocoileus has recurved antlers whereas those of Mazama are straight. Both specimens are naturally–shed antlers and do not represent specimens that were adhering to a skull at death. Antler 7811–15 is nearly complete; diameter of the base at the burr measures antero–posteriorly 40.6 mm and medio–laterally 36.7 mm. Measurement from the burr to the first tine is 43.8 mm, 1st to 2nd tine is 103.8 mm, 2nd to 3rd tine is 60.1 mm, and 3rd to 4th tine 76.4 mm. The second antler (78–10–7/8) is fragmentary but has approximately the same measurements. Hayden and Cox (1978) mentioned a third antler specimen (78–2–6); however, this specimen is identified here as a hyoid (see above discussion on gomphotheres; Figure 5a).
DISCUSSION AND CONCLUSIONS
Central America (that region from the Isthmus of Tehuantepec to Colombia) was the north–south avenue for dispersal of vertebrate species during the Great American Biotic Interchange. The trans–isthmian exchange of vertebrates (especially mammals) from North America and South America began in earnest at about 2.6 Ma and ended sometime after the fourth phase began at about 0.125 Ma (Woodburne, 2010). Many important details about the dispersal events and the taxa involved are still inadequately understood. Included among these details is which in–situ Central American faunas remained after the culmination of the interchange during the latest Pleistocene (Rancholabrean–Lujanian). Although tectonic elements of the land bridge were present well before the onset of the series of dispersal pulses, it appears that climate was the primary controlling factor during the events (Woodburne, 2010).
Habitats during the interglacials of the Pleistocene (as at present) would have been predominantly tropical rainforests across the land bridge, as also occurred during the Miocene and Pliocene (Graham and Dilcher, 1998). Glacially–driven climatic regimes would have resulted in the development of significantly drier landscapes during glacial episodes (Piperno, 2006). Central American lowlands from approximately 20,000 to 10,000 years ago had plant communities that reflected climates which were drier and cooler than present; forest canopies may have been more open than at present and rainfall was decreased by possibly 30% to 50% (Piperno, 2006). The area around Chivacabé is reconstructed to have been mostly undifferentiated thorn woodland, low scrub, and wooded savanna vegetation certainly during the full glacial period (approximately 18,000 to 16,000) (Piperno, 2006; Woodburne, 2010). With the onset of glacial climates, the mosaic of the local plant communities would have changed at all elevations; there is no apparent modern analog for these Pleistocene biomes (Piperno, 2006). During the final phase of the late glacial (~ 10,000 years ago; post–faunal occupation at Chivacabé), the pollen records indicate that the climate quickly warmed substantially, became considerably wetter and highly seasonal, and atmospheric concentrations of CO2 increased.
The Chivacabé fauna dating to between 15,700 and 12,920 calendar years ago was preserved immediately prior to the waning phase of the cooler and drier glacial climate regime. Specimens reported here include: at least one individual of a glyptodont (Glyptotherium sp.) represented by osteoderms and various postcranial long bones; three individuals (one juvenile and likely two adults) of a gomphothere (Cuvieronius cf. C. hyodon) represented by tusk fragments, numerous teeth, vertebrae, and limb elements; one individual of a horse (Equus sp.) represented by teeth and numerous postcranial elements; and two individuals of a deer (Odocoileus cf. O. virginianus) represented by two right, naturally–shed antlers. An additional specimen that originally was thought to represent a deer antler is identified here as a hyoid bone pertaining to Cuvieronius. We did not observe any vertebrate remains that could be assigned to Tayassu or Eremotherium as stated by Hayden and Cox (1978) and Ericastilla–Godoy (1996). Thus, we conclude that these species are not part of the Chivacabé local fauna. In discussing the fossils recovered and archived in the museum, Octavio Villatoro commented that he had never seen any skeletal remains that were thought to represent peccary (although its recovery would not be unusual, Woodburne, 1968). Octavio Villatoro also noted that some previous researchers thought that the giant ground sloth should be found at the locality so it was included on the original list, but he had never seen such remains at Chivacabé. No faunal specimens exhibit human modification, hunting, or butchering marks as suggested by previous investigators; all aberrations observed on the bones and teeth can be explained by physical taphonomic processes.
The Chivacabé fauna represents one of extremely few described late Pleistocene faunas from Guatemala. Ericastilla–Godoy (1996) and Ericastilla–Godoy and García (1994) provided lists of apparent late Pleistocene localities of Guatemala; we have annotated this and added the Chivacabé fauna (Table 2). Late Pleistocene faunas from surrounding countries are much better represented in the literature than those from Guatemala, yet none of the Central American faunas are adequately understood and almost all are heavily biased toward the megamammal taxa. Mammalian microfaunas and herpetofaunas (amphibians and reptiles) are almost unheard of from the late Pleistocene of Central America.

ACKNOWLEDGMENTS
This research was conducted under the authority of Guatemala's Departamento de Monumentos Prehispánicos y Coloniales of the Dirección General del Patrimonio Cultural y Natural–Instituto de Antropología e Historia in the Ministerio de Cultura y Deportes. Sincere appreciation is given to Lic. Juan Carlos Pérez, Jefe of Departamento de Monumentos, who took the time to inspect our project in person, and to Dr. Héctor Escobedo Ayala, Director General del Patrimonio Cultural y Natural, for granting the permission to conduct this research. The authors express their deep gratitude to the National Geographic Society for the support provided by grant number 8485–08 to J. Lohse. Additional funding was made available by the Gault School for Archaeological Research paid for the radiocarbon dates presented here. Sincere thanks goes to Octavio Villatoro and his family for allowing us access to the bone bed and fossils, and his passionate interest in and stewardship of this important site for over 30 years. We appreciate the help in the excavation and geologic stratigraphy by Charles Frederick, Miguel Medina, Luis Méndez, Molly Morgan, Jonathan Morgan, Drew Sitters, Judith Valley, and David Yelacic. JIM and SLS appreciate the many discussions with Michael O. Woodburne, the late Luis Diego Gómez, and Joaquin Arroyo–Cabrales about paleontology and changing environments of Central America. We greatly appreciate the helpful editing suggestions from Victor Manuel Bravo–Cuevas, Eduardo Jiménez–Hidalgo, and Thomas M. Lehman. Susanna Perea–Fox provided the Resumen.
REFERENCES
Arroyo–Cabrales, J., Polaco, O., 2003, Caves and the Pleistocene vertebrate paleontology of Mexico, in Schubert, B.W., Mead, J.I., Graham, R.W. (eds.), Ice Age Cave Faunas of North America: Indiana University Press, 273–291. [ Links ]
Arroyo–Cabrales, J., Polaco, O.J., Laurito–Mora, C., Johnson, E., Alberdi, M.T., Valerio Z., A.L., 1997, The Proboscideans (Mammalia) from Mesoamerica: Quaternary International, 169–170, 17–23. [ Links ]
Arroyo–Cabrales, J., Polaco, O. J., Johnson, E., 2002, La mastofauna del cuaternario tardío en México, in Montellano–Ballestereos, M., Arroyo–Cabrales, J. (eds.), Avances en los Estudios Paleomastozoológicos en México: México, Instituto Nacional de Antropología e Historia, Serie Arqueología, 103–123. [ Links ]
Bell, C.J., Lundelius, E.L., Barnosky, A.D., Graham, R.W., Lindsay, E.H., Ruez, D.R., Semken, H.A., Webb, S.D., Zakrzewski, R.J, 2004, The Blancan, Irvingtonian, and Rancholabrean mammal ages, in Woodburne, M. O. (ed.), Late Cretaceous and Cenozoic Mammals of North America; Biostratigraphy and Geochronology: Columbia University Press, 232–314. [ Links ]
Ceballos, G., Arroyo–Cabrales, J., Ponce, E., 2010, Effects of Pleistocene environmental changes on the distribution and community structure of the mammalian fauna of Mexico: Quaternary Research, 73, 464–473. [ Links ]
Cisneros, J.C., 2005, New Pleistocene vertebrate fauna from El Salvador: Revista Brasileira de Paleontologia, 9, 239–255. [ Links ]
Cisneros, J.C., 2008, The fossil mammals of El Salvador: New Mexico Museum of Natural History and Science Bulletin, 44, 375–380. [ Links ]
Cope, E.D., 1884, The extinct Mammalia of the Valley of Mexico: Proceedings of the American Philosophical Society, 22, 1–21. [ Links ]
Ericastilla–Godoy, S., 1992, Proyecto paleontológico Chivacabe. Report on file with Departamento de Monumentos Prehispánicos y Coloniales, Instituto de Antropología e Historia de Guatemala. [ Links ]
Ericastilla–Godoy, S., 1996, Proyecto paleontológico Chivacabe: Utz'ib, 2, 15–25. [ Links ]
Ericastilla–Godoy, S., García G.,V., 1994, Descubrimientos paleontológicos en el Rosario–Ipala, Oriente de Guatemala: Utz'ib, 1, 1–18. [ Links ]
Ferretti, M.P., 2008, A review of South American proboscideans, in Lucas, S.G., Morgan, G.S., Spielmann, J.A., Prothero, D.R. (eds.), Neogene Mammals: Bulletin of the New Mexico Museum of Natural History and Science, 44, 381–391. [ Links ]
Ferrusquía–Villafranca, I., Arroyo–Cabrales, J., Martínez–Hernández, E., Gama–Castro, J., Ruiz–González, J., Polaco, O.J., Johnson, E., 2010, Pleistocene mammals of Mexico: a critical review of regional chronofaunas, climate change response and biogeogrpahic provinciality: Quaternary International, 217, 53–104. [ Links ]
Fischer, G., 1814, Zoognosia, Tabulis Synopticis Illustrata: Moscow, Typis Nicolai S. Vsevolozsky, 3rd edition. [ Links ]
Frederick, C., Yelacic, D., 2010, Preliminary results of2009 geomorphology investigations at Chivacabé, Huehuetenango, Guatemala, in Lohse, J.C., Paiz. L. (eds.), Final Project Report: Exploring for Clovis Adaptations in Highland Mesoamerica: Washington, D.C, Report of findings submitted to the National Geographic Society. [ Links ]
Gazin, C.L., 1956, Exploration for the remains of giant ground sloths in Panama: Smithsonian Institution Annual Report, 1956, 344–354. [ Links ]
Gillette, D.D., Ray, C.E., 1981, Glyptodonts of North America: Smithsonian Contributions to Paleobiology, 40, 1–255. [ Links ]
Göhlich, U.B., 1998, Elephantoidea (Proboscidea, Mammalia) aus dem Mittel– und Obermiozän der Oberen SiiBwassermolasse Süddeutschlands: Odontologie und Osteologie: Münchner Geowissenschaftliche Abhandlungen. Reihe A. Geologie und Paläontologie, 36, 1–246. [ Links ]
Gómez, L.D., 1986, Vegetación de Costa Rica; Apuntes para una Biogeografía Costarricense; Vegetación y Clima de Costa Rica: San José, Costa Rica, Editorial Universidad Estatal a Distancia, v. 1. [ Links ]
Graham, A., Dilcher, D.L., 1998, Studies in Neotropical paleobotany, XII; A palynoflora from the Pliocene Rio Banco Formation of Costa Rica and the Neogene vegetation of Mesoamerica: American Journal of Botany, 85, 1426–1438. [ Links ]
Graham, R., 1986, Taxonomy of North American Mammoths; Appendix 2, Part 1, in Frison, G.C., Todd, L.C. (eds.), The Colby Mammoth Site – Taphonomy and Archaeology of a Clovis Kill in Northern Wyoming: Albuquerque, University of New Mexico Press, 165–190. [ Links ]
Hayden, B., 1980, A fluted point from the Guatemalan Highlands: Current Anthropology, 21, 702. [ Links ]
Hayden, B., Cox, J., 1978, The Villatoro Mastodon Site: Manuscript on file at the Departamento de Monumentos Prehispánicos y Coloniales, Instituto de Antropología e Historia de Guatemala. [ Links ]
Hill, R.V., 2006, Comparative anatomy and histology of xenarthran osteoderms: Journal of Morphology, 267, 1441–1460. [ Links ]
Holmes, W.W., Simpson, G.G., 1931, Pleistocene exploration and fossil edentates in Florida: Bulletin of the American Museum of Natural History, 59, 383–418. [ Links ]
Jackson, D.R., Fernandez, E., 2005, A small Pleistocene mammalian megafauna from southern Honduras: Bulletin of the Florida Museum of Natural History, 45, 261–269. [ Links ]
Laurito–Mora, C. 2003, Roedores fósiles del Pleistoceneo superior de la localidad La Palmera de San Carlos, Provincia de Alajuela, Costa Rica: Revista Geológica de América Central, 29, 43–52. [ Links ]
Lohse, J. C., Paiz, L., 2010, Final Project Report: Exploring for Clovis Adaptations in Highland Mesoamerica: Washington, D.C., Report of findings submitted to the National Geographic Society, 43 pp. [ Links ]
Lucas, S.G., 2008a, Cuvieronius (Mammalia, Proboscidea) from the Neogene of Florida, in Lucas, S.G., Morgan, G.S., Spielmann, J.A., Prothero, D.R. (eds.), Neogene Mammals: Bulletin of the New Mexico Museum of Natural History and Science, 44, 31–38. [ Links ]
Lucas, S.G., 2008b, Pleistocene mammals from Yeroconte, Honduras, in Lucas, S.G., Morgan, G.S., Spielmann, J.A., Prothero, D.R. (eds.), Neogene Mammals: Bulletin ofthe New Mexico Museum of Natural History and Science, 44, 403–407. [ Links ]
Lucas, S.G., 2008c, Taxonomic nomenclature of Cuvieronius and Haplomastodon, Proboscideans from the Plio–Pleistocene of the New World, in Lucas, S.G., Morgan, G.S., Spielmann, J.A., Prothero, D.R. (eds.), Neogene Mammals: Bulletin of the New Mexico Museum of Natural History and Science, 44, 409–415. [ Links ]
Lucas, S.G., Alvarado, G.E., 2010, Fossil Proboscidea from the upper Cenozoic of Central America: taxonomy, evolutionary and paleobiogeographic significance: Revista Geológica de América Central, 42, 9–42. [ Links ]
Lucas, S.G., Morgan, G., 2008, Taxonomy of Rhynchotherium (Mammalia, Proboscidea) from the Miocene–Pliocene of North America, in Lucas, S.G., Morgan, G.S., Spielmann, J.A., Prothero, D.R. (eds.), Neogene Mammals: Bulletin of the New Mexico Museum of Natural History and Science, 44, 71–87. [ Links ]
Lucas, S.G., Alvarado, J.E., Vega, E., 1997, The Pleistocene mammals of Costa Rica: Journal of Vertebrate Paleontology, 17, 413–427. [ Links ]
Lucas, S.G., Garcia, R., Espinoza, E., Alvarado, G.E., Hurtado de Mendoza, L., Vega, E., 2008, The fossil mammals of Nicaragua, in Lucas, S.G., Morgan, G.S., Spielmann, J.A., Prothero, D.R. (eds.), Neogene Mammals: Bulletin of the New Mexico Museum of Natural History and Science, 44, 417–429. [ Links ]
Marshall, L.G., Webb, S.D., Sepkoski, J.J., Raup, D.M., 1982, Mammalian evolution and the Great American Interchange: Science, 215, 1351–1357. [ Links ]
McDonald, H.G., 2002, Fossil Xenarthra of México: a review, in Montellano–Ballestereos, M., Arroyo–Cabrales, J. (eds.), Avances en los Estudios Paleomastozoológicos en México: México, Instituto Nacional de Antropología e Historia, Serie Arqueología, 227–248. [ Links ]
Mead, J.I., Swift, S.L., White, R.S., McDonald, H.G., Baez, A., 2007, Late Pleistocene (Rancholabrean) glyptodont and pampathere (Xenarthra, Cingulata) from Sonora, Mexico: Revista Mexicana de Ciencias Geológicas, 24(3), 439–449. [ Links ]
Mook, C.C., 1959, A new Pleistocene crocodilian from Guatemala: American Museum Novitates, 1975, 1–6. [ Links ]
Montellano–Ballesteros, M., 2002, New Cuvieronius from the Pleistocene of central Mexico: Journal of Paleontology, 76, 578–583. [ Links ]
Morgan, G.S., 2008, Vertebrate fauna and geochronology of the Great American Biotic Interchange in North America, in Lucas, S.G., Morgan, G.S., Spielmann, J.A., Prothero, D.R. (eds.), Neogene Mammals: Bulletin of the New Mexico Museum of Natural History and Science, 44, 93–140. [ Links ]
Piperno, D.R., 2006, Quaternary environmental history and agricultural impact on vegetation in Central America: Annals of the Missouri Botanical Garden, 93, 274–296. [ Links ]
Prado, J. L., Alberdi, M.T., Azanza, B., Sanchez, B., Frassinetti, D., 2005, The Pleistocene Gomphotheriidae (Proboscidea) from South America: Quaternary International, 126–128, 21–30. [ Links ]
Shoshani, J., 1996, Para– or monophyly of the gomphotheres and their position within Proboscidea, in Shoshani, J., Tassy, P. (eds.), The Proboscidea. Evolution and Palaeoecology of Elephants and their Relatives: Oxford University Press, 149–177. [ Links ]
Shoshani, J., Marchant, G.H., 2001, Hyoid apparatus: a little known complex ofbones and its 'contribution' to proboscidean evolution, in Cavaretta, G, Gioia, P., Mussi, M., Palombo, M. R. (eds.), The World of Elephants: Proceedings of the 1st International Congress, Rome, 16–20 October 2001: Consiglio Nazionale delle Ricerche, 668–675. [ Links ]
Thurmond, J.T., 1973, The Ciudad Real fauna of Pleistocene mammals from the Highlands of Guatemala: Geological Society of America Abstracts with Programs, 5(5), 445. [ Links ]
Waters, M.R., Stafford, T.W., 2007, Redefining the age of Clovis: implications for the peopling of the Americas: Science, 315, 1122–1126. [ Links ]
Webb, S.D., 1991, Ecogeography and the Great American Interchange: Paleobiology, 17, 266–280. [ Links ]
Webb, S.D., Perrigo, S.C., 1984, Late Cenozoic vertebrates from Honduras and El Salvador: Journal of Vertebrate Paleontology, 4, 237–254. [ Links ]
Woodburne, M.O., 1968, A late Pleistocene occurrence of the collared peccary, Dicotyles tajacu, in Guatemala: Journal of Mammalogy, 50, 121–125. [ Links ]
Woodburne, M.O., 2004, Global events and the North American mammalian biochronology, in Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic Mammals of North America. Biostratigraphy and Geochronology: Columbia University Press, 315–343. [ Links ]
Woodburne, M.O., 2010, The Great American Biotic Interchange: dispersals, tectonics, climate, sea level and holding pens: Journal of Mammal Evolution, 17, 245–264. [ Links ]
Yelacic, D.M., 2010, Proyecto arquelógico y paleontológico Chivacabé: a geomorphic and geoarchaeological investigation of late Quaternary environments in northwestern Highland Guatemala: San Marcos, U.S.A., Texas State University–San Marcos, M.S. thesis. [ Links ]

