
Sistemas de memoria: reseña histórica, clasificación y conceptos actuales. Segunda parte: Sistemas de memoria de largo plazo: Memoria episódica, sistemas de memoria no declarativa y memoria de trabajo
Memory systems: Historical background, classification and current concepts. Part two
Paul Carrillo–Mora1
1 Laboratorio de Aminoácidos Excitadores, Instituto Nacional de Neurología y Neurocirugía Manuel Velasco Suárez.
]]> Correspondencia:
]]> Abstract
According to recent evidence, memory can be conceptualized as a series of subsystems working together to reach the same final goal. In the present time, most authors coincide in proposing a main division or long–term memory systems in mammals. This division separates memory in two categories: declarative and not declarative, often conceptualized also as explicit and implicit memories, respectively, although the latter terms are more appropriate to describe the role of consciousness during the development of codification and recovery tasks in regard to the information of each system. Declarative memory, as indicated by its name, includes a variety of memories implicating information that can be verbalized and effectively transmitted from person to person. This type of memory is conceptualized as conformed by two distinct memory sub–categories sometimes collaborating and coinciding in the use of memory. The first of these sub–categories is semantic memory, the one including the information about concepts and precise facts, and it is frequently defined as «general knowledge». It also makes reference to conceptual information lacking from the temporal–spatial frame (this issue is detailed in the first part of this paper). The second sub–category is episodic memory, which refers to the memory for personal experienced events, or the memory used for «what?, where?, and when?», in regard to the occurrence of a given personal event. Altogether, autonoesis (the conscious concept of a personally experienced event), subjective temporal consciousness (or subjective temporal frame about when a given event occurred), and self–consciousness are considered indispensable for the definition of episodic memory. In spite of these characteristics, there is yet controversy on whether this type of memory could be observed in animals. In this regard, some researchers have demonstrated that the processes needed to define episodic memory are effectively employed by non–human primates and rodents.
In regard to the functional anatomy of episodic memory, an active role of the temporal lobe in the codification of some components of this memory is known from long time ago. Experimental studies employing hippocampal electro–physiological recordings have demonstrated that this region participates in two main components of episodic memory: i) the strong links of the subject with the environment or the spatial context, and ii) the temporal organization of the stored information. Some studies have clearly demonstrated the important participation of specific hippocampal cell areas (CA1 and CA3) for the resolution of behavioral tasks requiring episodic memory. However, clinical studies employing functional imaging have shown a considerable diversity of cortical areas as involved in the several processes of codification or recovery of episodic information, going from prefrontal medial and left ventrolateral regions to medial and lateral temporal regions, retrosplenial cortex, posterior cingulus posterior, and even the cerebellum.
Non–declarative memory systems
Non–declarative memory contains different categories: procedural memory, priming, associative learning (classic and operant conditioning) and non–associative learning (habituation and sensitization): a) Procedural memory refers to the storage and recovery of information on motor skills, or «know how to do» distinct tasks. Although this type of memory is considered as part of those memory systems often leading to unconscious learning, the relationship between procedural memory and consciousness is more complex. This is due in part to the fact that there is no evidence of a real association between the voluntary (conscious) desire of movement and the activation of motor brain areas. In addition, it has been demonstrated that movements consciously activated to start a given task may difficult the performance of the same task. Moreover, learning of motor skills exhibits a particular characteristic known as consolidation or off–line stabilization. This term refers to those motor skills exhibiting an improvement in performance during the interval of two practice sessions, meaning that the performance is improved if behavior is re–analyzed after an interval of rest typically occurring along the day or during sleep periods. This supports the theory that during sleep periods there is a recapitulation of events taking place during the day, thus favoring the strengthening of neuroplastic mechanisms involved in motor learning. b) Priming is a type of implicit memory not requiring any conscious recollection of previous experiences, and it shares some features with procedural and semantic memories. Similar to procedural memory, priming implies an increase of skills, but in this case, perceptual skills. It is also similar to semantic memory in terms of the involvement of cognitive representations of outdoor environment, and its representation is more cognitive than behavioral. The general characteristics of priming are: i) it is related with the perceptual identification of objects in general terms, including words or concepts; ii) its neuronal substrate is not depending on those brain regions needed for episodic or semantic memories; iii) it is developed early in life and its capacity remains stable all the time; iv) its activity is not related with consciousness and its function is not sufficient to recall a previous experience; v) it is relatively immune to the effects of drugs affecting the other memory systems; vi) its information is distributed in multiple representations of specific words and objects; and vii) the access to this representations is highly specific to each category of objects. Finally, among short–term memory systems, the concept of working memory proposed by Baddeley and Hitch is still accepted in our days, and is characterized by the transitory storage and administration of all kind of information useful for the achievement of a specific task. This system is based on three main components: i) a control system with limited attention capacity, also known as Executive Central Component; ii) the Phonologic Circuit, based on sound and language; and iii) the Visual–Spatial Scheme. The phonologic circuit is responsible for the transitory storage of language information based on sound and it is proponed to play an active role for learning of language. The Visual–Spatial Scheme is important for the visual–spatial representation of objects and their features, which in turn is relevant for the integration of concepts. The Executive Central Component is in charge of controlling the attention and the access to the other two subsidiary components. The knowledge on how the memory systems are functioning is and will remain in constant expansion, given the obvious contribution of recent studies on molecular biology and functional neuroimaging.
Key words: Memory, semantic, episodic, procedural, working memory.
Resumen
La memoria episódica (ME) forma parte de los sistemas de memoria declarativa y es considerada como aquella de los sucesos experimentados de forma personal, o sea la memoria del «¿qué?», «¿dónde?» y «¿cuándo?» ocurrió cierto suceso de la experiencia personal. La triada de autonoesis (es decir, la conciencia de que un suceso le ocurrió personalmente), conciencia temporal subjetiva (o el marco temporal subjetivo de cuando ocurrió un suceso y en qué orden) y la conciencia de sí mismo, son consideradas como requisito indispensable para la definición de la memoria episódica. Hasta hace algunos años la memoria episódica era típicamente considerada como una habilidad exclusiva del ser humano, debido a los procesos cognitivos con los que se asocia, sin embargo, recientemente se ha comprobado que otras especies animales tienen y utilizan eficazmente un sistema análogo al de la ME para resolver diversas tareas de memoria en el laboratorio. Estos modelos experimentales de ME han impulsado la generación de conocimientos acerca de este sistema. La participación del lóbulo temporal, y en especial del hipocampo, en la ME es conocida desde hace tiempo, sin embargo se han identificado diversas áreas cerebrales implicadas en la recuperación y codificación de la información episódica. Una de las más estudiadas actualmente es la corteza parietal posterior, pero existen otras cuyo rol preciso en este tipo de memoria aún no se ha esclarecido. Por otro lado, la otra gran categoría central de la memoria: la no declarativa, contiene diversas subcategorías heterogéneas en su contenido y función, como son: la memoria procedimental, el priming, el aprendizaje asociativo (condicionamiento clásico y operante) y el aprendizaje no asociativo (habituación, deshabituación y sensibilización) mediado por vías reflejas. La memoria procedimental se refiere al almacenamiento y recuperación de información sobre habilidades motoras, es decir, el aprendizaje de «saber cómo hacer» distintas tareas. La memoria procedimental tiene relaciones complejas con la «conciencia», se ha demostrado que presenta un proceso de consolidación off–line, es decir que la información se consolida mejor cuando existe un periodo de reposo entre las sesiones de entrenamiento motor, sobre todo si durante este descanso ocurre el sueño. Existe una serie de estructuras tanto corticales como subcorticales que participan en diversas funciones de la memoria procedimental y cuya lesión selectiva produce distintos trastornos de las funciones motoras, como la parálisis, la apraxia, el temblor, los movimientos coreicos, las distonías, las dismetrías o la ataxia. El priming es un tipo de memoria implícita que no requiere de ninguna recolección consciente de experiencias previas, y que comparte algunas características con la memoria procedimental pero también con la semántica. Se trata de una variedad de memoria en la cual se propone que participan áreas neocorticales de asociación visual. Finalmente, dentro de los sistemas de memoria de corto plazo se encuentra el concepto de memoria de trabajo, propuesto por Baddeley y Hitch, que sigue teniendo aceptación en la actualidad y que es considerado como un sistema encargado de almacenar y administrar transitoriamente toda la información (de distintas modalidades) que se encuentra actualmente en uso para la realización de una tarea específica. Esta memoria de trabajo se conceptualiza como conformada por un componente ejecutivo central y dos sistemas subsidiarios: el circuito fonológico (encargado entre otras cosas del reforzamiento verbal implicado en el aprendizaje del lenguaje) y el esquema visuo–espacial que estaría encargado del almacenamiento transitorio de la información espacial del ambiente externo. El conocimiento científico de la memoria continúa en expansión cotidiana, pero aún estamos lejos de conocer todos los detalles acerca de su funcionamiento desde el nivel molecular hasta el anatómico macroscópico. Sin duda los estudios moleculares y de imagen funcional ofrecerán en el futuro una panorámica más amplia al respecto.
Palabras clave: Memoria, semántica, episódica, procedimental, memoria de trabajo.
]]>INTRODUCCIÓN
Los sistemas de memoria de largo plazo en los mamíferos se consideran conformados por dos grandes categorías de información, la declarativa y la no–declarativa.1 La memoria declarativa se compone por dos variedades de memoria: la semántica (que ha sido expuesta en la primera parte de este artículo) y la episódica, que comprende una memoria compuesta por información sensorial de muy distintas variedades sobre un marco temporal y espacial definido acerca de hechos que ocurrieron en el pasado personal. La riqueza de información es la característica de esta variedad de memoria que ha dado origen y sentido a la expresión coloquial: «recordar es volver a vivir».
MEMORIA EPISÓDICA
Es aquella de los sucesos experimentados personalmente o el recuerdo del «¿qué?», «¿dónde?» y «¿cuándo?» ocurrió cierto hecho de la experiencia personal.2 Según esta definición, la memoria episódica se comporta como una función asociativa entre distintas modalidades de información (visual, espacial y temporal) que origina un estímulo con una configuración compleja que denominamos «suceso».3 El componente de la experiencia personal es importante para la memoria episódica, y éste se refiere a que los eventos recordados deben haber sido experimentados personalmente (por lo que también es referida como memoria autobiográfica). Pueden recordarse otros eventos que no hayan sido experimentados de primera mano (por ejemplo, sucesos históricos), pero éstos no son considerados como parte de la memoria episódica. Una de las características importantes de este tipo de memoria es lo que Tulving denominó «nivel de conciencia»:4,5 la memoria episódica necesita que la persona sea consciente de que lo que recuerda sea algo que le ocurrió personalmente (conciencia autonoética), pero esto a su vez requiere de lo que llamó «recolección consciente»; es decir, es necesario que el individuo sea capaz de discernir entre eventos pasados que no vivió personalmente, los de su pasado reciente o los que sólo le resultan «familiares» (esta noción de familiaridad se refiere a que el individuo reconoce cierta información del evento específico pero no puede identificar con claridad la fuente original de tal información).4,5 Otro componente importante de la memoria episódica es la ubicación de los eventos en un marco temporal subjetivo, lo cual provee del conocimiento sobre la secuencia de eventos que han ocurrido en el pasado (memoria retrospectiva), así como el orden preciso en que ocurrieron éstos (memoria de orden temporal).5 Todo esto requiere además de un conocimiento sobre sí mismo como una entidad distinta del resto del mundo.3 Esta triada de autonoesis, conciencia temporal subjetiva y conciencia de sí mismo son consideradas como requisito indispensable para la memoria episódica.6 A pesar de estas características bien definidas, la memoria episódica comparte algunas características con la memoria semántica: i) Ambas representan sistemas grandes y complejos de memoria que tienen una capacidad ilimitada (hasta donde sabemos) de almacenamiento de diversos tipos de información (a diferencia de la memoria de trabajo que tiene una capacidad limitada de información, o los sistemas de representación perceptual que manejan un solo tipo de información). ii) Ambos sistemas manipulan información cognitiva (declarativa y representacional), y en ambos sistemas su contenido se puede describir en términos de objetos y sus relaciones. iii) Ambos sistemas adquieren información de forma similar. Más aún, el acceso de información a uno de los dos sistemas no puede desligarse del acceso al otro sistema al mismo tiempo. iv) Puede accederse a la información almacenada en ambos sistemas de forma flexible, pudiendo ser evocada a través de distintas vías; ambos sistemas utilizan códigos de marcaje específicos y su recuperación exitosa depende no sólo del tipo de estímulo almacenado sino del contexto semántico o episódico con el que fue codificado inicialmente.7 Aun con estas coincidencias, la ciencia actual considera que ambos sistemas son esencialmente distintos, aunque como ya se mencionó, ambos trabajan en la práctica inexorablemente unidos.
El estudio y análisis de la memoria episódica en los seres humanos no ha sido sencillo. Desde antaño se consideraba que una tarea simple, como la identificación y recuerdo posterior de una serie de estímulos (por ejemplo visuales) era suficiente para explorar la memoria episódica; sin embargo, es claro que estas tareas pueden ser resueltas sin la necesidad de realizar una verdadera «recolección consciente» de eventos previos, sino únicamente basándose en una noción de «familiaridad» de los estímulos (familiaridad se refiere a que la persona puede saber que un estímulo ya ha sido experimentado aun sin que tenga un recuerdo consciente o claro de haberlo aprendido con anterioridad o bajo qué circunstancias). De la misma forma, es posible que la persona recuerde hechos de su pasado sin la necesidad de realizar un verdadero «viaje mental al pasado», lo cual complica la interpretación de los resultados obtenidos en estas pruebas.6,7 Estas discrepancias en la resolución de una misma tarea explican por qué los estudios clínicos de imagen funcional pueden demostrar activaciones diferentes en tareas de memoria episódica vs. autobiográfica, términos que generalmente se consideraban como sinónimos.8 Obviamente, si definir y estudiar la memoria episódica en seres humanos ha resultado polémico hasta la actualidad, el proponer que ésta pueda existir, modelarse y estudiarse en otro tipo de animales resultó aún más complicado.3 Para algunos autores este tipo de memoria debería ser por definición exclusiva de los seres humanos pues requiere de una serie de habilidades cognitivas y de expresión que sólo son factibles en el contexto humano.9,10 Sin embargo, cada vez más autores apoyan el hecho de que otros animales tienen y de hecho utilizan eficazmente procesos cognitivos semejantes a los requeridos por la memoria episódica, tales como la metacognición,11 la recolección consciente,12 la memoria de orden temporal, etc.13,14 Debido a lo anterior, se han desarrollado diversos paradigmas que intentan explorar estos sistemas integrados de memoria de objetos, lugares y de orden temporal para el análisis experimental de la memoria episódica;15,16 paradigma experimental con el que no se contaba hasta hace algunos años y cuyo desarrollo resultaba urgente debido a la diversidad de patologías humanas en las que se ve afectado este sistema y que requerían de un abordaje experimental adecuado para su estudio3 (cuadro 1).
En cuanto a la anatomía funcional de la memoria episódica, es conocida desde hace tiempo la participación del lóbulo temporal en la codificación de algunos de sus componentes17 Los estudios experimentales con registros electrofisiológicos hipocampales han mostrado que esta región participa en dos componentes fundamentales de la memoria episódica: las fuertes asociaciones del individuo con el ambiente o su contexto espacial, y la organización temporal de la información almacenada.18–20 Incluso algunos estudios han demostrado la importante participación de regiones celulares especificas dentro de la formación del hipocampo (CA1 y CA3) en la resolución de las tareas conductuales que requieren de la memoria episódica.21 Sin embargo, los estudios clínicos de imagen funcional han mostrado una gran diversidad de áreas corticales involucradas en los diversos procesos de codificación o recuperación de la información episódica, los cuales van desde las áreas prefrontales mediales y ventro–laterales izquierdas, hasta las regiones medial y lateral temporal, la corteza retroesplenial, el cíngulo posterior e incluso el cerebelo.22–24 Uno de los aspectos recientemente estudiados es lo que se conoce como «pensamiento episódico futuro» que se conceptualiza como la habilidad de proyectarse a sí mismo hacia el futuro dentro del marco de un evento previamente experimentado; se ha demostrado que esta capacidad se desarrolla alrededor de los 3–4 años de edad pero las características que la diferencian de la memoria episódica y su sustrato anatomo–funcional aún no han sido estudiadas por completo25. Otro aspecto que requiere de esclarecimiento futuro es la participación que tiene la corteza parietal en la memoria episódica. Los estudios actuales han demostrado una participación importante en la recuperación episódica de información antigua más que de la reciente, pero puede ser que también participe en los procesos de codificación inicial de la información.26,27
]]>MEMORIA NO DECLARATIVA
Memoria procedimental
La memoria procedimental se refiere al almacenamiento y recuperación de información sobre las habilidades motoras; es decir, el aprendizaje relacionado a «saber como hacer» distintas tareas.28 La definición de memoria procedimental enfrentó desde su inicio diversas dificultades conceptuales según los esquemas clásicos de clasificación de los sistemas de memoria:29 primero, se encuentra incluida en una categoría de sistemas de memoria que la clasifica con base en las características de que carece (es decir, no declarativa) más que hacer referencia a lo que sí contiene (habilidades motoras), lo cual a su vez la coloca dentro de los sistemas de memoria que aparentemente no requieren de auto conocimiento o conciencia (memoria implícita o inconsciente).2 Sin embargo, los conocimientos actuales sobre las propiedades de la memoria procedimental han enfatizado la necesidad de una nueva clasificación en la que se considere el involucramiento de los niveles de atención, además de la participación de la conciencia30 (figura 1).
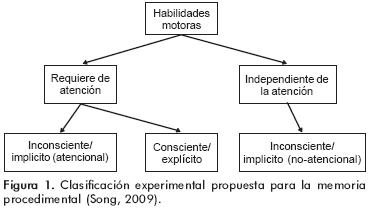
La relación entre conciencia y actividad motora es compleja; los estudios experimentales han demostrado que la noción de la planeación de movimiento en realidad no tiene una traducción en la actividad cerebral, sugiriendo así que existe activación neuronal incluso antes de que exista planeación «consciente» de movimiento por parte del individuo.31 Por ello se considera que este «deseo consciente de movimiento» es en realidad una ilusión creada por el cerebro en un afán de establecer una relación causal entre pensamiento y movimiento.28 Diversos estudios han evidenciado que la «conciencia» en la realización de una tarea que implique una secuencia de movimientos puede afectar negativamente el desempeño y la velocidad de realización de la misma; puesto en otras palabras: la acción «consciente» de una secuencia de movimientos puede entorpecer más que ayudar a la realización de la misma.32 De esta forma podemos decir que la relación entre memoria procedimental y conciencia es difícil de definir con precisión, por lo que algunos autores han propuesto un sistema de clasificación basado en la atención o en la intención con la que se practica la tarea más que en la toma de conciencia30 (figura 1). Además, dicho sistema debe incluir otras características particulares que se han documentado del proceso de consolidación motora, entre las cuales destacan las siguientes: la consolidación o estabilización off–line;29 este término se refiere a que las tareas motoras exhiben una mejoría en su desempeño durante el intervalo entre dos sesiones de práctica, es decir, el desempeño mejora si se re–analiza la conducta después de un intervalo de descanso que puede ocurrir durante el día o durante el sueño.33 Por ejemplo, en tareas de discriminación perceptual se ha observado un incremento de entre 15–20% en el rendimiento después de una noche de sueño.34 Esta mejoría no guarda relación con la duración total del sueño sino con la duración de una fase particular de éste (fase II del sueño no–MOR) o de una combinación especial de sus componentes.34 Se propone que lo anterior apoya la teoría de que durante el sueño existe una recapitulación de los eventos ocurridos durante el día, favoreciendo así el fortalecimiento de los mecanismos neuroplásticos implicados en el aprendizaje35 Las estructuras cerebrales relacionadas en el aprendizaje motor en el ser humano son diversas, según lo han indicado múltiples estudios clínicos y de imagen funcional: i) Corteza prefrontal: esta región es activada en las etapas iniciales de aprendizaje motor explícito. Se ha propuesto que la corteza prefontal izquierda está especializada en la codificación y la derecha en la recuperación de la información motora.36 ii) Corteza del cíngulo: se ha implicado en la retroalimentación sensorial de la corteza prefrontal, así como con la selección de una respuesta basada en el resultado esperado de una acción previa.37 iii) Área motora pre–suplementaria y suplementaria (AMPS y AMS): ambas están activadas en las etapas tempranas del aprendizaje, pero la AMS incrementa su actividad con la práctica de la tarea y está asociada con el desempeño de secuencias de movimientos, sobre todo los que requieren de ritmo o uso simultáneo de varias extremidades.38 iv) Área premotora: especialmente la del lado derecho se ha relacionado con la asociación de la información espacial y las respuestas motoras apropiadas; además existe evidencia de que en la corteza premotora ventral existen las que se han llamado «neuronas en espejo», que son células que descargan tanto cuando se realiza una acción específica como cuando se observa a otro individuo realizar la misma tarea u otra similar.39 Se ha propuesto que estas neuronas pueden estar relacionadas con el aprendizaje motor vicario (por imitación).39 v) Área motora primaria: Su activación es contralateral a las extremidades usadas en la tarea y se asocia con la velocidad y dirección del movimiento voluntario.40 vi) Estructuras subcorticales: Cerebelo, el cual está relacionado con la retroalimentación sensorial (propioceptiva, visual y vestibular) del movimiento que se está realizando en el momento actual, y la detección y corrección de errores en la ejecución de tareas;41 Ganglios Basales: En las etapas iniciales de la adquisición se ha reportado activación del caudado anterior, pero a los ganglios basales se les ha atribuido la función global del almacenamiento de secuencias de movimiento engramadas durante el aprendizaje implícito.42
PRIMING
El priming es un tipo de memoria implícita que no requiere de ninguna recolección consciente de experiencias previas, y que comparte algunas características con la memoria procedimental, pero también con la memoria semántica.43 Al igual que la memoria procedimental, el priming significa un incremento de habilidades, pero en este caso perceptuales. Por otro lado, también se asemeja a la memoria semántica en que involucra representaciones cognitivas del mundo exterior y su expresión es más cognitiva que conductual.5,43 En las pruebas típicas para el estudio de las habilidades perceptúales se dice que se ha demostrado el priming si la probabilidad de identificación de objetos actualmente presentados se incrementa con la visualización previa de dichos estímulos (ambas presentaciones separadas por un intervalo de tiempo que puede ser de minutos, horas o meses) o si el tiempo de latencia en la respuesta de identificación ha disminuido; en ambos casos se hace la comparación con respecto a un grupo control sin presentación previa de estímulos.5
]]> Existe evidencia neuropsicológica que demuestra que el priming significa un incremento en las operaciones de correlación cognitiva de un sistema general de representación perceptual (SRP), y esta evidencia tiene su origen en la variedad de disociaciones del priming que se han observado respecto a los otros sistemas de memoria explícita. Al mismo tiempo estos hallazgos también demuestran algunas de las propiedades generales de este SRP: i) Está relacionado con la identificación perceptual de objetos en general, incluyendo palabras. ii) El sustrato neuronal del SRP no es dependiente de las regiones cerebrales necesarias para la memoria episódica y semántica. iii) Se desarrolla de forma temprana en la vida y su capacidad se mantiene estable a lo largo de ella. iv) Su operación está desconectada de la conciencia y su funcionamiento no es suficiente para recordar la experiencia previa. v) Es relativamente inmune a los efectos de las drogas que afectan los otros sistemas de memoria. vi) La información en el SRP está distribuida en múltiples representaciones de palabras particulares y objetos; y vii) El acceso a estas representaciones es hiperespecífico. A pesar de estas diferencias importantes con el resto de los sistemas de memoria el priming también demuestra coincidencias con ellos, entre ellas que el priming se beneficia de las repeticiones, demuestra olvido en relación con el tiempo y varía según las condiciones de codificación y recuperación de la información. Estas discrepancias y coincidencias ponen de relieve que el priming forma parte de un sistema de memoria independiente de los demás, pero que al mismo tiempo comparte algunas características generales con ellos, por lo que la cuestión acerca de un sistema unitario vs. múltiples sistemas de memoria aún permanece sin resolver.5,43,44
SISTEMAS DE MEMORIA DE CORTO PLAZO
Memoria de trabajo
Desde 1949, Donald Hebb propuso una distinción de índole fisiológica entre la memoria de corto plazo (MCP) y la de largo plazo (MLP);45 la primera dependía de una activación eléctrica temporal y la segunda de un verdadero crecimiento o modificación neuronal. A raíz de esto, una gran diversidad de modelos fueron propuestos para explicar el funcionamiento diferencial de ambos tipos de memoria. Uno de los modelos más influyentes fue el planteado por Baddeley y Hitch en 1974.46 Estos autores propusieron un modelo llamado «memoria de trabajo» que todavía es aceptado en la actualidad. La memoria de trabajo es considerada como un sistema encargado de almacenar y administrar transitoriamente toda la información (de distintas modalidades) que se encuentra actualmente en uso para la realización de una tarea específica.46 Este sistema se basa en tres componentes principales: i) Un sistema de control con capacidad atencional limitada que denominaron: Componente Central Ejecutivo, el cual era asistido por dos sistemas subsidiarios de almacenamiento. ii) El circuito fonológico, que está basado en sonido y lenguaje. iii) El esquema visuo–espacial (figura 2).46,47

CIRCUITO FONOLÓGICO
El circuito fonológico contiene teóricamente dos componentes: un almacén fonológico que puede retener información por algunos segundos antes de que se olvide, y un sistema articulatorio de reforzamiento de repetición, análogo al de la repetición verbal48 (figura 3). Así, mediante este mecanismo la información puede ser «refrescada» con la repetición verbal; sin embargo la memoria inmediata es limitada porque la articulación ocurre en tiempo real, de modo que al incrementar el número de estímulos consecutivos por recordar, llega un momento en que el primero ha sido olvidado antes de poder ser repetido.47
]]>
Según los estudios neuropsicológicos, para el recuerdo de palabras en el corto plazo resultan más importantes sus propiedades acústicas o fonológicas (sonido) que las semánticas (significado), mientras que al explorar la memoria de largo plazo ocurre exactamente lo contrario.49–52
Este modelo de dos componentes para el circuito fonológico se apoya también en evidencias neuropsicológicas ya que se han encontrado pacientes que presentan déficits articulatorios fonológicos selectivos en el corto plazo, sin alteración de la funcionalidad general del resto del lenguaje, y en los cuales al cambiar la presentación de la información por vía visual y no auditiva, desaparece el defecto fonológico, sugiriendo así que esta estrategia visual evita el almacén fonológico defectuoso.53 En estos pacientes se han documentado típicamente lesiones del área 40 de Brodmann en el lóbulo parietal inferior. Además, el mecanismo de repetición verbal no depende de la capacidad de los órganos periféricos de control de la articulación fonatoria; es decir, que se ha encontrado preservado incluso en pacientes anártricos (incapaces de articular palabras). En contraste, en pacientes con trastornos práxicos del lenguaje en los cuales se ha perdido la capacidad de la representación motora interior, se ha observado una reducción de la capacidad de memoria de repetición.53 La evidencia sobre la existencia del circuito fonológico parece concluyente, aunque ¿cuál es la función de éste? Se ha propuesto que este circuito tiene la finalidad de facilitar la adquisición del lenguaje y la evidencia que apoya esta teoría proviene de que se han observado déficits en la adquisición de lenguaje nuevo en pacientes con defectos selectivos de este circuito fonológico.54 Por otro lado, se ha demostrado que algunos factores que alteran el circuito fonológico (como la supresión articulatoria, la similitud fonológica y la longitud de las palabras) también alteran el aprendizaje de un lenguaje extranjero, pero no alteran el aprendizaje de pares asociados de palabras no relacionadas del lenguaje materno.55,56 Incluso esta capacidad del circuito fonológico se utiliza exitosamente como un buen predictor de la habilidad futura de niños y adultos para aprender un segundo idioma.56
La forma en la que se propone que este circuito facilita el aprendizaje comprende dos vías: i) el almacén fonológico aporta una representación temporal relativamente extensa para nuevas secuencias fonémicas y ii) el sistema articulatorio puede facilitar el aprendizaje por medio del reforzamiento mediante repetición. Este proceso de facilitación ocurre de inmediato con las secuencias fonéticamente regulares pero puede requerir de exposiciones posteriores cuando las secuencias son irregulares o extrañas.57
ESQUEMA VISUO–ESPACIAL
De la misma forma que su contraparte verbal, la memoria de trabajo visual–espacial tiene una capacidad limitada. En los estudios clínicos típicamente es capaz de conservar entre 3–4 objetos. El recuerdo de objetos está compuesto por varias características como el color, la forma y la localización. Los estudios neuropsicológicos han demostrado la necesidad de distinguir la memoria visual de la memoria espacial, ya que se han reportado pacientes con disociaciones de ambas, es decir, con afección de la memoria visual sin alteración de la espacial y viceversa.58 Se piensa que la función del esquema visuo–espacial, como sucede con el circuito fonológico, es la de facilitar el aprendizaje, en este caso de tipo semántico, proporcionando información acerca de la apariencia de los objetos y la manera de usarlos, ayudando así a comprender visualmente sistemas complejos (como por ejemplo, el de alguna maquinaria), así como para la orientación espacial y el conocimiento geográfico.47 Algunos autores han propuesto una división del esquema visuo–espacial, distinguiendo el componente de almacén visual, el refugio visual y el proceso dinámico de recuperación–reforzamiento, aunque se trata de un campo aún en investigación debido a que sus funciones no están bien conocidas del todo.47
Componente ejecutivo central
El componente ejecutivo central es sin duda el más importante de los tres pero es el menos comprendido de todos los componentes de la memoria de trabajo. En la descripción del modelo original fue considerado simplemente como un fondo común con capacidad de procesamiento general que contenía la información que no estaba directamente asignada a alguno de los dos subsistemas de la memoria de trabajo.47 De forma general se asume que el componente ejecutivo es responsable del control atencional de la memoria de trabajo. La forma en la que se ejerce este control fue propuesta inicialmente por el modelo de Norman y Shallice,59 que proponía dos procesos de control: el primero controlaba la conducta por medio de hábitos y esquemas preestablecidos guiados de forma implícita por claves ambientales, en tanto que el segundo comprendía un controlador atencional limitado (el sistema supervisor de la actividad o SSA) que podía intervenir eventualmente cuando el control de rutina fuera insuficiente.59 Este modelo de tres componentes de la memoria de trabajo enfrenta algunas dificultades cuando trata de explicar cómo ocurre la interacción de este sistema con la memoria de largo plazo, además de que no cuenta con un mecanismo mediante el cual los dos subsistemas, el fonológico y el espacial, interaccionen entre sí. Por esta razón se ha propuesto un cuarto componente: «el buffer episódico», que se sugiere como un almacén de capacidad limitada que permite unir toda la información para integrar episodios congruentes.60 Se sugiere que está controlado atencionalmente por el componente ejecutivo y que es accesible a la conciencia. Su carácter de codificación multidimensional permite la integración de distintos sistemas, y su carácter consciente permite un adecuado proceso de recuperación de la información. Este buffer se presenta como un componente separado de los demás, pero también podría conceptualizarse como el almacén del componente ejecutivo.60
]]>ANATOMÍA
La ubicación anatómica de la memoria de trabajo proviene de estudios de lesiones cerebrales y de estudios de neuroimagen. Estos estudios han demostrado que los tres componentes básicos de la memoria de trabajo están localizados en distintas regiones cerebrales. El caso más comprobado es el del circuito fonológico que se ha asociado con la corteza témporo–parietal izquierda.61,62 En estudios de imagen se han corroborado estos datos, localizándose en el área 40 de Brodmann (AB 40) el componente de almacenamiento del circuito, y en el área 44 (área de Broca) el componente de recuperación de la información.63 La memoria de trabajo visuo–espacial ha sido asociada con el hemisferio derecho.64 Otras áreas relacionadas con la memoria de trabajo verbal son la corteza parietal inferior derecha (AB 40), la corteza premotora derecha (AB 6), la corteza frontal inferior derecha (AB 47) y la corteza occipital extra–estriada, la que se supone que está relacionada con la imaginación visual.65 En cuanto al componente ejecutivo, la mayoría de los estudios de lesión y de imagen funcional coinciden en un origen situado en los lóbulos frontales. Según algunos paradigmas ejecutivos, se activan áreas como la corteza prefrontal dorsolateral bilateral (BA 9 y 46), la corteza frontal inferior (BA 6 y 44) e incluso la corteza parietal (BA 7 y 44).66,67
CONSIDERACIONES FINALES
El estudio de los procesos de memoria y sus sistemas componentes continúa en expansión. En la actualidad podemos reunir un panorama general acerca de la composición de los sistemas de memoria y su funcionamiento; sin embargo estamos lejos de tener el rompecabezas completo, por lo que resulta importante profundizar en el futuro sobre diferentes aspectos relacionados con la memoria comenzando por las discrepancias que existen en definir adecuadamente la taxonomía y los diversos términos que se usan en su investigación para llegar a conocer mejor los circuitos neurales, los procesos celulares y moleculares responsables de cada uno de los componentes de la memoria (adquisición, almacenamiento y recuperación de la información). Se ha cumplido ya un siglo de prolífica investigación científica formal en el terreno de la memoria pero no cabe duda que quizá lo más interesante y revelador esté aún por venir, pues la revolución que han provocado los estudios de biología molecular por una lado y los estudios de imagen funcional en pacientes y personas sanas intactas por el otro, han abierto nuevas fronteras en el terreno de la investigación científica en general y de la memoria en particular, cuyos resultados estaremos por conocer en un futuro cercano. Conocer con precisión el verdadero engrama de la memoria en el Sistema Nervioso, así como la forma en que trabaja y se relaciona con otras funciones cerebrales, tanto en la salud como en la enfermedad, tendría profundas y extensas aplicaciones en muy diversos campos como la pedagogía, la medicina, la psicología, la psiquiatría, la filosofía, etc. Sin embargo, como ocurre en todas las investigaciones científicas, a la vez que día con día se desvelan más misterios en el campo de la memoria, seguramente también se revelarán y enfrentarán otros nuevos.
REFERENCIAS
1. Squire LR. Memory systems of the brain: A brief history and current perspective. Neurobiol Learn Mem 2004;82:171–177. [ Links ]
2. Tulving E. Episodic memory and common sense: how far apart? Philosophical Transactions of The Royal Society of London. Series B. Biological Sciences 2001;356:1505–1515. [ Links ]
3. Dere E, Kart–Teke E, Huston JP, De Souza–Silva MA. The case for episodic memory in animals. Neurosci Biobehav Rev 2006;30:1206–1224. [ Links ]
4. Tulving E. How many memory systems are there? Am Psychologist 1985;40:385–398. [ Links ]
5. Tulving E, Schacter DL. Priming and human memory systems. Science 1990;247:301–306. [ Links ]
6. Tulving E. Episodic memory from mind to brain. Ann Rev Psychol 2002;53:1–25. [ Links ]
7. Wheeler MA, Stuss DT, Tulving E. Toward a theory of episodic memory: The frontal lobes and autonoética consciousness. Psychol Bull 1997;121:331–354. [ Links ]
8. Gilboa A. Autobiographical and episodic memory–one and the same? Evidence form prefrontal activation in neuroimaging studies. Neuropsychologia 2004;42:1336–1349. [ Links ]
9. Tulving E, Markowitsch HJ. Episodic and declarative memory: Role of the hippocampus. Hippocampus 1998;8:198–204. [ Links ]
10. Suddendorf T, Corballis MC. Mental time travel and the evolution of the human mind. Gen Soc General Psychol Mono 1997;123:133–167. [ Links ]
11. Dally JM, Emery NJ, Clayton NS. Cache protection strategies by western scrub–jays (Aphelocoma californica): Hiding food in the shade. Proc Biol Sci 2004;271:S387–S390. [ Links ]
12. Eacott MJ, Easton A, Zinkivskay A. Recollection in an episodic–like memory task in the rat. Learn Mem 2005;12:221–223. [ Links ]
13. Fortin NJ, Agster KL, Eichenbaum HB. Critical role of the hippocampus in memory for sequences of events. Nature Neurosci 2002;5:458–462. [ Links ]
14. Hannesson DK, Howland JG, Phillips AG. Interaction between perirhinal and medial prefrontal cortex is required for temporal order but not recognition memory for objects in rats. J Neurosci 2004;24:4596–4604. [ Links ]
15. Griffiths DP, Clayton NS. Testing episodic memory in animals: A new approach. Physiol Behav 2001;73:755–762. [ Links ]
16. Dere E, Huston JP, De Souza–Silva MA. Integrated memory for objects, places and temporal order: Evidence for episodic–like memory in mice. Neurobiol Learn Mem 2005;84:214–221. [ Links ]
17. Shastri L. Episodic memory and cortico–hippocampal interactions. Trends Cog Sci 2002;6:162–168. [ Links ]
18. Eichenbaum H, Dudchenko PA, Wood ER, Shapiro MR, Tanila H. The hippocampus, memory and place cells: Is it spatial memory or a memory space? Neuron 1999;23:209–226. [ Links ]
19. Eichenbaum H, Fortin NJ. Episodic memory and the hippocampus: It's about time. Curr Dir Psychol Sci 2003;12:53–57. [ Links ]
20. Eichenbaum H, Fortin NJ. Bridging the gap between brain and behaviour. Cognitive and neural mechanism of episodic memory. J Exp Anal Behav 2005;84:619–629. [ Links ]
21. Hunsaker MR, Lee B, Kesner RP. Evaluating the temporal context of episodic memory: The role of CA3 and CA1. Behav Brain Res 2008;188:310–315. [ Links ]
22.Fletcher PC, Frith CD, Rugg MD. The functional neuroanatomy of episodic memory. Tren Neurosci 1997;20:213–218. [ Links ]
23. Nyberg L. Mapping episodic memory. Behav Brain Res 1998;90:107–114. [ Links ]
24. Svoboda E, McKinnon MC, Levine B. The functional neuronatomy of autobiographical memory: A meta–analysis. Neuropsychologia 2006;44:2189–2208. [ Links ]
25. Atance CM, O'Neill DK. The emergence of episodic future thinking in humans. Learn Motiv 2005;36:126–144. [ Links ]
26. Cabeza R. Role of lateral posterior parietal regions in episodic memory retrieval: The dual attention hypothesis. Neuropsychologia 2008;46:1813–1827. [ Links ]
27. Olson IR, Berryhill M. Some surprising findings on the involvement of parietal lobe in human memory. Neurobiol Learn Mem 2009;91:155–165. [ Links ]
28. Robertson EM, Pascual–Leone A, Miall RC. Current concepts in procedural consolidation. Nat Rev Neurosci 2004;5:576–582. [ Links ]
29. Halsband U, Lange RK. Motor learning in man: A review of functional and clinical studies. J Physiol–Paris 2006;99:414–424. [ Links ]
30. Song S. Consciousness and the consolidation of motor learning. Behav Brain Res 2009;196:180–186. [ Links ]
31. Libet B, Gleason CA, Wright EW, Pearl DK. Time of conscious intention to act in relation to onset of cerebral activity (readiness–potenntial). The unconscious initiation of freely voluntary act. Brain 1983;106:623–642. [ Links ]
32. Willingham DB, Salidis J, Gabrieli JDE. Direct comparison of neural systems mediating conscious and unconscious skill learning. J Neurophysiol 2002;88:1451–60. [ Links ]
33. Robertson EM, Pascual–Leone A, Miall RC. Current concepts in procedural consolidation. Nat Rev Neurosci 2004;5: 1–7. [ Links ]
34. Stickgold R, Hobson JA, Fosse R, Fosse M. Sleep, learning and dreams: off–line memory reprocessing. Science 2001;294:1052–1057. [ Links ]
35. Steriade M, Timofeev I. Neuronal plasticity in thalamocortical networks during sleep and waking oscillations. Neuron 2003;37:563–576. [ Links ]
36. Eliasen JC, Souza T, Janes JN. Human brain activation accompanying explicitly directed movement sequence learning. Exp Brain Res 2001;141: 269–280. [ Links ]
37. Mars RB, Coles MG, Grol MJ, Holroyd CB, Niewenhui S et al. Neural dynamics of error processing in medial frontal cortex. Neuroimage 2005; 28:1007–1013. [ Links ]
38. Halsband U, Matsuzaka Y, Tanji J. Neuronal activity in the primate supplementary, pre–suplementary and premotor cortex during externally and internally instructed sequential movements. Neurosci Res 1994;20: 149–155. [ Links ]
39. Ferrari PF, Rozzi S, Fogassi L. Mirror neurons responding to observation of actions made with tools monkey ventral premotor cortex. J Cog Neurosci 2005;17:212–226. [ Links ]
40. Van Mier H, Tempel LW, Perlmutter JS, Raichle ME, Petersen SE. Changes in brain activity during motor learning measured with PET: effects of hand of performance and practice. J Neurophysiol 1999;80:2177–2199. [ Links ]
41. Jueptner M, Weiller C. A review of differences between basal ganglia and cerebral control of movement as revealed by functional imaging studies. Brain 1998;121: 1437–1449. [ Links ]
42. Halsband U, Lange RK. Motor learning in man: A review of functional and clinical studies. J Physiol Paris 2006;99:414–424. [ Links ]
43. Shacter DL, Buckner RL. Priming and the brain. Neuron 1998;20:185–195. [ Links ]
44. Shallice T, Warrington EK. Independent functioning of verbal memory stores: a neuropsychological study. Q J Exp Psychol 1970;22:261–273. [ Links ]
45. Hebb DO. The organization of behavior. New York: Wiley; 1949. [ Links ]
46. Baddeley AD, Hitch GJ. Recent advances in learning and motivation. New York: Edit Bower G.A. Academic; 1974; pp. 47–89. [ Links ]
47. Baddeley A. Working memory: Looking back and looking forward. Nature Reviews 2003;4:829–839. [ Links ]
48. Vallar G, Baddeley AD. Fractionation of working memory. Neuropsychological evidence for a phonological short–term store. J Verbal Learn Verbal Behavior 1984;23:151–161. [ Links ]
49. Conrad R. Acoustic confusion in immediate memory. B J Psychol 1964;55:75–84. [ Links ]
50. Conrad R, Hull AJ. Information, acoustic confusion and memory span. Br J Psychol 1964;55:429–432. [ Links ]
51. Baddeley AD. Short term memory of word sequences as a function of acoustic, semantic and formal similarity. Q J Exp Psychol 1966;18:362–365. [ Links ]
52. Baddeley AD. The influence of acoustic and semantic similarity on long term memory for word sequences. Q J Exp Psychol 1966;18:302–309. [ Links ]
53. Baddeley AD, Wilson B. Phonological coding and short–term memory in patients without speech. J Mem Lang 1985;24:490–502. [ Links ]
54. Papagano C, Vallar G. Phonological short–term memory and the learning of novel words, the effect of phonological similarity and item length. Q J Exp Psychol 1992;44A:47–67. [ Links ]
55. Papagano C, Valentine T, Baddeley AD. Phonological short term memory and foreign language vocabulary learning. J Mem Lang 1991;30: 331–347. [ Links ]
56. Service E. Phonology, working memory and foreign language learning. Q J Exp Psychol 1992;45A:21–50. [ Links ]
57. Baddeley AD, Gathercole SE, Papagano C. The phonological loop as a lenguaje learning device. Psychol Rev 1998;105:158–173. [ Links ]
58. Della Sala S, Gray C, Baddeley AD et al. Pattern span: a tool for unwelding visuo–spatial memory. Neuropsychologia 1999;37:1189–1199. [ Links ]
59. Norman DA, Shallice T. En: Consciousness and self regulation. Advances in research and theory (eds.). Davidson RJ, Schwarts GE, Shapiro D. New York: Plenum Press; 1968; pp. 1–18. [ Links ]
60. Baddeley AD. The episodic buffer. A new component of working memory? Trends Cog Sci 2000;4:417–423. [ Links ]
61. Warrington EJ, Logue V, Pratt RTC. The anatomical localisation of selective impairment of auditory verbal short–term memory. Neuropsychologia 1971;9:377–387. [ Links ]
62. Vallar G, DiBetta AM, Silveri MC. The phonological short–term store rehearsal system: patterns of impairment and neural correlates. Neuropshychologia 1997;35:795–812. [ Links ]
63. Paulesu E, Frith CD, Frackowiak RSJ. The neural correlates of the verbal component of working memory. Nature 1993;362:342–345. [ Links ]
64. Hanley JR, Young AW, Pearson NA. Impairment of the visuospatial sketch pad. Q J Exp Pshychol 1991;43A:101–125. [ Links ]
65. Kosslyn SM et al. Visual mental imagery activates topographically organised cortex. PET investigations. J Cogn Neurosci 1993;5:263–287. [ Links ]
66. Braver TS et al. A parametric study of prefrontal cortex involvement in human working memory. Neuroimage 1997;5:49–62. [ Links ]
67. Baddeley AD. The fractionation of working memory. Proc Natl Acad Sci 1996;93:13468–13472. [ Links ]
68. Conway MA. Episodic memories. Neuropsychologia 2009;47( 11):2305–2313. [ Links ] ]]>