










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)  Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Accesos
Accesos
Links relacionados
 Similares en SciELO
Similares en SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.29 no.1 Ciudad de México abr. 2012
Gasterópodos y bivalvos cisuralianos (Pérmico inferior) de Otlamalacatla, Hidalgo, México
Cisuralian gastropods and bivalves (Lower Permian) from Otlamalacatla, Hidalgo, Mexico
Sara A. Quiroz–Barroso1*, Francisco Sour–Tovar1, Elena Centeno–García2
1 Museo de Paleontología, Departamento de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510 México D. F., México.
2 Departamento de Geología Regional, Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510 México D. F., México. *saqb@hp.fciencias.unam.mx.
Manuscrito recibido: Abril 12, 2011
Manuscrito corregido recibido: Septiembre 30, 2011
Manuscrito aceptado: Octubre 8, 2011
RESUMEN
En esta contribución se reporta la fauna de gasterópodos y bivalvos provenientes de la Formación Tuzancoa en las cercanías del poblado Otlamalacatla, en el noreste del estado de Hidalgo. Los ejemplares están preservados como moldes imperfectos con cierto grado de deformación, lo que dificulta su estudio y limita la precisión de las determinaciones taxonómicas. A pesar de ello, se advierte la presencia de cinco especies de gasterópodos pertenecientes a las familias Bellerophontidae, Omphalotrochidae, Eotomariidae, Naticopsidae y Soleniscidae, y 20 especies de bivalvos de las familias Nuculidae, Mytilidae, Myalinidae, Pterineidae, Bakevellidae, Euchondriidae, Aviculopectinidae, Deltopectinidae, Chaenocardiidae, Streblochondriidae, Heteropectinidae, Acanthopectinidae, Limidae, Schizodidae, Trigoniidae, Astartidae, Sanguinolitidae y Solenomorphidae, que en su mayoría representan su primer registro para México. Las especies determinadas tienen afinidad con faunas del oeste de Texas, que formó parte de la provincia biogeográfica Grandian durante el Pérmico temprano (Cisuraliano).
Palabras clave: Formación Tuzancoa, gasterópodos, bivalvos, Pérmico, México, provincia Grandian.
ABSTRACT
New fauna of gastropods and bivalves colleted in outcrops of Tuzancoa Formation, near Otlamalacatla town, northeast of Hidalgo state, are described. The specimens are preserved like imperfect moulds with some deformation that limit the precise taxonomic determination. However, it is possible to report the presence of five species of gastropods from the families Bellerophontidae, Omphalotrochidae, Eotomariidae, Naticopsidae and Soleniscidae, and 20 species of pelecypods from the families Nuculidae, Mytilidae, Myalinidae, Pterineidae, Bakevellidae, Euchondriidae, Aviculopectinidae, Deltopectinidae, Chaenocardiidae, Streblochondriidae, Heteropectinidae, Acanthopectinidae, Limidae, Schizodidae, Trigoniidae, Astartidae, Sanguinolitidae and Solenomorphidae; most of these species are reported for the first time in Mexico. The principal affinities of this fauna are found in West Texas and belong to earlier Permian (Cisuralian) of Grandian province.
Key words: Tuzancoa Formation, gastropods, bivalves, Permian, Mexico, Grandian province.
INTRODUCCIÓN
En el norte del estado de Hidalgo se encuentra la región históricamente conocida como anticlinorio de Huayacocotla, que forma parte del cinturón de pliegues y cabalgaduras de la Sierra Madre Oriental. En esta zona se encuentran una serie de cabalgaduras y fallas más jóvenes que permitieron que afloraran las rocas más antiguas de la columna estratigráfica, incluyendo el basamento precámbrico y una sucesión de rocas volcaniclásticas paleozoicas que corresponden a la Formación Tuzancoa, portadoras de una fauna de invertebrados marinos que incluyen fusulí–nidos (Rosales–Lagarde et al., 2005). La sección tipo de esta formación está expuesta en las cercanías del poblado Otlamalacatla. Parte de la fauna de esta sección y otra proveniente de localidades cercanas, ha sido estudiada en contribuciones anteriores (Sour–Tovar et al., 2005), reconociéndose una notable afinidad con faunas norteamericanas del Cisuraliano. En este trabajo se dan a conocer los gasterópodos y bivalvos de la Formación Tuzancoa encontrados hasta el momento y se analizan sus implicaciones estratigráficas, paleoecológicas y paleobiogeográficas.
ANTECEDENTES
Carrillo–Bravo (1965) consideró a las rocas pérmicas que afloran en el norte del estado de Hidalgo como el extremo sur de la Formación Guacamaya (Carrillo–Bravo, 1961) de edad Pérmico temprano, cuya localidad tipo se encuentra en el estado de Tamaulipas y que consiste en una sucesión turbidítica de gran espesor, conformada por lutita, arenisca y un conglomerado de clastos de origen ígneo. Dicho autor interpreta a esta unidad como unflysch orogénico que se desarrolló durante la aproximación de Laurasia (Euramérica) y Gondwana durante la formación de Pangea. Ochoa–Camarillo (1996) reconoció diferencias litológicas en las rocas pérmicas de Hidalgo y las dividió en dos unidades que nombró informalmente formación Tuzancoa, formada por depósitos volcano–sedimentarios con intercalaciones de lentes calcáreos, y la formación Otlamalacatla, conformada por una sucesión de arenisca y lutita. Posteriormente, se observó que estas dos unidades en realidad constituyen una sola unidad, a la que Rosales–Lagarde et al., (2005) denominaron Formación Tuzancoa, designando como sección tipo a la sucesión de rocas de más de 700 m de espesor que afloran a lo largo del río Tlacolula, entre los poblados de Otlamalacatla y Chapula (Figura 1). A diferencia de la Formación Guacamaya, la abundancia de rocas volcánicas y volcaniclásticas y su afinidad geoquímica, evidencian que las rocas de la Formación Tuzancoa representan un arco continental más que unflysch orogénico (Rosales–Lagarde, 2002; Rosales–Lagarde et al., 1997, 2005).

Los fósiles de la Formación Tuzancoa han sido listados en diferentes publicaciones y provienen de diversos afloramientos ubicados en el área Calnali–Tlahuelompa; entre ellos se encuentran especies de microforaminíferos, fusulínidos, braquiópodos, crinoideos y algas, así como fragmentos de corales rugosos, amonites y briozoarios (Carrillo–Bravo, 1965; Pérez–Ramos, 1978; Vachard et al., 1997; Arellano–Gil et al., 1998; Silva–Pineda et al., 2003). Silva–Pineda et al. (2003) mencionan la presencia de bivalvos asignados a los géneros Edmondia, Aviculopinna, y Schizodus y entre los gasterópodos Bellerophon. En los alrededores de Calnali han sido referidas plantas fósiles de edad Pérmica temprana (Silva–Pineda, 1987); otra paleoflora de probable edad pérmica (Leonoardiano) ocurre en estratos continentales que afloran alrededor de San Mateo, ubicados a pocos kilómetros al sur del área de estudio (Weber, 1997).
Para la misma localidad de la que provienen los moluscos que se describen en este trabajo, Sour–Tovar et al. (2005) reportan y describen a los braquiópodos Krotovia sp., Dasysaria sp., Derbyoides cf. D. dunbari, Neospirifer cf. N. amphigyus, Neospirifer sp., Spiriferellina tricosa, IHolosia sp. y al trilobite Anisopyge whitei.
UBICACIÓN GEOGRÁFICA Y ESTRATIGRAFÍA
La localidad de estudio se ubica en el noreste de Hidalgo, entre los 20°47'y 20°48' latitud norte y los 98° 33' y 98°34' longitud oeste, cerca del límite con el estado de Veracruz. Las muestras analizadas provienen de un afloramiento ubicado 200 m al sur de Otlamalacatla, sobre el camino que une este poblado con el de Pemuxco (Figura 1).
Las rocas más antiguas que afloran en el área forman parte del Gneis Huiznopala de edad precámbrica. Suprayaciendo a esta unidad se presenta la sucesión de rocas paleozoicas marinas de edad pérmica correspondiente a la Formación Tuzancoa, constituida, en su base, por arenisca cuarzosa y lutita; seguida por una alternancia de lutita y arenisca volcánicas, las cuales son portadoras de los gasterópodos y bivalvos fósiles que aquí se describen. Por encima, la secuencia cambia a flujos de lava masivos y brechoides de composición andesítica, conglomerado volcánico, toba intercalada con turbidita volcaniclástica (alternancia de lutita y arenisca). La sucesión presenta, en diferentes niveles estratigráficos, flujos de detritos calcáreos que contienen abundantes tallos de crinoideos, corales solitarios y fusulínidos. Las rocas volcaniclásticas de la parte superior contienen lentes de conglomerado, depositados como rellenos de canal, formados de clastos de roca volcánica, granito, gneis y caliza (Rosales–Lagarde, 2002; Rosales–Lagarde et al., 1997, 2005) (Figura 2).
En diferentes puntos al norte y este del poblado de Otlamalacatla, se observa el contacto tectónico de las rocas paleozoicas, que cabalgan a rocas del Jurásico Superior de la Formación Huayacocotla o rocas del Cretácico Inferior de la Formación Pimienta. Estas relaciones tectónicas han sido explicadas como resultado de la deformación originada por la orogenia Laramide, que a lo largo del Cretácico Tardío–Eoceno tardío formó el conjunto de pliegues y cabalgaduras del anticlinorio de Huayacocotla y que afectó tanto a las rocas paleozoicas como al basamento y a la cobertura mesozoica (Ochoa–Camarillo, 1996). Hacia su límite oeste y sur, la Formación Tuzancoa está cubierta discordantemente por una unidad clástica del Jurásico Inferior, que cambia a una sucesión calcárea del Jurásico Superior–Cretácico (Rosales–Lagarde et al., 2005). El mapa geológico del área y la ubicación de los afloramientos de la Formación Tuzancoa pueden consultarse en Rosales–Lagarde et al. (2005).
El área de estudio forma parte del terreno tectónico Sierra Madre (Campa y Coney, 1983) que posteriormente es redefinido como el micro–continente Oaxaquia, propuesto por Ortega–Gutiérrez et al. (1995).
PALEOAUTOECOLOGÍA Y CONSIDERACIONES PALEOAMBIENTALES
En estudios anteriores se ha interpretado que la Formación Tuzancoa se desarrolló en una cuenca ligada a un arco volcánico generado sobre corteza continental y que las rocas fosilíferas representan flujos de origen turbidítico y flujos de detritos calcáreos intercalados dentro de la secuencia volcánica (Rosales–Lagarde, 2002; Rosales–Lagarde et al., 2005). El registro de gasterópodos y bivalvos de Otlamalacatla consiste en formas juveniles y fragmentos de conchas de mayor tamaño asociados con restos de braquiópodos, crinoideos, trilobites, corales rugosos y fusulínidos que indican, independientemente de los sesgos tafonómicos, que durante el Pérmico temprano existieron en el área comunidades con una gran biodiversidad propiciada por la ubicación latitudinal y por el mismo vulcanismo, el cual normalmente influye en el calentamiento del mar y en el aporte de grandes cantidades de nutrientes y de dióxido de carbono (Vermeij, 1995).
La diversidad de gasterópodos y bivalvos de la Formación Tuzancoa se presenta tanto desde un punto de vista taxonómico como ecológico, ya que no solo se refiere al número de especies presentes, sino también a la variedad de características autoecológicas particulares (guilds en el sentido de Bambach, 1983) que tuvieron en vida. En la Tabla 1 se listan los géneros de gasterópodos y bivalvos estudiados y el guild correspondiente.

La talla pequeña de los organismos que caracteriza a la mayoría de los invertebrados encontrados en la Formación Tuzancoa podría relacionarse con una alta salinidad, como sucede con las comunidades que se desarrollaron en la cuenca hipersalina de Landlocked del Pérmico de Europa (Ramsbottom, 1981). En estas comunidades, los moluscos fueron la fauna principal, con gasterópodos de talla muy pequeña y bivalvos de tallas variables, desde las pequeñas especies del género Bakevellia que fueron las formas dominantes, hasta representantes de formas grandes y abundantes de la familia Schizodidae. Estas comunidades presentan también microorganismos como foraminíferos y ostrácodos, pero a diferencia de lo que ocurre en la Formación Tuzancoa, no hay briozoarios ni braquiópodos articulados, que son organismos estenohialinos. De acuerdo a esto, la selección de tamaños y el estado fragmentario de los moluscos estudiados puede ser más bien el resultado de un transporte del material por corrientes de turbidez que removieron depósitos marinos someros hacia zonas más profundas en donde quedaron sepultados, constituyendo una tanatocenosis o asociación de muerte.
BIOESTRATIGRAFÍA Y PALEOBIOGEOGRAFÍA
Las familias de gasterópodos y de bivalvos registradas en la Formación Tuzancoa son características del Paleozoico superior. Las especies determinadas han sido referidas para el Leonardiano de Norteamérica, equivalente al piso Kunguriano de la serie Cisuraliana (Tabla 2). De confirmarse en estudios posteriores la presencia de Lyroschizodus orbicularis en la Formación Tuzancoa (considerada a nivel de cf. en este trabajo) se ampliaría el alcance estratigráfico de esta especie, ya que en la actualidad únicamente ha sido referida para el Roadiano de Texas.
Durante el Paleozoico temprano, el micro–continente Oaxaquia (originalmente descrito como los terrenos Coahuila, Sierra Madre y Oaxaca por Campa y Coney, 1983), formó parte de Gondwana, como lo sugieren las faunas fósiles que contiene (Stewart et al., 1999, y referencias listadas en Centeno–García, 2005). Posteriormente, el micro–continente Oaxaquia se separó de Gondwana y colisionó con Laurasia antes del Misisípico, como lo demuestra la afinidad de las faunas que vivieron en los mares epicontinentales que lo cubrían en ese tiempo, y otras faunas norteamericanas (Quiroz–Barroso et al., 2000; Navarro–Santillán et al., 2002).
Durante el Morrowano–Desmoinesiano (equivalente al Bashkiriano–Moscoviano) se acentuó esta afinidad biogeográfica con faunas de bivalvos del Mesocontinente de los Estados Unidos y con faunas de bivalvos de asociaciones paleoecuatoriales del norte de América del Sur, como la Formación Piauí de Brasil y las faunas andinas del Paleozoico superior de los grupos Tarma y Copacabana de Perú (Chronic, 1953; Quiroz–Barroso y Perrilliat, 1997, 1998; Anelli et al., 2002, 2006), así como las de la región centro–oeste de Argentina, que se diferencian de las faunas de otras cuencas de América del Sur en las que predominan elementos gondwánicos (González, 1997; Pagani, 2000, 2004a, 2004b, 2005, 2006a, 2006b; Sterren 2004, 2005; Lech y Milana, 2006).
Durante el Carbonífero y el Pérmico temprano evolucionó en el este de México (Oaxaquia) un arco continental con una zona de subducción paleo–Pacífica inclinada al este, contemporáneo a la deformación en el cinturón Ouachita (Centeno–García, 2005). Para este tiempo existía ya una conexión geográfica entre los cinturones orogénicos Apalachiano–Ouachita con México, Centroamérica y la región andina (Centeno–García, 2005), que ayuda a explicar la similitud faunística entre las contrapartes norte y sur americanas (Hoover, 1981). La conexión marina de aguas poco profundas entre el oeste de Norteamérica, Oaxaquia y el oeste de Sudamérica desarrolló gradientes térmicos que redujeron la dispersión de muchas faunas.
Dentro del Reino Paleoecuatorial del Pérmico temprano y medio, Shen y Shi (2004) distinguen las provincias Cathaysia y Grandian, esta última definida previamente por Yancey (1975). Las biotas marinas tropicales y subtropicales de la provincia Grandian ocuparon el área de Texas–Nuevo México y Coahuila durante el Pérmico temprano y medio; la formación de arrecifes en el área durante este tiempo demuestra la ocurrencia de climas cálidos y los datos paleomagnéticos colocan un paleoecuador que pasa a través o muy cerca de esta área. Estas biotas de aguas cálidas tienen una alta diversidad taxonómica e incluyen grandes foraminíferos, corales, esponjas, algas, así como abundantes braquiópodos y moluscos ornamentados (Yancey, 1975). En este contexto, los bivalvos y gasterópodos de Otlamalacatla presentan una clara afinidad con las faunas tropicales y subtropicales de la provincia Grandian, específicamente con las referidas para Texas. De manera particular Lyroschizodus orbicularis Newell y Boyd, 1975, Euchondria cooperi Newell y Boyd, 1995 y Leptodesma falcata Boyd y Newell, 2001, tres de las especies con las que se compara parte del material estudiado, han sido descritas exclusivamente para Texas (Tabla 2); la última forma parte de una serie de especies endémicas con una existencia aparentemente muy corta (Boyd y Newell, 2001), por lo que de confirmarse su presencia en Hidalgo permitiría explicar algunos registros aparentemente disjuntos que han sido relacionados con los taxa Lazarus, taxa que desaparecen del registro fósil durante intervalos que pueden ser de diferente magnitud y reaparecen en estratos más jóvenes (Jablonski, 1986).
PALEONTOLOGÍA SISTEMÁTICA
Los ejemplares de este estudio se recolectaron en diferentes temporadas de campo y todos ellos provienen de un lente de lutita de aproximadamente dos metros de espesor, localizado en la parte inferior de la secuencia pérmica descrita por Rosales–Lagarde (2002). Actualmente se encuentran depositados en el Museo de Paleontología de la Facultad de Ciencias, Universidad Nacional Autónoma de México, con número de catálogo FCMP 603–650.
Debido al estado actual de los análisis filogenéticos que han llevado a nuevas interpretaciones de las relaciones entre los taxa superiores de gasterópodos (Ponder y Lindberg, 1997; Kues y Batten, 2001; Wagner y Erwin, 2006) y de bivalvos (Waller, 1978, 1998; Amler, 1999; Carter et al., 2000) sin llegar a una aceptación generalizada, los taxa de gasterópodos descritos en este trabajo se asignan a partir de familia, mientras que los taxa de bivalvos se asignan a partir de superfamilia, utilizando para ello la terminación indicada en el Código Internacional de Nomenclatura Zoológica, artículo 29.2 (International Code of Zoological Nomenclature, 2000). Para los gasterópodos se sigue la clasificación de Bouchet et al. (2005) con excepción de los Naticopsidae en la que se sigue la propuesta de Bandel (2007); para los bivalvos se adopta la clasificación de Bouchet et al. (2010).
Phylum Mollusca
Clase Gastropoda Cuvier, 1797
Familia Bellerophontidae M Coy, 1851
Género Bellerophon Montfort, 1808
Especie tipo. Bellerophon vasulites Montfort, 1808, por designación original, Devónico Medio de Alemania.
?Bellerophon sp.
Figuras 3.1, 3.2
Descripción. Concha de tamaño mediano, de 19 mm de longitud, expandida anteriormente hacia el margen anterior y aplanada de manera abrupta hacia la abertura, por lo que la anchura es casi igual que la longitud; perfil transversal redondeado. Ombligo circular, profundo y relativamente grande. No se observan detalles de la superficie exterior.
Material examinado. Un molde interno imperfecto (FCMP 603).
Discusión. La escasa preservación de este ejemplar único impide asignarlo con certeza a este género.
Familia Omphalotrochidae Knight, 1945
Género Babylonites Yochelson, 1956
Especie tipo. Babylonites carinatus Yochelson, 1956, por designación original. Pérmico medio de Texas, Estados Unidos.
Babylonites sp.
Figuras 3.3, 3.4
Descripción. Concha cónica–globosa de tamaño pequeño, de 5.3 a 10.5 mm de anchura. Sección de la vuelta triangular; ángulo pleural de 89° a 92°, sin variaciones importantes durante el desarrollo ontogenético.
Material examinado. Dos moldes internos imperfectos: (FCMP 604 y FCMP 605).
Discusión. A pesar de lo imperfecto de la conservación del material estudiado, la forma cónica de vuelta triangular es característica del género Babylonites. Este género ha sido descrito para el Pérmico (Sakmariano?–Kunguriano y Wordiano) de Norteamérica. Su presencia en afloramientos de México fue referida por Yochelson (1956, p. 202), quien consideró a Omphalotrochus (?) especie A de Knight (1953, p. 88–89, lám. 24F, figs. 26, 27) del área de El Antimonio, Sonora, como un claro representante de este género; en trabajos posteriores se ha referido para la Formación Plomosas, Chihuahua (Bridges, 1964). La especie tipo Babylonites carinatus ha sido referida para el Pérmico medio de la Formación Los Arcos, en Olinalá, estado de Guerrero (González–Arreola et al., 1994, lám. 2, figs. f y g). Debido a que los ejemplares estudiados no conservan las características externas de la concha, es imposible reconocer su identidad a nivel de especie.
Familia Eotomariidae Wenz, 1938
Subfamilia Eotomariinae Wenz, 1938
Género Glabrocingulum Thomas, 1940
Especie tipo. Glabrocingulum beggi Thomas, 1940, por designación original. Carbonífero inferior de Escocia.
Glabrocingulum cf. G. coronatum Chronic, 1952
Figura 3.5
Descripción. Concha cónico–comprimida, pequeña, de 6.8 mm de anchura; espira baja, de aproximadamente la mitad de la altura total de la concha. Perfil de la vuelta convexo por debajo de la sutura, haciéndose cóncavo hacia la periferia por lo que presenta un aspecto sigmoide. Sólo se conservan las dos últimas vueltas, con líneas de crecimiento finas y arqueadas, desarrolladas cerca de la sutura como ondulaciones radiales. Selenizona limitada por dos carenas.
Material examinado. Molde compuesto de un ejemplar juvenil, incompleto (FCMP 606).
Discusión. El ejemplar de Hidalgo presenta las dimensiones y la ornamentación típica de Glabrocingulum? coronatum Chronic, 1952, p. 118, lám. 3, figs. 1a–3, que ha sido referida para la Formación Kaibab (Sakmariano?–Kunguriano), en Walnut Canyon, Arizona y posiblemente también esté presente en la Formación Hueco (Asseliano–Sakmariano), del oeste de Texas (Chronic, 1952). G. (G) coronatum se distingue de una especie afín, G.(G.) n. sp. del Asseliano de la Formación Laborcita, Sacramento Mountains, Nuevo México (Kues, 1991, p. 225, figs. 3.1 a 3.3), ya que esta última presenta nódulos en su superficie. Debido al estado fragmentario del material estudiado, no es posible su asignación definitiva.
Familia Naticopsidae (Miller, 1889)
Género Naticopsis M'Coy, 1844
Especie tipo. Naticopsis phillipsii M'Coy, 1844, por designación subsecuente de Meek y Worthen, 1866. Carbonífero inferior de Irlanda.
?Naticopsis sp.
Figura 3.6
Descripción. Se observa exclusivamente la espira baja, que sobresale ligeramente del fragmento de una concha juvenil de contorno globular. No se observan detalles de la ornamentación.
Material examinado. Fragmento de molde compuesto (FCMP 607).
Discusión. Se trata de un género cosmopolita con un alcance estratigráfico muy amplio (Ordovícico a Triásico), que ha sido ampliamente referido para Norteamérica y Europa.
Familia Soleniscidae Knight, 1931
Género Strobeus Meek y Worthen, 1866
Especie tipo. Strobeus ventricosus de Koninck, 1881, por designación original. Carbonífero de Bélgica.
Strobeus cf. S. welleri (Knight, 1931)
Figuras 3.7, 3.8
Descripción. Concha subglobosa, con espira baja y vuelta corporal inflada. Tamaño mediano, de 12 mm de anchura y más de 14 mm de altura. Superficie ornamentada por costillas espirales finas, apenas visibles. Apertura alta y angosta, conservada de manera incompleta.
Material examinado. Un molde compuesto (FCMP 608).
Discusión. La espira baja en el ejemplar de Hidalgo permite relacionarlo con Strobeus welleri, sin embargo al no quedar preservados los detalles de otras estructuras diagnósticas como el pliegue columelar en el interior de la apertura, no es posible afirmar su identidad. S. welleri fue descrita inicialmente para el Pensilvánico de Missouri (Knight, 1931) y de Nuevo México (Kues y Batten, 2001). Posteriormente ha sido referida para el Pérmico inferior, Cisuraliano (Sakmariano–Artinskiano) de Texas (Nützel et al., 2000).
Clase Bivalvia Linnaeus, 1758
Superfamilia Nuculoidea Gray, 1824
Familia Nuculidae Gray, 1824
Género Nuculopsis Girty, 1911
Especie tipo. Nucula ventricosa Hall, 1858, por designación original. Carbonífero superior de Iowa, Estados Unidos.
?Nuculopsis sp.
Figuras 3.9, 3.10
Descripción. Concha pequeña, de 6.9 a 7.1 mm de longitud por 5.0 a 5.6 mm de altura, lo que da una relación L/A de 1.38 a 1.26. Equivalva, inequilateral, de contorno subtrigonal y moderadamente inflada. Umbo opistogiro que sobrepasa ligeramente el margen dorsal. Superficie externa con líneas concéntricas finas e irregulares, poco visibles en los ejemplares. Lúnula apenas perceptible; escudete pequeño, levemente marcado. No se observan los caracteres internos.
Material examinado. Un molde externo de una valva izquierda (FCMP 609) y un molde externo de una valva derecha (FCMP 610).
Discusión. El género Nuculopsis incluye varias especies del Carbonífero y Pérmico de Europa, América del Norte, América del Sur, Nueva Zelanda y Asia (Cox et al., 1969; Pagani, 2004a). Debido a que las características de las impresiones musculares, del ligamento y de la charnela no se han conservado en los ejemplares estudiados, no es posible confirmar su relación con estas especies.
Superfamilia Mytiloidea Rafinesque, 1815
Familia Mytilidae Rafinesque, 1815
Subfamilia Modiolinae Termier y Termier, 1950
Género Promytilus Newell, 1942
Especie tipo. Promytilus annosus, Newell, 1942, por designación original. Pensilvánico (Missuriano superior = Kasimoviano) de Kansas, Estados Unidos.
Promytilus sp.
Figura 3.11
Descripción. Concha oblicua, alargada y pequeña, de 7.2 mm de longitud por 5.1 mm de altura. Margen cardinal recto, posterior redondeado, formando ambos un ángulo de 132°; margen ventral oblicuo, ligeramente cóncavo posteriormente pero incrementando su convexidad cerca del margen anterior, el cual forma un tenue lóbulo. Umbo prosoclino, con un surco umbonal prominente. No se observan los detalles de la ornamentación.
Material examinado. Molde interno de una valva izquierda (FCMP 611).
Discusión. El género Promytilus ha sido descrito para rocas del Misisípico al Pérmico de América del Norte y América del Sur (Cox et al., 1969; Pagani, 2000). El ejemplar de Hidalgo es de dimensiones y forma muy parecidas a la especie Promytilus retusus descrita por Chronic (1952, p. 144, figs. 12–15) para la Formación Kaibab del Kunguriano–Roadiano de Walnut Canyon, Arizona, y al igual que el ejemplar de Arizona, parece ser equivalvo. Sin embargo, debido a que no se observan las características internas de la concha, no puede establecerse una asignación específica.
Superfamilia Ambonychioidea Miller, 1877
Familia Myalinidae Frech, 1891
Género Posidoniella de Koninck, 1885
Especie tipo. Inoceramus vetustus Sowerby, 1829, por designación original. Carbonífero inferior de Inglaterra.
Posidoniella sp.
Figura 3.12
Descripción. Concha de tamaño mediano, de 14 mm de longitud y 16.5 mm de altura, de forma ovalada. Umbo terminal que sobresale a una línea cardinal muy reducida, ya que no hay desarrollo de aurículas. Superficie cubierta por amplias arrugas concéntricas, sin ornamentación radial.
Material examinado. Molde externo de una valva izquierda (FCMP 612).
Discusión. El género Posidoniella es característico del Misisípico de Europa y ha sido referido para el Misisípico de Chile (Isaacson y Dutro, 1999) y para el Pensilvánico (Pottsville) de los Estados Unidos (Newell, 1969, p. 291). En México, Posidoniella sp. ha sido referida para el Pérmico medio de Olinalá, Guerrero (González–Arreola et al., 1994, p. 217). El ejemplar único no conserva estructuras diagnósticas que permitan establecer su identidad a nivel de especie.
Superfamilia Pterioidea Gray, 1847
Familia Pterineidae Meek, 1864
Género Leptodesma Hall, 1883
Especie tipo. Leptodesma potens Hall, 1883, por designación subsecuente de Miller, 1889. Devónico Superior de Nueva York, Estados Unidos.
Subgénero Leptodesma Hall, 1883
Leptodesma (Leptodesma) cf. L. (L.) falcata Boyd y Newell, 2001
Figura 3.13
Descripción. Concha de tamaño medio, de 21 mm de longitud y 16.1 mm de altura; de forma prosoclina y alargada transversalmente hacia el margen posterior. Aurícula anterior pequeña y angular; aurícula posterior incompleta, de forma extendida y aguzada en su extremo distal. Surcos umbonales anterior y posterior bien marcados. No se observan detalles de la ornamentación externa, de las impresiones musculares, del ligamento o de su dentición.
Material examinado. Molde interno de una valva izquierda (FCMP 613).
Discusión. El ejemplar de Hidalgo es considerado como Leptodesma (Leptodesma) por poseer una aurícula anterior angular, que lo distingue del otro subgénero, Leptodesma (Leiopoteria) Hall, 1883, en el cual es lobulada ( Newell y La Roque, 1969).
El ejemplar de Hidalgo es imperfecto, pero presenta el alargamiento con forma de hoz de la parte posterior del cuerpo de la valva, característico de Leptodesma (Leptodesma) falcata Boyd y Newell 2001, p. 4. Los ejemplares de esta especie conocidos hasta ahora provienen de una sola localidad (USNM 703c, in Cooper y Grant 1972) de la Formación Road Canyon del Pérmico medio (piso Roadiano) en Glass Mountains, oeste de Texas, EUA (Boyd y Newell, 2001). La especie, Pteria guadalupensis Girty 1908, p. 426, de la Formación Capitan de Guadalupe Mountains, tiene una apariencia externa similar a Leptodesma (L.)falcata, y sólo difiere de ésta en la forma de la aurícula anterior; sin embargo, el ejemplar estudiado por Girty carecía de esta aurícula y su forma original fue interpretada mediante el dibujo de línea con que ilustró su trabajo, pudiendo haberse alterado su forma original, por lo que posiblemente se trate de la misma especie. Si al encontrarse material mejor preservado se llega a comprobar la afinidad del ejemplar de Hidalgo con L. (L.) falcata, las implicaciones biogeográficas serían importantes, ya que esta especie no tiene una distribución amplia y forma parte de una serie de especies endémicas de vida corta (Boyd y Newell, 2001), por lo que su presencia en el Pérmico de Hidalgo permitiría explicar algunos registros disjuntos del Paleozoico superior relacionados con los taxa "Lazarus" (en el sentido de Jablonski, 1986).
Una especie afín, Leptodesma (Leptodesma) longa (Geinitz, 1866), del Pensilvánico de Norteamérica, presenta una concha más angosta y con un ángulo mayor entre la aurícula posterior y el surco umbonal.
Familia Bakevellidae King, 1850
Género Bakevellia King, 1848
Especie tipo. Avicula antiqua von Münster in Goldfuss, 1836, por designación original. Pérmico de Inglaterra.
Bakevellia sp.
Figuras 3.14–3.18
Descripción. Concha pequeña, de contorno romboidal y particularmente prosoclina, con un ángulo umbonal de casi 30°, que incrementa con el tamaño de los ejemplares. Valva izquierda ligeramente más grande, con un umbo que sobresale al margen dorsal; aurícula anterior lobulada; aurícula posterior alargada, con su extremo posterior sinuoso, ligeramente cóncavo. Externamente presenta costillas concéntricas gruesas, que en algunos ejemplares están dispuestas en series de pequeñas láminas finas, separadas por otras más gruesas. La valva derecha se observa parcialmente en un ejemplar en el que está sobrepuesto el molde de la valva izquierda, en ella se observa un umbo más bajo y su superficie cubierta únicamente por costillas finas y simples; en este mismo ejemplar se observa la impresión parcial del ligamento multivincular.
Material examinado. Dos moldes externos de valvas izquierdas (FCMP 614, FCMP 615); dos moldes internos de valvas izquierdas (FCMP 616, FCMP 617) y seis fragmentos (FCMP 618 – FCMP 623).
Dimensiones. (Tabla 3)
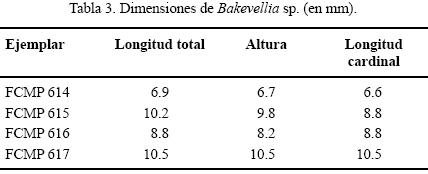
Discusión. La especie de Hidalgo se distingue rápidamente de otras especies de Bakevellia por la presencia de las arrugas concéntricas sobre su superficie. Sin embargo, al no conservarse estructuras diagnósticas internas en los ejemplares estudiados, no es posible proponer una especie nueva. El problema de homeomorfía entre las familias de bivalvos pterioideos ha sido discutido anteriormente por Boyd y Newell (2001) y de no haberse preservado la impresión del ligamento en uno de los ejemplares de Hidalgo (Figura 3.18), no hubiera sido posible su identificación genérica, pues es evidente la semejanza superficial del género Bakevellia con algunos representantes del género Leptodesma (Familia Pterineidae). Bakevellia fue establecido para el Pérmico de Inglaterra (Boyd y Newell, 2001) y varias especies del Pérmico de Norteamérica han sido relacionadas con él, a pesar de que en la mayoría de los casos no han quedado preservadas las características del ligamento, el cual es duplivincular en Leptodesma y multivincular en Bakevellia, por lo que es muy probable que algunas de las especies de Bakevellia referidas para el Pérmico de Norteamérica correspondan en realidad al género Leptodesma. Otro caso de convergencia se observa entre la ornamentación de la especie de Hidalgo con la del género Elversella McRoberts y Newell, 2001 (Familia Myalinidae), ya que su especie tipo, Elversella rugosa McRoberts y Newell, 2001, presenta una ornamentación similar; sin embargo, al igual que en el caso anterior, esta familia presenta un ligamento duplivincular.
Se trata de un género cosmopolita con un amplio alcance estratigráfico que va del Carbonífero al Cretácico. En México, el género Bakevellia ha sido referido para el Pérmico (Artinskiano–Kunguriano) de la Caliza Colina, en el norte de Sonora (Blodgett et al., 2002).
Superfamilia Incierta
Familia Euchondriidae Newell, 1938
Género Euchondria Meek, 1874
Especie tipo. Pecten neglectus Geinitz, 1866, por designación original. Pensilvánico de Nebraska, Estados Unidos.
Euchondria cf. E. cooperi Newell y Boyd, 1995
Figuras 3.19, 4.1
Descripción. Fragmento de una valva izquierda pequeña, de 9 mm de longitud, inequilateral y prosoclina. Cuerpo de la valva con 10 costillas principales ampliamente espaciadas y ligeramente arqueadas, siguiendo la forma del disco de la valva; cada costilla está intercalada con una costilla secundaria que se extiende antes de la mitad de la valva hacia el margen ventral del disco. Espacio intercostelar cubierto por líneas de crecimiento muy próximas entre sí, de 17 a 20 por mm y ligeramente arqueadas hacia el umbo. Las aurículas no se conservaron.
Material examinado. Molde externo de una valva izquierda (FCMP 632).
Discusión. Si bien algunas de sus especies han sido referidas para Australia y para Argentina, el género Euchondria, al igual que la Familia Euchondriidae, estuvo ampliamente distribuido en Laurasia, desde el Misisípico hasta el Pérmico, (Newell y Boyd, 1995). La especie Euchondria cooperi Newell y Boyd, 1995 se ha referido para el Kunguriano (Formación Road Canyon)–Guadalupiano Inferior (Formación Willis Ranch), del oeste de Texas (Newell y Boyd, 1995). El ejemplar único de Hidalgo comparte la ornamentación típica de esta especie, pero su estado fragmentario impide una determinación definitiva.
Superfamilia Aviculopectinoidea Meek y Hayden, 1865
Familia Aviculopectinidae Meek y Hayden, 1865
Género Aviculopecten M'Coy, 1851
Especie tipo. Aviculopecten planoradiatus M'Coy, 1851, por designación subsecuente de Hind, 1903. Carbonífero de Derbyshire, Inglaterra.
Aviculopecten cf. A. basilicus Newell, 1938
Figura 4.2
Descripción. Un fragmento de una valva izquierda. Su superficie está cubierta por costillas finas con interespacios dos veces el ancho de las mismas; en la parte central del disco las costillas están divididas a partir del extremo cercano al umbo.
Material examinado. Un fragmento (FCMP 636).
Discusión. El género Aviculopecten ha sido descrito para el Carbonífero superior–Pérmico medio de Laurasia (Newell y Boyd, 1995). El estado fragmentario del material no permite una comparación más precisa con las especies de este género, sin embargo, en el ejemplar de Hidalgo se observa la división de las costillas hacia el margen ventral descrita para los estadios ontogenéticos seniles de la especie A. basilicus Newell, 1938. Esta especie, junto con A. arctisulcatus Newell, A. exemplarius Newell y A. occidentalis (Shumard), son cuatro especies del Carbonífero Superior descritas para el sur del Mesocontinente (Midcontinent) de los Estados Unidos que son muy similares entre sí y que han sido consideradas como posibles variedades de una sola especie (Ciriacks, 1963). Una especie del Pérmico de Wyoming ha sido referida a A. basilicus y se encuentra entre los límites de variabilidad de este grupo (Ciriacks, 1963). Esta misma especie ha sido referida para el Morrowano–Desmoinesiano de Oaxaca, en México (Quiroz–Barroso y Perrilliat, 1998). Lo imperfecto e incompleto del ejemplar único de Hidalgo no permite asegurar su identidad.
Familia Deltopectinidae Dickins, 1957
Subfamilia Cyrtorostrinae Newell y Boyd, 1995
Género Cyrtorostra Branson, 1930
Especie tipo. Cyrtorostra varicostata Branson, 1930, por designación original. Pérmico inferior de Wyoming, Estados Unidos.
Cyrtorostra cf. C. varicostata Branson, 1930
Figura 4.3
Descripción. Valva pequeña, de 10.2 mm de longitud por 10.1 mm de altura. Su forma es convexa, casi circular. La superficie del disco presenta once costillas angulares y gruesas; en dos de ellas es posible observar la forma de espina en que terminan. Los interespacios son anchos con una sola costilla. Las aurículas no se conservaron.
Material examinado. Un molde externo de una valva izquierda (FCMP 637).
Discusión. El género Cyrtorostra se ha referido para el Pérmico inferior y medio, y es cosmopolita en el Hemisferio Norte (Newell y Boyd, 1995). El ejemplar de Hidalgo posiblemente esté relacionado con la especie C. varicostata Branson, 1930, descrita para los miembros Ervay, Tosi, Shedhorn y Rex Chert del Complejo Park City–Phosphoria en Wyoming (Ciriacks, 1963), y para el Asseliano–Sakmariano (Formación Neal Ranch), Sakmariano?–Kunguriano, (Fm. Cathedral Mt.) Wordiano (Fm. Road Canyon) y Capitaniano (Fm. Bell Canyon) de Glass Mountain (Newell y Boyd, 1995), pues el tamaño y la forma del disco, así como la forma y el número de las costillas, están dentro del rango de variabilidad de la especie, pero al no quedar preservadas las aurículas, el umbo, ni los detalles terminales de las costillas, no puede asegurarse su relación.
Superfamilia Chaenocardioidea Miller, 1889
Familia Chaenocardiidae Miller, 1889
Género Obliquipecten Hind, 1901
Especie tipo. Obliquipecten laevis Hind, 1901, por monotipia. Carbonífero inferior de Inglaterra.
Obliquipecten sp.
Figura 4.4
Descripción. Un molde interno imperfecto de una valva derecha de tamaño medio (14.3 mm de ancho por 21.3 mm de altura). Su forma es ovalada, con el margen dorsal posterior obtuso. El umbo es bajo, pequeño, prosogiro. La superficie interna es lisa. Se conserva parte de una aurícula anterior grande y subcircular con seno bisal profundo, mientras que la aurícula posterior es muy reducida. El ejemplar está comprimido lateralmente por procesos diagenéticos.
Material examinado. Molde interno de una valva derecha (FCMP 638).
Discusión. El género se ha referido para el Misisípico de Europa; para el Pérmico (Asseliano–Guadalupiano) del oeste de Texas (Newell y Boyd, 1995) y para el Pérmico de Wyoming (Ciriacks, 1963). Al no haber sido encontrados ejemplares pensilvánicos, Newell y Boyd (1995) consideran que el género ocupó un refugio en alguna otra parte durante el intervalo comprendido entre el Misisípico y el Pérmico. En su forma general, la especie de Hidalgo presenta afinidad con Obliquipecten granti Newell y Boyd, 1995 del Pérmico (Sakmariano–Roadiano) del oeste de Texas, pero lo imperfecto de la conservación impide una comparación más detallada.
Familia Streblochondriidae Newell, 1938
Subfamilia Streblochondriinae Newell, 1938
Género Streblochondria Newell, 1938
Especie tipo. Aviculopecten sculptilis Miller, 1891, por designación original. Pensilvánico de Missouri, Estados Unidos.
Streblochondria sculptilis (Miller, 1891)
Figuras 4.5, 4.6
Aviculopecten sculptilis Miller, 1891, p. 92, lám. 20, fig. 5; Beede, 1900, p. 122, lám. 13, figs. 3–3b; Sayre, 1930, p. 119, lám. 11, figs. 9, 10.
Streblochondria sculptilis Newell, 1938, p. 82, lám. 16, figs. 5a–c, 7, 9a, b, 11; Newell y Boyd, 1985, p. 2, figs. 1.1, 1.2a, 1.2b; Newell y Boyd, 1995, p. 56, fig. 41.
Descripción. Concha ligeramente más alta (10.3 mm) que larga (10.1 mm); el margen dorsal es la mitad de la longitud del disco; aurícula anterior aproximadamente la mitad a tres quintas partes la longitud de la línea cardinal. Superficie del disco con 60 a 70 costillas radiales en dos rangos de tamaños alternantes, cruzadas por laminitas concéntricas dispuestas regularmente formando un patrón cancelado con escamas delicadas en las intersecciones.
Material examinado. Molde externo de una valva derecha (FCMP 634); molde externo de una valva izquierda (FCMP 635).
Discusión. El género se presenta en el Misisípico–Pérmico Superior de Laurasia (Newell y Boyd, 1995). La especie Streblochondria sculptilis (Miller, 1891) tiene una distribución estratigráfica muy amplia que va del Pensilvánico Superior al Pérmico medio (Capitaniano). Es especialmente abundante en la Formación Bone Springs (Sakmariano?–Kunguriano), del oeste de Texas (Newell y Boyd, 1995).
Subfamilia Streblopteriinae Waterhouse, 2008
Género Streblopteria M'Coy, 1851
Especie tipo. Meleagrina laevigata M'Coy, 1844, por designación subsecuente de Meek y Worthen, 1866, p. 333. Carbonífero inferior de Irlanda.
Streblopteria montpelierensis (Girty, 1908)
Figura 4.7
Aviculopecten? montpelierensis Girty, 1910, p. 42, pl. 4, figs. 9, 10
Streblochondria? montpelierensis Girty. Newell, 1938 , p. 83, pl. 14, figs. 6, 7; Ciriacks, 1963, p. 55, pl. 8, figs. 9–15
Streblopteria montpelierensis Girty. Newell y Boyd, 1995, p. 50, fig. 37
Descripción. El único espécimen es una valva derecha pequeña, de 9.4 mm de longitud, 9.5 mm de altura y 5.5 mm de longitud en la línea cardinal; su forma es marcadamente convexa, de contorno subcircular. El umbo sobresale ligeramente del margen dorsal; aurícula anterior pequeña, recta, alargada y lobulada, con un seno bisal profundo; aurícula posterior muy reducida, angosta y triangular. Superficie externa lisa.
Material examinado. Molde externo de una valva derecha (FCMP 633).
Discusión. Streblopteria montpelierensis (Girty, 1908) es una especie muy abundante en el oeste americano, encontrándose en el Asseliano–Capitaniano del oeste de Texas y en el Miembro Meade Peak de la Formación Phosphoria en Middle Rockies, en los estados de Idaho, Wyoming y Montana (Newell, 1938; Ciriacks, 1963). En México, una especie no figurada de la Formación Monos del Pérmico de El Antimonio, Sonora, fue referida por Cooper et al. (1965) como Aviculopecten aff. A. montpelierensis Girty.
Superfamilia Heteropectinoidea Beurlen, 1954
Familia Heteropectinidae Beurlen, 1954
Subfamilia Cassianoidinae Newell y Boyd, 1995
Género Cassianoides Newell y Boyd, 1995
Especie tipo. Cassianoides kingorum Newell y Boyd, 1995, por designación original. Pérmico inferior a medio del oeste de Texas, Estados Unidos.
Cassianoides sp.
Figura 4.8
Descripción. Concha pequeña, de 5.5 mm de longitud y 5.4 mm de altura, retrocrescente, más alta que larga y fuertemente convexa. Sobre su superficie se observan claramente seis costillas primarias ampliamente espaciadas y laminillas intercostelares concéntricas, cóncavas hacia el margen ventral. Las aurículas no se conservaron.
Material examinado. Molde externo de una valva izquierda (FCMP 627).
Discusión. El pequeño tamaño del ejemplar de Hidalgo y la presencia de seis costillas radiales separadas por interespacios anchos y profundos, atravesadas por laminitas concéntricas muy finas, poco espaciadas y arqueadas dorsalmente, permite relacionarlo con Cassianella sexradiata (Branson, 1930) del Miembro Ervay en Bull Lake, Wyoming (Ciriacks, 1963). Sin embargo, en el ejemplar de Hidalgo no es posible observar la depresión radial que separa a la aurícula anterior de la región umbonal, característica de este género. Otras especies de tamaño y formas similares a la especie de Hidalgo son Cassianoides kingorum Newell y Boyd, 1995, del Pérmico inferior–medio del oeste de Texas, y Aviculopecten crassispinosus Chronic, 1953, p. 155, lám. 33, figs. 13a–b, del Pérmico inferior del Grupo Copacabana, Perú. Ambas poseen costillas con espinas gruesas de forma tubular, características del género Cassianoides. Las tres especies anteriores son consideradas actualmente como pertenecientes al género Cassianoides. En el ejemplar de Hidalgo no se observan espinas, pero debido a lo imperfecto del molde no es posible asegurar que no hubieran estado en la concha original; por otra parte, la variabilidad intraespecífica de este caracter es muy grande, y uno de los ejemplares descritos por Chronic (1953, lám. 33, fig. 13a, b) posee espinas reducidas y un aspecto muy parecido al ejemplar de Hidalgo. De acuerdo a lo anterior no es posible asegurar la especie con el ejemplar único.
Familia Acanthopectinidae Newell y Boyd, 1995
Género Acanthopecten Girty, 1903
Especie tipo. Pecten carboniferous Stevens, 1858, por designación original. Pensilvánico Superior de Nebraska, Estados Unidos.
Acanthopecten coloradoensis (Newberry, 1861)
Figuras 4.9–4.12
Pecten (Monotis?) coloradoensis Newberry, 1861, p. 129, pl. 1, figs. 6, 6a. Acanthopecten coloradoensis Newberry. Newell, 1938, p. 75, pl. 12, figs. 7a, b, 13–15b; Chronic, 1952, p. 144, pl. 8, fig. 8; Ciriacks, 1963, p. 52, pl. 7, figs. 8–11; Newell y Boyd, 1995, p. 45, figs. 32, 33.
Descripción. Valvas izquierdas más largas que altas en las formas juveniles y circulares o subcirculares en los especimenes adultos. Margen dorsal recto, con la aurícula posterior esbelta, triangular y más larga que la anterior, la cual está definida por un surco angosto. Ángulo umbonal en la concha adulta es de 109°, contrastado con los 115° referidos anteriormente para la especie. Ornamentación externa con 22 costillas gruesas que no incrementan en número hacia el margen ventral, cruzadas por laminitas concéntricas que forman espinas pequeñas con aspecto de V invertida sobre su superficie.
Material examinado. Moldes externos de las valvas izquierdas de dos ejemplares juveniles (FCMP 628, FCMP629) y de dos adultos (FCMP 630, FCMP 631).
Dimensiones. (Tabla 4)
Discusión. La presencia de numerosas costillas radiales estrechamente espaciadas, el gran número de laminitas concéntricas y la forma de las espinas que cubren su superficie, permiten relacionar a los ejemplares de Hidalgo con la especie descrita para Estados Unidos. Los ejemplares completos son formas juveniles y su aspecto es similar a formas también juveniles figuradas previamente (Ciriacks, 1963, pl. 7, fig. 9). El fragmento mejor conservado presenta costillas y espinas iguales al ejemplar referido en Newell y Boyd (1995, figs. 32–4b). Acanthopecten coloradoensis se ha encontrado en el Pérmico (Cisuraliano Superior y Guadalupiano) de la Formación Word del oeste de Texas (Newell y Boyd, 1995); de la Formación Kaibab de Arizona (Newell, 1938; Chronic, 1952) y de Nevada (Newell, 1938). También ha sido referida para la Formación Phosphoria (Guadalupiano?) de Montana, Wyoming y Utah (Ciriacks, 1963).
Superfamilia Limoidea Rafinesque, 1815
Familia Limidae Rafinesque, 1815
Género Palaeolima Hind, 1903
Especie tipo. Pecten? simplex Phillips, 1836, p. 212, por designación subsecuente de Cox, 1952, p. 48. Carbonífero inferior de Inglaterra.
?Palaeolima sp.
Figuras 4.13–4.15
Descripción. Concha pequeña, opistoclina, moderadamente convexa, de forma ovalada y oblicua, ligeramente más alta que ancha ( 5.5 mm / 4.7 mm en el ejemplar mejor conservado). Umbo ortogiro de posición media, que sobresale levemente a la línea cardinal. Margen cardinal recto y corto, que forma ángulos obtusos con las aurículas; margen anterior ligeramente curvo; margen posterior redondeado, haciéndose más convexo hacia el margen ventral. Superficie de la valva con 18 a 20 costillas radiales delgadas y angulares, algunas de ellas desarrolladas por intercalación; interespacios anchos; ornamentación concéntrica consiste en numerosas líneas, muy próximas entre sí y espaciadas regularmente, que atraviesan tanto los interespacios como la superficie de las costillas. No se distingue el surco umbonal; superficie de las aurículas con una ornamentación de costillas muy finas, atravesadas por líneas concéntricas.
Material examinado. Molde externo de dos valvas derechas (FCMP 624; FCMP 625); fragmento de la impresión de una valva derecha (FCMP 626).
Discusión. La mala preservación del material de Hidalgo impide una comparación detallada con otras especies. Sin embargo, presenta la ornamentación de costillas finas y el delicado patrón de líneas concéntricas que caracteriza a la especie Limatulina striaticostata Girty (1908) de la parte media de la Formación Capitan, Guadalupe Mountains, Texas.
Superfamilia Trigonioidea Lamarck, 1819
Familia Schizodidae Newell y Boyd, 1975
Subfamilia Schizodinae Newell y Boyd, 1975
Género Schizodus de Verneuil y Murchison, 1844
Especie tipo. Axinus obscurus Sowerby, 1821, p. 12, por designación subsecuente de Verneuil, 1845. Pérmico superior de Inglaterra.
Schizodus texanus Clifton, 1942
Figuras 4.16 y 5.1
Schizodus texanus Clifton, 1942, p. 691–693, lám. 101, figs. 20–28, lám. 102, figs. 10–13; Newell y Boyd, 1975; p.116, figs. 13A y C, 52, 53.
Descripción. Dos conchas de tamaño medio, de 47–48 mm de longitud por 38–39 mm de altura, ortogira, con el umbo elevado. El margen anterior describe una amplia curva que se extiende desde el umbo hasta el margen ventral, el cual es suavemente convexo; el margen posterior está atenuado, redondeado en los extremos dorsal y ventral. Se presenta una carina posterior ampliamente redondeada, que se extiende desde el umbo hacia el margen postero–ventral. Canal del ligamento alargado y conservado parcialmente. La superficie está cubierta con líneas de crecimiento finas.
Material examinado. Moldes compuestos de una valva izquierda (FCMP 639) y una valva derecha (FCMP 640).
Discusión. Schizodus texanus Clifton, 1942 es una especie ampliamente distribuida en rocas del Asseliano al Roadiano de Texas, Nuevo México y Arizona, con sus mejores representaciones en las formaciones San Andres, Blaine y Kaibab (Newell y Boyd, 1975). Presenta una ornamentación similar a la especie pensilvánica S. ulrichi Worthen, 1890, pero esta última presenta un margen posterior truncado oblicuamente y un umbo anterior.
Es la especie de mayor tamaño del material estudiado de Hidalgo.
Subfamilia Eoastartinae Newell y Boyd, 1975
Género Eoastarte Ciriacks, 1963
Especie tipo. Eoastarte subcircularis Ciriacks, 1963, por designación original. Pérmico inferior (Sakmariano?–Kunguriano) de Arizona, Estados Unidos.
Eoastarte subcircularis Ciriacks, 1963
Figuras 5.2–5.4
Eoastarte subcircularis Ciriacks, 1963, p. 66,–67, lám. 12., figs 11–15; Newell y Boyd, 1975, p. 133, figs. 68–70.
Descripción. Tres moldes internos imperfectos de tamaño medio, el ejemplar más completo mide 36 mm de altura. El umbo es prominente y prosogiro. La forma de la concha es convexa, con el margen posteroventral ligeramente alargado. Se observa claramente un surco radial posterior.
Material examinado. Molde interno de una valva izquierda (FCMP 641); dos fragmentos (FCMP 642 y FCMP 643).
Discusión. El género Eoastarte es característico del Sakmariano?–Kunguriano de Wyoming, Texas y Arizona. La especie E. subcircularis ha sido referida para el Pérmico (Sakmariano?–Kunguriano) de la Formación Park City, Miembro Grandeur, en Wyoming (Ciriacks, 1963); de las formaciones Cathedral Mountain y Road Canyon, en el oeste de Texas, y de la Formación Kaibab, Miembro Gamma, en Arizona (Newell y Boyd, 1975). Los ejemplares provenientes de estas formaciones presentan una combinación inusual de caracteres y una notable variación ontogenética y han sido considerados como una sola especie por Newell y Boyd (1975).
Los ejemplares de Hidalgo son más grandes que la mayoría de los especimenes referidos para esta especie, sin embargo, un ejemplar de Texas presenta dimensiones mayores (41 mm de longitud y 42 mm de altura). El surco posterior ha sido interpretado como posible área de fijación de dos pares de placas branquiales y es una característica diagnóstica de esta especie (Newell y Boyd, 1975).
Familia Trigoniidae Lamarck, 1819
Género Lyroschizodus Newell y Boyd, 1975
Especie tipo. Lyroschizodus orbicularis Newell y Boyd, 1975, por designación original. Pérmico de Texas, Estados Unidos.
Lyroschizodus cf. L. orbicularis Newell y Boyd, 1975
Figuras 5.5, 5.6
Descripción. Concha subtrigonal, de altura y longitud similares. El umbo es prosogiro, situado ligeramente anterior a la línea media. El margen anterior es convexo; el margen posterior es trunco, con el margen respiratorio más grande que la mitad de la altura de la concha. Presenta una costilla posterior bien definida, que es más angular en la forma juvenil. La superficie de la concha presenta costillas concéntricas angostas y estrechamente espaciadas.
Material examinado. Un molde externo de una valva derecha de juvenil (FCMP 644); un molde externo imperfecto de una valva derecha de adulto (FCMP 645); un fragmento (FCMP 646).
Dimensiones. La concha del juvenil tiene 15.2 mm de ancho por 12.3 mm de altura; la del adulto tiene 44.2 mm de ancho por 38.1 mm de altura.
Discusión. Los ejemplares de Hidalgo son muy similares a Lyroschizodus orbicularis en dimensiones y ornamentación, con excepción de su margen ventral que es recto. Sin embargo, al no poder observarse su morfología interna no es posible afirmar esta asignación. L. orbicularis es abundante en el Sakmariano?–Kunguriano y Guadalupiano inferior y se ha referido para las formaciones Cathedral Mountain y Road Canyon del oeste de Texas (Newell y Boyd, 1975). Se distingue de otra especie pérmica de Oklahoma y Texas, Lyroschizodus oklahomensis Beede, 1907, por su costilla posterior claramente definida.
Superfamilia Crassatelloidea Férussac, 1822
Familia Astartidae d'Orbigny, 1844
Género Astartella Hall, 1858
Especie tipo. Astartella vera Hall, 1858, por designación subsecuente de Miller, 1889. Pensilvánico de Estados Unidos.
Astartella subquadrata Girty, 1909
Figura 5.7
Astartella subquadrata Girty, 1909, p. 94, pl. 10, figs. 1013; Clifton, 1942, p. 693; Chronic, 1952, p. 150, pl. 10, figs. 5–15; Ciriacks, 1963, p. 65, pl. 12, figs. 6–9.
Descripción. Concha pequeña (el ejemplar mejor conservado presenta 12 mm de longitud y 11 mm de altura), por lo que es casi tan larga como alta. Los márgenes cardinal y ventral son ligeramente curvos y casi paralelos. El umbo es prosogiro y moderadamente alto. La lúnula es alargada y cóncava; el escudete está alargado y bien marcado. La superficie está cubierta con 18 costillas concéntricas delgadas y regularmente espaciadas, separadas por interespacios anchos y cóncavos. Los rasgos internos de la concha no se conservaron.
Material examinado. Molde externo y contramolde de valva derecha (FCMP 647); un molde interno imperfecto de una valva izquierda (FCMP 648).
Discusión. Astartella subquadrata Girty, 1909 se ha descrito para el Pérmico inferior y medio del suroeste de Estados Unidos: Formación Kaibab, en Arizona; formaciones San Andres y Yeso en Nuevo Mexico; caliza Word Number 1, y probablemente otros horizontes del oeste de Texas. También ha sido referida para Utah, y Idaho (Ciriacks, 1963). En México, ha sido referida (no figurada) para el Pérmico (Sakmariano?–Kunguriano) de la Caliza Colina, en el norte de Sonora (Blodgett, et al., 2002). La especie Astartella nasuta, descrita por Girty (1908) para la formación Delaware Mountain (Sakmariano?–Kunguriano) del oeste de Texas, y referida también para el Grupo Copacabana (Pérmico inferior) de Perú (Chronic, 1953), es muy similar a A. subquadrata, si bien es ligeramente más larga que esta última. Por otra parte, varios ejemplares figurados para A. subquadrata presentan la proyección en la parte inferior del extremo anterior, característica de A. nasuta y a la cual deben su nombre. Ciriacks (1963), menciona que especies similares han sido referidas para Groenlandia, Inglaterra, Alemania y Rusia y que muy probablemente las dos especies de Norteamérica antes referidas sean en realidad miembros de una sola especie, ampliamente distribuida durante el Pérmico.
Superfamilia Pholadomyoidea King, 1844
Familia Sanguinolitidae S. A. Miller, 1877
Subfamilia Sanguinolitinae S. A. Miller, 1877
Género Sanguinolites M'Coy, 1844
Especie tipo. Sanguinolites discors, por designación subsecuente de Stoliczka, 1871. Carbonífero inferior de Irlanda.
Sanguinolites sp.
Figura 5.8
Descripción. Se trata de un fragmento del margen postero–ventral de una valva izquierda, en el que se observa parte de la forma y de la ornamentación de la concha características del género: un margen posterior truncado; carena umbonal que se extiende hacia el margen postero–ventral; laminillas de crecimiento concéntricas, paralelas entre sí y separadas por interespacios angostos que terminan en la carena umbonal a partir de la cual se transforman en líneas que recorren el extremo posterior.
Material examinado. Impresión externa del extremo posterior de una valva izquierda (FCMP 649).
Discusión. En México se ha referido con duda la presencia del género Sanguinolites para el Pérmico (Sakmariano–Artinskiano?) de la Caliza Colina, norte de Cananea, Sonora (Blodgett et al., 2002). El fragmento de Hidalgo presenta similitud con la parte posterior de Sanguinolites costata (Meek y Worthen, 1869) del Carbonífero de Norteamérica, especialmente con los especímenes figurados para los grupos Allegheny y Conemaugh del Pensilvánico de Ohio (Hoare et al., 1979). También es similar a Sanguinolites tentoriolus (Chronic, 1953), descrita para el Grupo Copacabana de Perú, del Pérmico inferior. Al desconocerse la morfología de la mayor parte de la concha, no puede conocerse su identidad a nivel de especie.
Superfamilia Orthonotoidea S. A. Miller, 1877
Familia Solenomorphidae Cockerell, 1915
Subfamilia Solenomorphinae Cockerell, 1915
Género Solenomorpha Cockerell, 1903
Especie tipo. Solenopsis minor M'Coy, 1844, por monotipia. Carbonífero inferior de Irlanda.
?Solenomorpha sp.
Figura 5.9
Descripción. Una sola valva derecha de tamaño medio, de 36 mm de longitud y 9.2 mm de altura. Su aspecto es subcuneiforme, cilíndrica, fuertemente inequilateral, con el umbo situado en el extremo anterior. El margen anterior no es visible en el ejemplar estudiado; el margen dorsal es recto; el margen ventral es ligeramente convexo; el margen posterior está suavemente redondeado. La superficie externa presenta suaves líneas de crecimiento, sin ornamentación radial.
Material examinado. Molde externo de una valva derecha (FCMP 650).
Discusión. El estado de conservación del ejemplar único de Hidalgo no permite observar las características del escudete y de la lúnula. Tampoco es visible la presencia de una carena umbonal, por lo que se relaciona con duda al género Solenomorpha, el cual fue cosmopolita y cuyo registro se ha encontrado en rocas del Devónico Inferior al Pérmico superior. El género Alula Girty, 1912 del Pérmico inferior de América del Norte es similar externamente, pero se diferencía por presentar costillas radiales finas.
CONCLUSIONES
Se reconocen cinco especies correspondientes a cinco familias de gasterópodos y 20 especies correspondientes a 18 familias de bivalvos, las cuales representan en su mayoría nuevos registros para México.
El estado fragmentario y la selección de tamaños de los individuos indica un retrabajo que concuerda con el modelo propuesto por Rosales–Lagarde et al., (2005) el cual plantea que el material quedó sepultado por flujos turbidíticos ligados a un arco magmático submarino, que removieron depósitos marinos someros.
La actividad volcánica debió ser un factor externo importante que influyó en la diversidad y abundancia de la biota marina al mantener una temperatura elevada y un aumento en la cantidad de nutrientes disponibles.
Los gasterópodos y bivalvos de Otlamalacatla son característicos del Pérmico temprano (Cisuraliano), especialmente del Kunguriano y presentan una fuerte afinidad con las faunas tropicales y subtropicales de la provincia Grandian (Yancey, 1975) que ocupó el área de Texas–Nuevo México–Coahuila durante el Pérmico temprano y medio.
AGRADECIMIENTOS
A Daniel Navarro Santillán por su colaboración durante el trabajo de campo y a Héctor Hernández Campos por la elaboración del material fotográfico. A la Universidad Nacional Autónoma de México por su apoyo a esta investigación a través de los proyectos PAPIIT IN 209308 e IN 213710–3. A M. Alejandra Pagani y a Roberto Ricardo Lech por las revisiones hechas a este trabajo, las cuales enriquecieron su contenido.
REFERENCIAS
Aberhan, M., Alroy, J., Fursich, F.T., Kiessling, W., Kosnik, M., Madin, J., Patzkowsky, M., Wagner, P., 2004, Ecological Attributes of Marine Invertebrates. Unpublished database, available online: http://paleodb.org/. [ Links ]
Amler, M.R.W., 1999, Synoptical classification of fossil and recent Bivalvia: Geologica et Paleontologica, 33, 237–248. [ Links ]
Anelli, L.E., Rocha–Campos, A.C., Simoes, M.G., 2002, Protobranch bivalves from the Piauí Formation (Middle Pennsylvanian), Paranaíba Basin, Brazil: Revista Española de Paleontología, 17, 165–176. [ Links ]
Anelli, L.E., Rocha–Campos, A.C., Simoes, M.G., 2006, Pennsylvanian pteriomorphian bivalves from the Piauí Formation, Parnaíba Basin, Brazil: Journal of Paleontology, 80(6), 1125–1141. [ Links ]
Arellano–Gil, J., Vachard, D., Yussim, S., Flores de Dios–González, L., 1998, Aspectos estratigráficos, estructurales y paleogeográficos del Pérmico Inferior al Jurásico Inferior en Pemuxco, estado de Hidalgo, México: Revista Mexicana de Ciencias Geológicas, 15(1), 9–13. [ Links ]
Bambach, R.K., 1983, Ecospace utilization and guilds in marine communities through the Phanerozoic, en Tevesz, M. and McCall, P. (eds.), Biotic Interactions in Recent and Fossil Benthic Communities: Plenum, 719–746. [ Links ]
Bandel, K., 2007, Description and classification of Late Triassic Neritimorpha (Gastropoda, Mollusca) from the St. Cassian Formation, Italian Alps: Bulletin of Geosciences, 82(3), 215–274. [ Links ]
Beede, J.W., 1900, Carboniferous invertebrates: Kansas Geological Survey, Report, 6(2), 187 p. [ Links ]
Beede, J.W. , 1907, Invertebrate paleontology of the Upper Permian red beds of Oklahoma and the Panhandle of Texas: Kansas University Science Bulletin, 4, 113–171. [ Links ]
Beurlen, K., 1954, As faunas de lamellibranquios do sistema genduanico no Parana, en Lange, F.W. (ed.), Paleontologia do Parana: Comissao de Comemoracoes do Centenario do Parana, Curitiba, 107–136. [ Links ]
Blodgett, R.B. , Moore, T.E., Gray, F., 2002, Stratigraphy and paleontology of Lower Permian rocks north of Cananea, northern Sonora, Mexico: Journal of South American Earth Sciences, 15, 481–495. [ Links ]
Bouchet, P., Rocroi, J.P., Fryda, J., Hausdorf, B., Ponder, W., Valdés, A., Warén, A., 2005, Classification and nomenclator of gastropod familias: Malacologia, 47(1–2), 397 p. [ Links ]
Bouchet, P., Rocroi J.P., Bieler, R., Carter, J.G, Coan E.V., 2010. Nomenclator of bivalve families with a classification of bivalve families: Malacologia, 52(2), 184 p. [ Links ]
Boyd, D.W., Newell, N.D., 2001, Silicified Leptodesma (Bivalvia; Pteriomorphia) from the Texas Permian: American Museum Novitates, 3347, 10 p. [ Links ]
Branson, C.C., 1930, Paleontology and stratigraphy of the Phosphoria Formation: University of Missouri Studies, 5(2), 98 pp. [ Links ]
Bridges, L.W., 1964, Geology of mina Plomosas–Placer de Guadalupe area, Chihuahua, Mexico: West Texas Geological Society, 50–64. [ Links ]
Campa, M.F., Coney, P.J., 1983, Tectono–stratigraphic terranes and mineral resource distributions in Mexico: Canadian Journal of Earth Sciences, 20, 1040–1051. [ Links ]
Carrillo–Bravo, J., 1961, Geología del Anticlinorio Huizachal– Peregrina al NW de Ciudad Victoria, Tamaulipas: Boletín de la Asociación Mexicana de Geólogos Petroleros, 13(1–2), 1–98. [ Links ]
Carrillo–Bravo, J., 1965, Estudio geológico de una parte del Anticlinorio de Huayacocotla: Boletín de la Asociación Mexicana de Geólogos Petroleros, 17(5–6), 73–96. [ Links ]
Carter, J.G., Campbell, D.C., Campbell, M.R., 2000, Cladistic perspectives on early bivalve evolution, en Harper, E.M., Taylor, J.D., Crame, J.A. (eds.), The Evolutionary Biology of the Bivalvia: Geological Society Special Publication 177, 47–79. [ Links ]
Centeno–García, E., 2005, Review of Upper Paleozoic and Lower Mesozoic stratigraphy and depositional environments of central and West Mexico: Constraints on terrane analysis and paleogeography, en Anderson, T.H., Nourse, J.A., McKee, J.W., Steiner, M.B. (eds.), The Mojave–Sonora megashear hypothesis: Development, assessment and alternatives: Geological Society of America Special Paper 393, 233–258. [ Links ]
Chronic, H., 1952, Molluscan fauna from the Permian Kaibab Formation, Walnut Canyon, Arizona: Geological Society of America Bulletin, 63, 95–166. [ Links ]
Chronic, H., 1953, Invertebrate palentology (excepting fusulinids and corals), en Newell, N.D., Chronic, J., Roberts, T.G. (eds.), Upper Paleozoic of Peru: Geological Society of America Memoir 58, 43–165. [ Links ]
Ciriacks, K.W., 1963, Permian and Eotriassic bivalves of the Middle Rockies: American Museum of Natural History Bulletin, 125, 100 pp. [ Links ]
Clifton, R.L., 1942, Invertebrate faunas from the Blaine and the Dog Creek formations of the Permian Leonard Series: Journal of Paleontology, 16(6), 685–699. [ Links ]
Cockerell, T.D.A., 1903, Some homonymous generic names: Nautilus, 16, 118. [ Links ]
Cockerell, T.D.A., 1915, The Solenopsidae: The Nautilus, 29(7), 84. [ Links ]
Cooper, G.A., Grant, R.E., 1972, Permian brachiopods of West Texas, I: Smithsonian Contributions to Paleobiology, 14, 231 p. [ Links ]
Cooper, G.A., Dunbar, C.O., Duncan, H., Miller, A.K., Night, J.B., 1965, Fauna Pérmica de El Antimonio, oeste de Sonora: Boletín del Instituto de Geología, Universidad Nacional Autónoma de México, 58, 1–122. [ Links ]
Cox, L.R., 1952, Notes on Trigoniidae, with outlines of a classification of the family: Proceedings of the Malacological Society of London, 29(2–3), 45–70. [ Links ]
Cox, L., Newell, N., Branson, C., Casey R., Cavan, A., Coogan, A., Dechaseaux, C., Fleming, C., Hass, F., Hertelin, L., Keen, A., Laroque, A., McAlester, A., Perkins, B., Puri, H., Smith, L., Soot–Ryen, T., Stenzel, H., Turner, R., Weir, J. , 1969, Systematic Descriptions, en Moore, R.C., Weir, J. (eds.), Treatise on Invertebrate Paleontology, Part N, Mollusca 6: Geological Society of America and University of Kansas Press, Lawrence, 225–868. [ Links ]
Cuvier, G., 1797, Tableau élémentaire de l'histoire naturelle des animaux: Paris, xvi+710 p., 14 pl. [ Links ]
Dickins, J.M., 1957, Lower Permian pelecypods and gastropods from the Carnarvon Basin, Western Australia: Bulletin of Bureau of Mineral Resources, Geology and Geophysics (Australia) 41, 1–75. [ Links ]
Férussac, A.E. de., 1822, Tableaux systématiques des animaux mollusques: Paris and London, 111 p. [ Links ]
Frech, F., 1891, Die devonischen Aviduliden Deutschlands: ein Beitrag zur Systematik und Stammesgeschichteder Zweischaler. K. Preussischen geolog. Landesanstalt, 261 pp. [ Links ]
Geinitz, H.B., 1866, Carbonformation und Dyas in Nebraska: Verhandlungen der K. Leopoldino–Carolinishe Deutschen Akademie der Naturforscher, 33, 1–91. [ Links ]
Girty, G.H., 1903, The Carboniferous Formations and faunas of Colorado: United States Geological Survey Professional Paper 16, 546 pp. [ Links ]
Girty, G.H., 1908, The Guadalupian fauna: United States Geological Survey Pofessional Paper 58, 651 pp. [ Links ]
Girty, G.H., 1909, Paleontology of the Manzano Group, en Lee, W.T., Girty, G.H., (eds.), The Manzano Group of the Rio Grande Valley, New Mexico: Bulletin of the United States Geological Survey, 389, 41–136. [ Links ]
Girty, G.H. 1910. The fauna of the phosphate beds of Park City formation in Idaho, Wyoming, and Utah: Bulletin of the United States Geological Survey, 436, 82 pp. [ Links ]
Girty, G.H. 1911. The Fauna of the Moorefield Shale of Arkansas: Bulletin of the United States Geological Survey, 439, 148 p. [ Links ]
Girty, G.H., 1912, On some new genera and species of Pennsylvanian fossils from the Wewoka Formation of Oklahoma: Annals of the New York Academy of Sciences, 21(2),119–156. [ Links ]
Goldfuss, A., 1836, Petrefacta Germaniae, Teil 2 (1834–1840): Dusseldorf, Arns and Company, 312 p. [ Links ]
González–Arreola, C., Villaseñor–Martínez, A.B., Corona–Esquivel, R., 1994, Permian fauna of the Los Arcos Formation, Municipality of Olinalá, State of Guerrero, México: Revista Mexicana de Ciencias Geológicas, 11(2), 214–221. [ Links ]
González, C.R., 1997, Late Carboniferous Bivalvia from Western Argentina: Geologica et Paleontologica, 31, 193–214. [ Links ]
Gray, J.E., 1824, Shells, en Parry, W.E. (ed.) A Supplement to the Appendix of Captain Parry's Voyage for the Discovery of a North–West Passage, in the years 1819–1820: London, 240–256. [ Links ]
Gray, J.E., 1847, A list of the genera of Recent Mollusca, their synonyms and types: Zoological Society of London Proceedings, 15, 129–219. [ Links ]
Hall, J., 1858, Lamellibranchiata of the Coal Measures, en Hall, J., Whitney, J.D. (eds.), Report on the Geological Survey of the State of Iowa, embracing the results of investigations made during portions of the years 1855, 1856 and 1857, Volume 1, Part 2 Paleontology, Albany, New York, 715–717. [ Links ]
Hall, J., 1883, Natural history of New York: New York Geological Survey, Paleontology, 5(1), 1–20. [ Links ]
Hind, W., 1901–1905, A monograph of the British Carboniferous Lamellibranchiata: Palaeontographical Society Proceedings, 2, 222 pp. [ Links ]
Hoare, R.D., Sturgeon, M.T., Kindt, E.A., 1979, Pennsylvanian marine Bivalvia and rostroconchia of Ohio: Ohio Geological Survey Bulletin 67, 77 p. [ Links ]
Hoover, P.R., 1981, Paleontology, taphonomy, and paleoecology of the Palmarito Formation (Permian of Venezuela): Bulletin of American Paleontology, 80, 1–138. [ Links ]
International Code of Zoological Nomenclature, 2000, 4a ed.: International Commission on Zoological Nomenclature, 156 pp. [ Links ]
International Stratigraphic Chart, 2010: International Commission on Stratigraphy, International Union of Geological Sciences, <http://stratigraphy.org>, consulta: 12 abril de 2011. [ Links ]
Isaacson, P.E., Dutro, J.T. Jr., 1999, Lower Carboniferous brachiopods from Sierra de Almeida, northern Chile: Journal of Paleontology, 73(4), 625–633. [ Links ]
Jablonski, D., 1986, Causes and consequences of mass extinction: a comparative approach, en Elliott, D.K. (ed.): Dynamics of mass extinction, New York, John Wiley and Sons, 183–229. [ Links ]
King, W. , 1844, On a new genus of Palaeozoic shells: Annals and Magazine of Natural History, 14(92), 313–317. [ Links ]
King, W., 1848, A Catalogue of the Organic Remains of the Permian Rocks of Northumberland and Durham, Newcastle upon Tyne. [ Links ]
King, W., 1850, A Monograph of the Permian Fossils of England: London, Palaeontographical Society Monographs, 258 pp. [ Links ]
Knight, J.B., 1931, The gastropods of the St. Louis, Missouri, Pennsylvanian outlier: the Subulitidae: Journal of Paleontology, 5(3), 177–229. [ Links ]
Knight, J.B., 1945, Some new genera of Paleozoic gastropoda: Journal of Paleontology, 19(6), 573–587. [ Links ]
Knight, J.B., 1953, Gastropoda, en Cooper, G.A., Dunbar, C.O., Duncan, H, Miller, A.K., Knight., J.B. (eds.), Permian fauna at El Antimonio, western Sonora, Mexico: Smithsonian Miscellaneous Collections, 119(2), 83–91. [ Links ]
Koninck, L.G. de, 1881, Faune du calcaire carbonífère de la Belgique, 3 partie, Gasteropodes: Annales du Musée Royal d'Historie Naturelle de Belgique, serie. Paleontologie, 6, 1–170. [ Links ]
Koninck, L.G. de, 1885, Faune du Calcaire Carbonifère, Lamellibranches: Annales du Musée Royal d'Historie Naturelle de Belgique, 11, 1–277. [ Links ]
Kues, B.S., 1991, Some gastropods from the Lower Wolcampian (basal Permian) Laborcita Formation, Sacramento Mountains, New Mexico: New Mexico Geological Society Guidebook 42, 221–230. [ Links ]
Kues, B.S., Batten, R.L., 2001, Middle Pennsylvanian gastropods from the Flechado Formation, north–central New Mexico: Journal of Paleontology 75(1 supp.), 1–95. [ Links ]
Lamarck, J.B. de, 1818–1819, Histoire naturelle des animaux sans vertèbres: Paris, 5, 612 p.; 6(1), 343 p. [ Links ]
Lebold, J.G., Kammer, T.W., 2006, Gradient analysis of faunal distributions associated with rapid transgression and low accommodation space in a Late Pennsylvanian marine embayment: Biofacies of the Ames Member (Glenshaw Formation, Conemaugh Group) in the northern Appalachian Basin, USA: Palaeogeography, Palaeoclimatology, Palaeoecology, 231, 291–314. [ Links ]
Lech, R.R., Milana, J.P., 2006, Nuevos registros de moluscos carboníferos en la sierra del Tontal, Precordillera de San Juan: Revista de la Asociación Geológica Argentina, 61(1), 57–62. [ Links ]
Linnaeus, C. (C. von Linné), 1758, Systema naturae per regna tria naturae: edición 10 reformada, Regnum animale, Estocolmo, 824 pp. [ Links ]
M'Coy, F., 1844, A Synopsis of the characters of the Carboniferous Limestone fossils of Ireland: Dublin, University Press, 207 pp. [ Links ]
M'Coy, F.,1851, Description of some new Mountain Limestone fossils: Annals and Magazine of Natural History, 2(7), 167–175. [ Links ]
McRoberts, C.A., Newell, N.D., 2001, A New Permian Myalinid Genus, Elversella, of West Texas: American Museum Novitates, 3311, 8p. [ Links ]
Meek, F.B., 1864, Remarks on the family Pteriidae(=Aviculidae) with descriptions of some new fossil genera: The American Journal of Science and Arts, 2(37),212–220. [ Links ]
Meek , F.B., 1874, New genus Euchondria Meek: American Journal of Science, 7(3), 445. [ Links ]
Meek, F.B., Hayden, F.V., 1865, Palaeontology of the Upper Missouri, Invertebrates, Part I: Smithsonian Contributions to Knowledge, 172, 135 p. [ Links ]
Meek , F.B., Worthen, A.H., 1866, Descriptions of invertebrates from the Carboniferous System: Illinois Geological Survey, 2, 145–411. [ Links ]
Meek, F.B., Worthen, A.H., 1869, Descriptions of new Carboniferous fossils from the western states: Philadelphia Academy of Natural Sciences Proceedings, 137–172. [ Links ]
Miller, S.A., 1877, The American Palaeozoic Fossils, a Catalogue of the Genera and Species: Cincinnati, 253 pp. [ Links ]
Miller, S.A., 1889, North American Geology and Paleontology: Cincinnati, Ohio, Western Methodist Book Concern Press, 793 pp. [ Links ]
Miller, S.A., 1891, Palaeontology: Indiana Geological Survey 17 th Annual Report, advance sheets, 611–705. [ Links ]
Montfort, P.D., 1808, Conchyliologie systématique, et classification methodique des coquilles: Tome I, Coquilles univalves, cloisoneés. Paris, F. Schoell, 409 pp. [ Links ]
Navarro–Santillán, D., Sour–Tovar, F., Centeno–García, E., 2002, Lower Mississippian (Osagean) brachiopods from the Santiago Formation, Oaxaca, México: Stratigraphic and tectonic implications: Journal of South American Earth Sciences, 15, 327–336. [ Links ]
Newberry, J.S., 1861, Paleontology, in Lieutenant Ives's Report on the Colorado River of the West: Gov. Printing Office, Washington, D. C., 116–129. [ Links ]
Newell, N.D., 1938, Late Paleozoic Pelecypods: Pectinacea: Kansas Geological Survey, 10(1), 1–123. [ Links ]
Newell, N.D., 1942, Late Paleozoic Pelecypods: Mytilacea: Kansas Geological Survey, 10(2), 1–115. [ Links ]
Newell, N.D., 1969, Classification of Bivalvia, en Moore, R.C., Teichert, C. (eds.), Treatise on Invertebrate Paleontology, part N, Mollusca 6(1): Geological Society of America and University of Kansas Press, Lawrence, N205–N218. [ Links ]
Newell, N.D., Boyd, D.W., 1975, Parallel evolution in early Trigoniacean Bivalves: Bulletin of the American Museum of Natural History 154, 57–162. [ Links ]
Newell, N.D., Boyd, D.W., 1985, Permian Scallops of the Pectinacean Family Streblochondriidae: American Museum Novitates, 2831,13 pp. [ Links ]
Newell, N.D., Boyd, D.W., 1995, Pectinoid Bivalves of the Permian–Triassic Crisis: Bulletin of the American Museum of Natural History 227, 95 pp. [ Links ]
Newell, N.D., La Rocque, A., 1969, Family Pterineidae Miller, 1877, en Moore, R.C. (ed.), Treatise on invertebrate paleontology, part N, Mollusca 6 (1): Geological Society of America and University of Kansas Press, N298–N302. [ Links ]
Nützel A., Erwin, D.H., Mapes, R.H., 2000, Identity and phylogeny of the Late Paleozoic Subulitoidea (Gastropoda): Journal of Paleontology, 74(4), 575–598. [ Links ]
Ochoa–Camarillo, H.R., 1996, Geología del anticlinorio de Huayacocotla en la región de Molango, estado de Hidalgo: México, D.F., Universidad Nacional Autónoma de México, Facultad de Ciencias, tesis de maestría, 91 pp. [ Links ]
Orbigny, A. de, 1843–1847, Mollusques, quatrième classe, Lamellibranchia, Paléontologie Francaise, 3, 807 pp. [ Links ]
Ortega–Guitérrez, F., Ruíz, J., Centeno–García, E., 1995, Oaxaquia—A Proterozoic microcontinent accreted to North America during the Late Paleozoic: Geology, 23, 1127–1130. [ Links ]
Pagani, M.A., 2000, Bivalvos del Pérmico Inferior de la Formación Bonete, Sierras Australes (Provincia de Buenos Aires, Argentina): Ameghiniana, 37, 301–320. [ Links ]
Pagani, M.A., 2004a, Los bivalvos carboníferos y pérmicos de Patagonia (Chubut, Argentina). Parte I: Introducción, familias Nuculidae y Malletidae: Ameghiniana, 41, 225–244. [ Links ]
Pagani, M.A., 2004b, Los bivalvos carboníferos y pérmicos de la Patagonia (Chubut, Argentina). Parte II: Familias Malletidae, Polidevciidae, Myalinidae e Inoceramidae: Ameghiniana, 41, 271–288. [ Links ]
Pagani, M.A. 2005, Los bivalvos carboníferos y pérmicos de Patagonia (Chubut, Argentina). Parte III: Familias Mytilidae, Pterineidae, Limidae, Leptochondriidae, Etheripectinidae, Euchondriidae y Streblochondriidae: Ameghiniana, 42, 579–596. [ Links ]
Pagani, M.A., 2006a, Los bivalvos carboníferos y pérmicos de Patagonia (Chubut, Argentina). Parte IV: Familias Avicu– lopectinidae, Deltopectinidae y Schizodidae: Ameghiniana, 43, 461–476. [ Links ]
Pagani, M.A., 2006b, Los bivalvos carboníferos y pérmicos de Patagonia (Chubut, Argentina). Parte V: Familias Trigoniidae, Permophoridae, Cardiniidae, Crassatellidae, Pholadomyidae, Sanguinolitidae y Megadesmidae. Conclusiones: Ameghiniana, 43, 539–556. [ Links ]
Pérez–Ramos, O., 1978; Estudio estratigráfico del Paleozoico Superior del Anticlinorio Huayacocotla, Veracruz: Instituto Mexicano del Petróleo, Subdirección Técnica de Exploración. Proyecto C–3016, 41 pp. [ Links ]
Phillips, J., 1836, Illustrations of the geology of Yorkshire, part 2, The Mountain Limestone district: London, John Murray, 253 pp. [ Links ]
Ponder, W.F., Lindberg, D.R., 1997, Toward a phylogeny of gastropod molluscs: an analysis using morphological characters: Zoological Journal of the Linnean Society, 119, 83–265. [ Links ]
Quiroz–Barroso, S.A., Perrilliat M.C., 1997, Pennsylvanian nuculoids (Bivalvia) from the Ixtaltepec Formation, Oaxaca, Mexico. Journal of Paleontology, 71(3), 400–407. [ Links ]
Quiroz–Barroso, S.A., Perrilliat, M.C., 1998, Pennsylvanian Bivalves from the Ixtaltepec Formation, Mexico: Journal of Paleontology, 72(6), 1011–1024. [ Links ]
Quiroz–Barroso, S.A., Pojeta, J., Jr., Sour–Tovar, F., Morales–Soto, S., 2000, Pseudomulceodens, a Mississippian Rostroconch from Mexico: Journal of Paleontology, 74(6), 1184–1186. [ Links ]
Rafinesque, C.S., 1815, Analyse de la Nature du Tableau de l'Univers et des Corps Organisés, in Rafinesque, C. S.,Palerme, from the Press of Jean Barravecchia, 225 p. [ Links ]
Ramsbottom, W.H.C., 1981, Permian, en McKerrow, W. S. (ed.), The ecology of Fossils, an illustrated guide: Cambridge Massachusetts, The MIT Press, p. 190. [ Links ]
Rosales–Lagarde, L., 2002, Estratigrafía y Geoquímica de la secuencia vulcano–sedimentaria paleozoica del noreste del Estado de Hidalgo: México, D.F., Universidad Nacional Autónoma de México, Posgrado en Ciencias de la Tierra, Tesis de Maestría, 89 pp. [ Links ]
Rosales–Lagarde, L., Centeno–García, E., Ochoa–Camarillo, Sour–Tovar, F., 1997, Permian volcanism in Eastern Mexico –preliminary report, en Segunda Convención sobre la Evolución Geológica de México y Recursos Asociados, Pachuca, Hidalgo, México: Instituto de Investigaciones en Ciencias de la Tierra, Universidad Autónoma del Estado de Hidalgo e Instituto de Geología, Universidad Nacional Autónoma de México, Libro–guía de las excursiones geológicas, Excursión 1, 27–32. [ Links ]
Rosales–Lagarde, L., Centeno–García, E., Dostal, J., Sour–Tovar, F., Ochoa–Camarillo, H., Quiroz–Barroso, S., 2005, The Tuzancoa Formation: Evidence of an early Permian submarine continental arc in East–central México: International Geology Review, 47(9), 901–919. [ Links ]
Sayre, A.N., 1930, The fauna of the Drum Limestone of Kansas and western Missouri: Bulletin of the Kansas Geological Survey, 19(2), 75–202. [ Links ]
Shen, S.Z., Shi, G.R., 2004, Capitanian (Late Guadalupian, Permian) global brachiopod paleobiogeography and latitudinal Diversity pattern: Palaeogeography: Palaeoclimatology, Palaeoecology 208, 235–262. [ Links ]
Silva–Pineda, A., 1987, Algunos elementos paleoflorísticos del Pérmico de la Región de Calnali, Hidalgo: Revista de la Sociedad Mexicana de Paleontología, 1 (1), 313–327. [ Links ]
Silva–Pineda, A., Buitrón–Sánchez, B.E., Arellano–Gil, J., Vachard, D., Ramírez, J., 2003, Permian continental and marine biota of South–Central Mexico: a synthesis, en Bartolini, R., Buffer, T., Blickwede, J. (eds.), The Circum–Gulf of Mexico and the Caribbean: Hydrocarbon habitats, basin formation, and plate tectonics: American Association of Petroleum Geologists Memoir 79, 462–475. [ Links ]
Sour–Tovar, F., Pérez–Huerta, A., Quiroz–Barroso, S.A, Centeno–García, E., 2005, Braquiópodos y trilobite del Pérmico Inferior del noroeste del Estado de Hidalgo, México: Revista Mexicana de Ciencias Geológicas, 22(1), 24–35. [ Links ]
Sowerby, J.de C., 1821–1825, The Mineral Conchology of Great Britain, London, 5, 168 p. [ Links ]
Sowerby, J. de C., 1829, Transactions of the Geological Society of London, 3,1–120. [ Links ]
Stanley, S.M., 1970, Relation of shell form to life habits in the Bivalvia (Mollusca): Geological Society of America Memoir 125, 1– 296. [ Links ]
Sterren, A.F., 2004, Bivalvos pérmicos de la Formación Tupe en la quebrada de la Herradura, provincia de San Juan: Ameghiniana, 41, 57–74. [ Links ]
Sterren, A.F., 2005, Bivalvos carboníferos de la Formación La Capilla en el área de Las Cambachas, provincia de San Juan: Ameghiniana, 42, 209–219. [ Links ]
Stevens, R.P., 1858, Descriptions of new arboniferous fossils from the Appalachian, Illinois, and Michigan coal fields: American Journal of Science, 25, 258–265. [ Links ]
Stewart, J.H., Blodgett R.B., Boucot A.J., Carter J.L., López, R., 1999, Exotic Paleozoic strata of Gondwana provenance near Ciudad Victoria, Tamaulipas, México, in Ramos, V.A., and Keppie, J.D., (eds.), Laurentia–Gondwana Connections before Pangea: Geological Society of America Special Paper 336, p. 227–252. [ Links ]
Stoliczka, F.,1870–1871, Cretaceous fauna of southern India: 3. the Pelecypoda: Memoirs of the Geological Survey of India: Palaeontologia Indica, 537 pp. [ Links ]
Termier, G., Termier, H., 1950, Paléontologie marocaine. 2, Invértebrés de l'ere primaire. Fascicule 3, Mollusques: Paris, Hermann, 246 pp. (Service Géologique du Maroc, Notes et Memoires, 78). [ Links ]
Thomas, E.G., 1940, Revision of the Scottish Carboniferous Pleurotomariidae: Geological Society of Glasgow Transactions, 20, 30–72. [ Links ]
Vachard, D., Arellano, J., Buitrón, B., Flores de Dios, A., Yussim, S., 1997, Datación del Sakmariano?–Artinskiano (Pérmico Inferior) en Pexmuco, Hidalgo; consecuencias paleogeográficas (resumen), en Segunda Convención sobre la evolución geológica de México y Recursos Asociados, Simposia y Coloquio, Instituto de Investigaciones en Ciencias de la Tierra: Pachuca, Hidalgo, Universidad Autónoma del Estado de Hidalgo, Instituto de Geología, Universidad Nacional Autónoma de México, 37–41. [ Links ]
Verneuil, E. de, Murchison, R.I., 1844, Note sur les équivalents du systéme Permien en Europe, suivie d'un coup d'oeil general su l'ensemble de ses fossiles, et d'un tableau des espécies: Société Géologique de France Bulletin, 1, 475–517. [ Links ]
Verneuil, E. de, 1845, Mollusques, p. 37–376, en Murchison, R.I. Verneuil, E. de, Keyserling, A. de (eds.), Géologie de la Russie d' Europe, 2(3). [ Links ]
Vermeij, G.J., 1995, Economics, volcanoes, and Phanerozoic revolutions: Paleobiology, 21, 125–152. [ Links ]
Wagner, P.J., Erwin, D.H., 2006, Patterns of convergence in general shell from among Paleozoic gastropods: Paleobiology, 32(2), 316–337. [ Links ]
Waller, T.R., 1978, Morphology, morphoclines and a new classification of the Pteriomorphia (Mollusca: Bivalvia), p. 345–365, en Yonge, C. M., Thompson, T. E. (eds.), Evolutionary Systematics of bivalve molluscs: Philosophical Transactions of the Royal Society of London, B. Biological Sciences, 284, 345–365. [ Links ]
Waller, T.R., 1998, Origin of the molluscan class Bivalvia and a phylogeny of major groups, en Johnston, P.A., Haggart, J.W. (eds.), Bivalves: An Eon of Evolution: Calgary, University of Calgary Press, Paleontological Studies honoring Norman D. Newell, 1–45. [ Links ]
Waterhouse, J.B., 2008, Aspects of the evolutionary record for fossils of the bivalve subclass Pteriomorphia Beurlen: Earthwise, 8, 1–219. [ Links ]
Weber, R., 1997, How old is the Triassic Flora of Sonora and Tamaulipas and news on Leonardian floras in Puebla and Hidalgo, Mexico: Revista Mexicana de Ciencias Geológicas, 14(2), 225–243. [ Links ]
Wenz, W., 1938–1944, Gastropoda. Allgemeiner Teil und Prosobranchia, en Schindewolf, O.H. (ed.): Berlin, Handbuch der Paläozoologie, Band 6, 480 pp. [ Links ]
Worthen, A.H., 1890, Description of fossil invertebrates: Illinois Geological Survey, 8, 69–154. [ Links ]
Yancey, T.E., 1975, Permian marine biotic provinces in North America: Journal of Paleontology, 49(4), 758–766. [ Links ]
Yochelson, E.L., 1956, Permian gastropoda of the Southwestern United States: I. Euomphalacea, Trochonematacea, Pseudophoracea, Anomphalacea, Craspedostomatacea, Platyceratacea: Bulletin of the American Museum of Natural History, 110(3), 173–276. [ Links ]

