Comparación de las estructuras morfológicas en raíz e hipocótilo en frijol*
Comparison of morphological structures in bean root and hypocotyl
Edwin Javier Barrios-Gómez1§, Cándido López-Castañeda2, Josué Kohashi-Shibata2, Jorge Alberto Acosta-Gallegos3, Salvador Miranda-Colín2, Jaime Canul Ku1 y Netzahualcóyotl Mayek-Pérez4
1 Campo Experimental Zacatepec, INIFAP. Carretera Zacatepec-Galeana, km 0.5 Zacatepec, Mor. México. C. P. 62780. Tel. 01 734 3430230. Fax. 01 734 3433820. (canul.jaime@inifap.gob.mx).§Autor para correspondencia: barrios.edwin@inifap.gob.mx.
]]> 2 Genética y Botánica, Colegio de Postgraduados. Carretera México-Texcoco km 36.5 Montecillo, Texcoco, Estado de México. C. P. 56230. Tel. 01 595 9520200. Ext. 1587,1318 y 1551. (clc@colpos.mx), (jkohashi@colpos.mx), (smiranda@colpos.mx).3 Campo Experimental Bajío, INIFAP, carretera Celaya-San Miguel de Allende km 6.5, Col. Roque, Celaya, Guanajuato, México. C. P. 38110. Tel. 01461 6115326. Ext. 164. (acosta.jorge@inifap.gob.mx).
4 Centro de Biotecnología Genómica, Instituto Politécnico Nacional, Boulevard del Maestro, s/n Esquina Elias Piña, Colonia Narciso Mendoza, Reynosa, Tamaulipas, México. C. P. 77810. Tel. 01 899 9243627. Ext. 7712 y 7742. (nmayek@ipn.mx).
*Recibido: agosto de 2011
Aceptado: abril de 2012
Resumen
En el año 2006 se realizó un experimento, con el objetivo de determinar las diferencias en desarrollo de variedades contrastantes de frijol (Phaseolus vulgaris L.) en su raíz y parte aérea de la plántula. Como se sabe los cultivares tolerantes a sequía tienen vasos de xilema de la raíz e hipocótilo de menor diámetro que los cultivares susceptibles a sequía, se utilizaron cinco materiales de frijol contrastantes, de habito indeterminado tipo III, de los cuales se cosecharon plántulas a los 7,11,15,19 y 23 días después de la siembra (dds); además, en la última cosecha de plántulas (23 dds), se tomaron ejemplares de raíz de aproximadamente 1 cm de longitud. Los cortes en la raíz principal se hicieron en tres posiciones; en la región nodal a 1 cm debajo del eje hipocótilo-raíz, en la parte media y a 1 cm del ápice de cada variedad. El cultivar tolerante a sequía (FM Corregidora) tuvo raíces y vasos de xilema de menor diámetro que el cultivar susceptible a sequía (FM RMC) y el criollo de bajo rendimiento (Michoacán 128). FM Corregidora mostró mayor longitud y peso seco total de raíces que FM RMC y Michoacán 128. Las raíces de FM Corregidora crecen a mayor velocidad que los demás cultivares, confiriéndole un mayor desarrollo en etapa de plántula. El diámetro del hipocótilo y la oquedad central e s mayor en FM Corregidora y los cultivares de alto rendimiento (FJ Marcela y FM Bajío) que en el cultivar susceptible a sequía y el criollo. FM Corregidora, FJ Marcela, FM Bajío y FM RMC tienen corteza más gruesa y mayor número de vasos de xilema que Michoacán 128. El material tolerante a sequía (FM Corregidora) mostró raíces y vasos de xilema de menor diámetro que el cultivar susceptible a sequía (FM RMC) y la variedad criolla de bajo rendimiento (Michoacán 128).
Palabras clave: Phaseolusvulgaris L., transporte vascular, morfología de raíz e hipocótilo, desarrollo de raíz.
]]>Abstract
In 2006 an experiment was conducted with the aim to determining the differences in development of contrasting varieties of bean (Phaseolus vulgaris L.) root and shoot of the seedling. As known, drought-tolerant cultivars have xylem vessels of the root and hypocotyl of smaller diameter than the cultivars susceptible to drought, using five contrasting beans, of indeterminate habit type III, from which seedlings were harvested at 7, 11, 15, 19 and 23 days after sowing (DAS); moreover, in the latest seedling harvest (23 DAS), root samples approximately 1 cm in length were taken. The cuts in the main root were made at three positions; in the nodal región, 1 cm below the hypocotyl-root axis, in the middle and, 1 cm from the apex of each variety. The drought-tolerant cultivar (FM Corregidora) had roots and xylem vessels of smaller diameter than the drought-susceptible cultivar (FM CMR) and low performance landrace (Michoacán 128). FM Corregidora showed greater length and total dry weight of the roots than FM CMR and Michoacán 128. FM Corregidora's roots grow faster than the other cultivars', giving a greater development in the seedling stage. The diameter of the hypocotyl and the central hole is greater in FM Corregidora and, the high-yielding cultivars (FJ Marcela and FM Bajío) than in the susceptible to drought and the landrace. FM Corregidora, FJ Marcela, FM Bajío and FM RMC have a thicker crust and a higher number of xylem vessels than Michoacán 128. Drought-tolerant material (FM Corregidora) showed roots and xylem vessels of smaller diameter than the drought-susceptible cultivar's (FM CMR) and low-yielding landrace's (Michoacán 128).
Key words: Phaseolus vulgaris L., root development, root and hypocotyl morphology, vascular transport.
Introducción
El xilema es el componente anatómico del sistema vascular de las plantas superiores, directamente involucrado en el movimiento del agua de la raíz a los diferentes órganos aéreos. La velocidad con que el agua se mueve a través del xilema hasta el área foliar de la planta, sobre todo cuando las plantas están sometidas a condiciones de estrés hídrico, depende varios factores (Kaufmann et al, 2009; Nardinia et al., 2011); p.e., la densidad de raíces (cm de raíz/cm3 de suelo) que determina la capacidad de extracción de agua del suelo adyacente alas raíces (Wilkinson, 2004; Zimmermann et al, 2004), y el diámetro de los vasos de xilema y la resistencia longitudinal o axial al flujo de agua a través de los vasos del xilema que determina la velocidad con la que el agua se mueve através del sistema vascular (Passioura, 1982; Salleo et al., 1996). Otro factor importante en el movimiento del agua entre el suelo y la raíz es lainterfase entre estos dos, p.e., la continuidad en el flujo de agua en el suelo acoplada a la resistencia hidráulica al fluj o de agua en la raíz, que en condiciones de déficithídrico del suelo, domínala velocidad de absorción del agua (Faiz y Weatherley, 1978). Por otro lado, el floema no merece menor importancia ya que es el principal actor en el movimiento de nutrientes y energía a los diferentes órganos de la planta (Dinant y Lemoine, 2010).
La raíz del frijol es tetrarca; el crecimiento primario de los vasos del xilema está compuesto de protoxilema (vasos pequeños e inmaduros en la conducción de agua) y metaxilema (vasos grandes, que conducen el agua), y el sistema radical está constituido por una raíz o eje principal (crece en forma vertical en el perfil del suelo), tiene un número variable de raíces básales, raíces adventicias que se originan del hipocótilo y raíces laterales que se originan en cada una de éstas categorías de raíces (Rubio y Lynch, 2007).
''La resistencia longitudinal al flujo de agua en las raíces principales puede influenciar la velocidad a la que el agua del subsuelo puede ser transportada por las raíces a través de una capa de suelo seco a los órganos aéreos de la planta y que este carácter está relacionado con el diámetro de los vasos del xilema en las raíces seminales en las plantas de trigo'' (Passioura, 1982). Lawlor (1972) sugiere que las características de la planta y su ambiente aéreo dominan la velocidad de absorción del agua, a menos que el suelo esté tan seco que virtualmente no haya más humedad disponible. En lo que respecta al vastago, la detección de variedades de alto vigor inicial puede contribuir a realizar una mejor selección para condiciones adversas de crecimiento, aunque también se menciona que el vigor es una característica determinada por factores genéticos, pero influenciada por el ambiente (Anday Pinter, 1994).
Leskovar y Stoffella, (1995) y Román-Aviles et al. (2004) indican que la detección de diferencias en el crecimiento y arquitectura de las raíces entre genotipos puede ofrecer posibilidades para selección a tolerancia a enfermedades en las raíces, sequía, inundación, mejor absorción de nutrientes, entre otros. Se ha observado que plantas de hábito I y II tienen una raíz principal más larga, en contraste con las variedades de hábito III. Sus raíces presentan un crecimiento mayor de raíces adventicias (Román-Aviles et al., 2004).
]]> Las raíces adventicias del frijol por definición son aquellas que se desarrollan del hipocótilo y pueden ser numerosas y de crecimiento horizontal (Leskovar y Stoffella, 1995). En maíz, Pérez de la Cerda et al. (2007), encuentra una alta relación (r= 0.99) entre características que confieren el vigor a la plántula y los componentes del rendimiento. Sin embargo, también se ha encontrado en maíz inconsistencia en correlaciones entre los caracteres de plántula y planta adulta (Ájala y Fakorede, 1988). La selección por caracteres anatómicos de la raíz, podría utilizarse en el mejoramiento de la resistencia a sequía al economizar humedad y lograr una utilización más eficaz de esta en condiciones de déficithídrico del suelo. El objetivo del presente trabajo fue determinar las diferencias en desarrollo de variedades contrastantes de frijol en cuanto a su raíz y parte aérea en estado de plántula. Además sí cultivares tolerantes asequía tienen vasos de xilema de la raíz de menor diámetro y menor número de haces de xilema en el hipocótilo, que los cultivares susceptibles a sequía.
Materiales y métodos
Se sembró un experimento en macetas de plástico a la intemperie, el 11 de Mayo de 2006, en el áreade invernaderos del Colegio de Posgraduados en Montéenlo, Municipio de Texcoco, Estadode México(19° 21 'N, 98°55'O y 2250 msnm).
Material vegetal
Se incluyeron cinco variedades comerciales de frijol , de habito indeterminado tipo III, caracterizadas por su rendimiento de semilla y respuesta a la sequía edifica en condiciones de campo (Barrios-Gómez et al., 2010) (Cuadro 1).
Detalle experimental
Para la siembra se emplearon tubos de PVC de 10.5 cm de diámetro y 50 cm de altura, los cuales se llenaron con suelo. En cada tubo se sembraron tres semillas del mismo peso individual (260 ± 5 mg) a una profundidad de 3 cm. Las variedades se asignaron en un diseño experimental de bloques completamente al azar concuatro repeticiones.La unidad experimental consistió de un tubo con suelo y tres plántulas. El suelo empleado fue de textura franco-arenoso [Densidad aparente= 1.0 (g cm3), pH= 6.9, materia orgánica= 9.0%], previamente tratado con calor a una temperatura de 70 °C durante 2 h. Se aplicaron riegos frecuentes, manteniendo el contenido de humedad edáfica cercano a capacidad de campo durante el experimento. Las plántulas crecieron bajo temperaturas máximas y mínimas promedio de 29.6 °C y 7.7 °C, fotoperiodo (13.03) y radiación fotosintéticamente activa de 2 032 mol nr2 s1.
Análisis de crecimiento
Se cosecharon plántulas completas a los 7, 11, 15, 19 y 23 días después de la siembra (dds) en las cuatro repeticiones y en cada plántula se determinó la longitud de la raí z principal (LRP, cm), número de raíces (NRA) y longitud de raíces adventicias (LRA, cm), y el área foliar (AF, cm2) que fue determinada con un integrador de área foliar (marca LI-COR, modelo LI-3100). Los diferentes órganos de las plántulas se secaron en una estufa con circulación de aire forzado (marca RIOSSA, modelo HS 82) a una temperatura de 70 °C durante 72 h, para determinar el peso seco total de raíces (PSTR, mg), peso seco de hojas (PSH, mg), peso seco de tallo (PST, mg) y peso seco de los pecíolos (PSP, mg).
]]> Con los datos obtenidos se calculó la longitud total de raíces [LTR= raíz principal + raíces adventicias + (raíces laterales de primer orden, segundo orden, tercer orden, etc.), cm],peso seco de la parte aérea [(PSPA= PSH + PST + PSP), mg] y cociente raíz/parte aérea (CRPA= PSTR/PSPA).Se calculó la tasa relativa de crecimiento (TRC, mgmg1 d1) de las raíces; TRC= loge (W2) - loge (W1)/(t2 - t1), donde W1 y W2, y t2 y t1 representan el peso seco total de raíces (mg) y el tiempo transcurrido entre la primera y la segunda cosecha de plántulas, respectivamente (Hunt, 1978).
Análisis de estructuras anatómicas de raíz e hipocótilo
En la última cosecha de plántulas (23 dds), se cortaron ejemplares de la raíz principal de un cm de longitud, en tres posiciones; 1 cm debajo de la ''región nodal'' (B), en la parte media entre la ''región nodal'' (M) y 1 cm arriba del ápice de la raíz (A) en las tres plantas de las cuatro repeticiones de cada variedad. Inmediatamente las muestras de tejido se colocaron en viales de plástico de 1.5 mm (Marca Axygen, Inc. USA) con una solución de FAA (alcohol 50%, agua 25%, formol 20%, ácido acético 5%). A los cuatro días de tomadas las muestras, se lavaron con agua destilada y se cambiaron a una solución de GAA (Alcohol 50%, agua 25% y glicerina 25%). Las muestras tratadas con GAA se lavaron con agua destilada y se colocaron en una solución de alcohol a una concentración de 30%duranteunahora;despuéslasmuestras se transfirieron a soluciones de alcohol de 40,50,60 y 70% por espacio de una hora en cada solución. Posteriormente, se procedió a hacer los cortes transversales (aprox. ±0.1 mm), y éstos se colocaron en una solución de alcohol de 80,90 y 100% durante una hora, respectivamente, con el propósito de lograr una deshidratación completa de los tejidos; las muestras de tejido se deshidrataron con una secadora de punto crítico (Sandri-780A), se colocaron y orientaron en porta muestras de latón 2 x 2 cm (hechos en el Colegio de Postgraduados), utilizando cinta doble adhesiva de cobre (Marca Scotch Electrical Tape 3M) y se recubrieron con oro durante cuatro minutos en una ionizadora (Ion Sputter JFC-1100, Jeol, Fine Coat). Concluido el proceso las preparaciones se llevaron aun Microscopio Electrónico (Marca Jeol modelo 35-C, JSM-35C) de la Unidad de Microscopia Electrónica del Colegio de Postgraduados, las imágenes se capturaron en una película negativa en blanco y negro Plus-X 125 asa 120 (Kodak Profesional). Estas imágenes se escanearon y se procedió a identificar y medir las estructuras vasculares de la raíz e hipocótilo, mediante el programa de microcomputadora Image Tool for Windows (Wilcox et a.l, 2002) en el Laboratorio de Morfología de Insectos, del Colegio de Postgraduados.
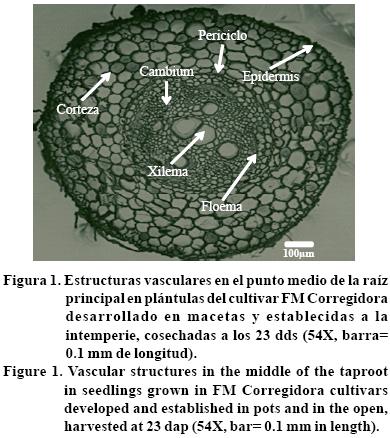
De las secciones anatómicas transversales (tres puntos: B, M y A) de la raíz principal, se contó el número total de vasos del xilema (NTVX), y se midieron el diámetro de raíz (DR, μm), diámetro de los vasos de xilema (DVX, μm), diámetro del cilindro vascular o médula (DCV, μm) y grosor de la corteza (GC, μm); en todos los casos el diámetro se midió en forma horizontal y vertical para calcular un diámetro promedio (Figura 1).
Se muestrearon también secciones anatómicas transversales del hipocótilo a 1 cm arribade la''región nodal'' (H); se contó el número total de haces de xilema (NTHX) y se midió el diámetro del hipocótilo (DH, μm), diámetro de la oquedad central (DOC, μm), diámetro de la médula (DM, μm) y grosor de la corteza (GC, μm); éstos datos se determinaron en dirección horizontal y vertical de laimagen en cadacorte para calcular un promedio, debido a que las estructuras vasculares no son completamente cilindricas y tener una mejor estimación del diámetro de las diferentes estructuras anatómicas (Figura 2).
Se realizó el análisis de varianza para todas las variables medidas con el programa SAS (2008) en forma individual como un bloques completamente al azar (Yij=μ+Ti+Bj+Eij) para cada experimento. Se calculó también la diferencia significativa de Tukey (DSH, p≤ 0.05) para la comparación de medias.
Resultados y discusión
Diversidad en caracteres morfológicos y anatómicos de la raíz
La diversidad en caracteres morfológicos y anatómicos de cortes en diferentes puntos de la raíz principal a los 23 dds, mostraron que todas las variables fueron altamente significativas (p≤ 0.01), excepto el diámetro de la raíz (DR), fue significativa (p≤ 0.05). En el punto M y punto A, la raíz mostró menor diámetro que en el punto B; la raíz tiende a ser más delgada conforme se aproxima al ápice, esto se reflejó en una disminución del diámetro de los vasos de xilema (DVX) y diámetro del cilindro vascular (DCV) y disminución del número total de vasos del xilema (NTVX) (Cuadro 2). La reducción en el DR principal al aproximarse al ápice, se debe a que la división celular activa o mitótica de la raíz no aparece en la región apical, sino a una cierta distancia que varía con las distintas regiones de los tejidos (Esau, 1982).
Por el contrario, el grosor de la corteza (GC) fue menor en el punto B que en los puntos M y A, y aumentó significativamente al aproximarse al ápice de la raíz, al indicar un menor DVX, DCV, y NTVX (Cuadro 2); lo anterior se debe a que en estos puntos de la raí z, el cilindro vascular ocupa menor volumen que la corteza, debido a que la diferenciación longitudinal de los tejidos vasculares primarios es acrópeta (Esau, 1982). Al considerar la variación observada en el DR principal en los tres puntos analizados, se decidió utilizar un valor promedio de los tres puntos, para las comparaciones subsecuentes entre cultivares.
Caracteres morfológicos y anatómicos de raíz en variedades
FM Corregidora, FJ Marcela y FM Bajío tuvieron ejes de la raíz principal con menor DR que el susceptible a sequía (FM RMC) y el criollo (Michoacán 12 8). El menor DR en estos cultivares se reflejó en menor DCV y DVX, y menor GC, excepto FJ Marcela, cuyo DVX fue similar al de FM RMC y al criollo. El NTVX no presentó una tendencia definida entre los cultivares estudiados (Cuadro 3). El DR, DCV y DVX son importantes para el transporte interno del agua en la planta, sobre todo cuando ésta experimenta estrés hídrico. En los cereales un mayor DVX favorece mayor conductancia hidráulica dentro de la raíz y menor resistencia al flujo del agua en el xilema (Passioura, 1982). El DVX se podría utilizar como carácter de selección, al incrementar la resistencia longitudinal al flujo del agua o resistencia axial, en la raíz; sí la humedad edifica disminuye a un nivel que la planta ya no puede absorberla a una velocidad que le permita mantener la tasa transpiratoria durante el llenado del grano, esto resulta en un cierre estomático, y al final del ciclo en semillas pequeñas y un bajo índice de cosecha y rendimiento de semilla. En éstas circunstancias la selección de genotipos con raíces y vasos de xilema de menor diámetro, podría contribuir a conservar agua para la formación de la semilla después de la floración, al reducir el impacto de las deficiencias hídricas. Por el contrario, sí los cultivos dejan una alta proporción de humedad disponible en el suelo en la madurez fisiológica, lo más conveniente será seleccionar cultivares con mayor número y longitud e raíces, de tal forma que favorezcan una mayor absorción de agua y nutrientes (Passioura, 1980; Richards y Passioura, 1981).
El estudio de la variabilidad en las estructuras internas de las raíces del frijol, ofrece un gran potencial para el mejoramiento del rendimiento en condiciones de secano, donde la escasez de humedad y la pre sencia de deficiencias hídricas edáficas en las etapas críticas de desarrollo, afectan severamente el rendimiento de semilla y sus componentes. El presente trabajo muestra claramente que los caracteres más importantes para la adaptación de la planta a la sequía son aquellos que permiten una captura y transporte eficiente del agua y nutrientes al interior de la planta como lo señalan Nardinia et al. 2011 y Van Bela et al. 2011; el diámetro de la raíz y los vasos del xilemade la raíces, sin duda son caracteres importantes para determinar la cantidad y velocidad de movimiento del agua al interior de la planta, sobre todo cuando las raíces detectan deficiencias en la cantidad del agua disponible, para los procesos de crecimiento de la planta (Holste et al, 2006). La longitud y peso seco de las raíces determinan el tamaño del sistema radical y el volumen de suelo que pueden explorar, para la absorción de agua y nutrientes; la velocidad de crecimiento (tasa relativa de crecimiento o tasa de elongación radical) y la abundancia de las ramificaciones de las raíces laterales, son también características importantes para el desarrollo de un sistema radical extendido y profundo, que pueda capturar humedad eficazmente en las capas cercanas a la superficie del suelo y a mayor profundidad en el perfil del suelo.
]]> La asignación de materia seca entre las raíces y la parte aérea de la planta (cociente raíz/parte aérea) constituye otro factor importante en la adaptación a la sequía; los genotipos tolerantes a sequía requieren desarrollar un sistema radical extenso y profundo, constituido por raíces delgadas y con vasos de xilema de menor diámetro, que permita mantener la absorción de agua y nutrientes, para la expansión de nuevos tejidos tanto en el sistema radical como en los órganos aéreos de la planta (área foliar, tallos y estructuras reproductoras). En éste aspecto es importante señalar que el cociente de raíz/parte aérea por sí mismo, no es un criterio de selección efectivo para incrementar la absorción de agua y nutrientes, y la tolerancia a la sequía y rendimiento, habrá que tener cuidado de seleccionar cultivares que además de producir sistemas radicales y extensos también tengan alta acumulación de biomasa en la parte aérea; de otra manera un crecimiento radical excesivo a expensas de los órganos aéreos de la plantapuede resultar en bajo rendimiento.Caracteres morfológicos y anatómicos del hipocótilo en variedades
Las variedades FM Corregidora, FJ Marcela y FM Bajío mostraron mayor diámetro de hipocótilo (DH) que FM RMC y Michoacán 128; ésta diferencia en DH también se observó en el diámetro de la oquedad central (DOCH). FM Corregidora FJ Marcela, FM Bajío y FM RMC tuvieron mayor grosor de la corteza del hipocótilo (GCH) y número total de haces vasculares de xilema (NTHX) que Michoacán 128. No se detectó diferencia significativa entre cultivares para el diámetro del cilindro vascular (DCV) del hipocótilo (Cuadro 3). La anatomía de las estructuras vasculares del hipocótilo difieren de las estructuras traqueales de la raíz; el DH es casi tres veces más grueso que el DR principal; el DCV del hipocótilo es dos veces mayor que el DCV de la raíz principal (Cuadro 6 vs. Cuadro 3). El hipocótilo tiene una oquedad central que podría ocupar el 85 % del diámetro de la raíz principal y una corteza 23% más delgada que la raíz principal (Cuadro 3).


Variación en caracteres de crecimiento de la raíz
Los cultivares FM Corregidora y FJ Marcela produjeron raíces más largas y acumularon mayor peso seco total de raíces (PSTR) que los otros cultivares a los 23 dds. FM Corregidora produjo mayor longitud total de raíces (LTR), de raíz principal (LRP), de raíces adventicias y de raíces laterales (LRAL), mientras que FJ Marcela produjo mayor LTR, LRP y LRAL que los demás cultivares (Cuadro 4). La mayor acumulación de PSTR en FM Corregidora se debió a mayor producción de materia seca en las raíces adventicias y raíces laterales, mientras que el mayor PSTR en FJ Marcela se obtuvo con un mayor PSRP y PSRAL que los otros cultivares (Cuadro 4).
Román-Aviles et al. (2004) estudiando variedades de frijol de diferentes hábitos de crecimiento en invernadero encontraron entre 0 y 8 raíces adventicias por planta, siendo las líneas susceptibles las que tuvieron un menor número de raíces, también encontraron que la línea con mayor número de raíces adventicias, reportada como tolerante a sequía, presentó mayor vigor y mayor profundización de raíces. FM Baj ío y F J Marcela mostraron una raíz principal de mayor longitud, lo cual puede estar relacionado al hábito más compacto, esto se ha observado en plantas de hábito I y II que tienen una raíz pivotante a diferencia de las variedades de hábito III; sin embargo, no encontraron diferencias significativas en cuanto a la longitud de raíces en frijol, donde compararon varias clases comerciales de frijol, encontrando en variedades del tipo I, II y III una longitud total de 1094, 1116 y 1094 cm por planta (Román-Avilé et al.,2004).
]]> En otras especies se ha observado en cuanto a la longitud de las raíces de trigo, cebada y avena (López-Castañeda et al, 1996), maíz (Pérez de la Cerda et al, 2007), en trigo (Liao et al., 2004) y sorgo (Valadez-Gutiérrez et al., 2007) que un mayor desarrollo de la plántula es debido a un mayor número de raíces seminales o adventicias. La opinión tradicional sugiere que un sistema radical vigoroso y extenso combinado con mecanismos de escape a déficit hídricos de la planta es una de las mejores estrategias de la resistencia a sequía (Ludíow y Muchow, 1988). Un alto vigoren el crecimiento de las raíces en la plántula de frijol favorece una alta absorción de agua y nitrógeno (Liao et al, 2004).El cultivar tolerante a sequía (FM Corregidora) produjo sus raíces a mayor velocidad que los demás cultivares; FM Corregidora tuvo mayor tasa relativa de crecimiento (mg de materia seca producida en las raíces por mg de materia seca presente en las raíces por día) de raíces que los otros cultivares (Cuadro 5). Esta ventaja en el rápido crecimiento de las raíces, aunada a sus características de ser raíces más delgadas (menor diámetro) y tener vasos de xilema más estrechos (menor diámetro), pueden ayudar al cultivar FM Corregidora como atributos que le permiten adaptarse mejora la sequía y producir alto rendimiento de semilla (Barrios-Gómez et al, 2010).
Otros estudios realizados en variedades de cebada y trigo harinero han mostrado que un sistema radical vigoroso con abundantes ramificaciones de las raíces laterales, que se distribuyan más uniformemente en el suelo cercano a la superficie, como es el caso de cebada, tienen mayor eficiencia en la absorción y conducción del agua al interior de la planta (López-Castañeda y Richards, 1994); en variedades y líneas recombinantes de frijol también se ha determinado que un sistema radical vigoroso con crecimiento extendido cercano ala superficie del suelo debajo del cultivo favorece mayor absorción de agua y elementos nutritivos como el Fósforo, que tiene baja movilidad en la solución del suelo y baja eficiencia de absorción por la planta(Liao et al., 2004).
Los cultivares FM Corregidora (tolerante a sequía) y FJ Marcela (alto rendimiento) acumularon mayor peso seco de la parte aérea que los demás cultivares, y sólo FJ Marcela (alto rendimiento) produjo mayor área foliar total que todos los demás cultivares en la cosecha final (23 dds) (Cuadro 6). Los cultivares FM RMC (susceptible a sequía), FM Bajío (alto rendimiento) y FM Corregidora (tolerante a sequía) mostraron mayor cociente raíz/parte aérea que los cultivares FJ Marcela (alto rendimiento) y la variedad criolla Michoacán 128 (criollo) (Cuadro 6). Sin embargo, puede observarse que el alto cociente raíz/parte aérea del cultivar FM Corregidora se debió a su mayor peso seco total de raíces que los cultivares FM RMC y FM Bajío, que en realidad deben su alto cociente raíz/parte aérea, al haber acumulado mucho menor peso seco de la parte aérea que FM Corregidora.
Peso seco total de plántula (parte aérea y raíz)
El peso seco total de la plántula (PSTR + PSPA) mostró diferencias (p≤ 0.01) desde los 11 a 23 dds. En el último muestreo, FJ Marcela y FM Corregidora mostraron el mayor (p≤ 0.01) peso seco total de plántula. FM Corregidora siempre fue constante en su desarrollo y obtuvo significativamente los mayores valores, a comparación de FJ Marcela, que en las últimas dos cosechas (19 y 23 dds) logró sobrepasar a todas las demás variedades. FM Bajío fue una variedad que mayor valor (p≤ 0.05) presentó, excepto en el último muestreo. La variedad criolla Michoacán 128 en todos los muéstreos presentó el valor más bajo (p≤ 0.01) para peso seco total de plántula. Se observo, que en estas variedades en el lapso de 15 a 19 dds se comportan como autótrofas, ya que después de haber disminuido su peso seco en promedio de todas las variedades un 33%, a los 19 dds lo incrementaron un 14% del peso inicial. FJ Marcela fue la variedad que perdió más peso (15 dds) y la que se recuperó más rápidamente. Asimismo, fue la que más peso total de plántula registró en los muéstreos subsecuentes (Datos no mostrados). Se ha observado que en cereales (López-Castañeda et al., 1996) esto ocurre como lo encontrado en frijol en el presente estudio alrededor de los 15 dds (150 grados-día).
Conclusiones
Las raíces de FM Corregidora crecen a mayor velocidad y en mayor número que los demás cultivares, lo que le confiere mayor tamaño en la etapa de plántula. Los cultivares FM Corregidora (tolerante a sequía) y FJ Marcela (alto rendimiento) acumularon mayor peso seco de la parte aérea que los demás cultivares, y sólo FJ Marcela produjo mayor área foliar total que todos los demás cultivares en la cosecha final.
Las variedades estudiadas que tuvieron raíces y vasos de xilemade menor diámetro son consideradas como tolerantes a sequía, a diferencia que en el hipocótilo no se observo relación alguna, de mayor número de haces estuviera relacionado con variedades tolerantes a sequía; sin embargo, todas las variedades mejoradas tuvieron mayor número de haces vasculares que la variedad criolla. Esto puede representar parte del trabajo del fito mejoramiento al crear indirectamente el mayor potencial de las variedades mejoradas el poder tener un mayor vigor, que posiblemente sea por la mayor cantidad de agua que pueda distribuir o conducir a los órganos aéreos.
]]>Agradecimiento
Los costos de publicación de este trabajo fueron cubiertos por el Fondo Mixto-Gobierno del estado de Tamaulipas.
Literatura citada
Ájala, S. O. and Fakorede, M. A. B. 1988. Inheritance of seedling-vigor and its association with mature plant traits inmaize populations at two levels of breeding. Maydica 33:121-129. [ Links ]
Anda, A. and Pinter, L. 1994. Sorghum germination and development as influenced by soil temperature and water content. Agron. J. 86:621-624. [ Links ]
Barrios-Gómez, E. J.; López-Castañeda, C; Kohashi-Shibata, J.; Acosta-Gallegos, J. A.; Miranda-Colín, S. y Mayek-Pérez, N. 2010. Rendimiento de semilla y sus componentes en frijol Flor de Mayo en el Centro de México. Agrociencia 44: 481-489. [ Links ]
Dinant, S. and Lemoine, R. 2010. The phloem pathway: Newissues and old debates. Plant Biol. and Pathol. 333: 307-319. [ Links ]
Holste, E. K.; Jerke, M. J. and Matzner, S. L. 2006. Long-term acclimatization of hydraulic properties, xylem conduit size, wall strength and cavitation resistance in Phaseolus vulgaris in response to different environmental effeets. Plant, Cell Environ. ment 29:836-843. [ Links ]
Hunt, R. 1978. Plant growth analysis. The Institute of Biology's Studies in Biology No. 96. Edward Arnold (Publishers) Limited. London, Great Britain. 67 p. [ Links ]
Kaufmann, I.; Schulze-Till, T.; Schneider, H. U.; Zimmermann, U.; Jakob, P and Wegner, L. H. 2009. Functional repair of embolized vessels in maize roots after temporal drought stress, as demonstrated by magnetic resonance imaging. New Phytol. 184:245-256. [ Links ]
Liao, H.; Yan, X.; Rubio, G.; Beebe, S. E.; Blair, M. W. and Lynch, J. P. 2004. Genetic mapping of basal root gravitropism and phosphorous acquisition efficieney in common bean. Functional Plant Biology 31:959-970. [ Links ]
Lawlor, D. W. 1972. Growth and wateruse of Loliumperenne. I. Watertransport. J. of Appl. Ecol. 9:79-98. [ Links ]
Leskovar, D. I. and Stoffella, P. J. 1995. Vegetable seedling root systems: morphology, development, and importance. Hort Science 30: 1153-1159. [ Links ]
López-Castañeda, C. and Richards, R. A. 1994. Variationin temperate cereals in rainfed environments. I. Grain yield, biomass and agronomic characteristics. Field Crops Research 37:51-62. [ Links ]
López-Castañeda, C; Richards, R. A.; Farquhar, G. D. and Williamson, R. E. 1996. Seed and seedling characteristics contributing to variation in early vigor among temperate cereals. Crop Sci. 36: 1257-1266. [ Links ]
Ludlow, M. M. and Muchow, R. C. 1988. Critical evaluation of the possibilities for modifying crops for high production per unit of precipitation. In: Bidinger, F.R. and Johansen, C. (eds.). Drought research priorities for the dryland tropics. ICRISAT, Patancheru, India, p. 179-211. [ Links ]
Nardinia, A.; Lo, G. M. A. and Salleo, S. 2011. Refilling embolized xylem conduits: Is it a matterof phloem unloading? Plant Science 180:604-611. [ Links ]
Passioura, J. B. 1980. The transport of water from soil to shoot in wheat seedlings. J. of Exp. Bot. 31 (120):333-345. [ Links ]
Passioura, J. B. 1982. The role of root system characteristics inthe drought resistance of crop plants. In: Drought resistance in crops with emphasis on rice. IRRI, The Philippines. 71-82 p. [ Links ]
Esau, K. 1982. Anatomía de las plantas con semilla. Primera edición en Español, Editorial Hemisferio Sur, S.A. Buenos Aires, Argentina. 512 p. [ Links ]
Faiz, S. M .A. and Weatherley, P. E. 1978. Further investigations into the location and magnitude of the hydraulic resistances in the soil: plant system. New Phytologist 81:19-28. [ Links ]
Pérez de la Cerda, F. J.; Córdova Téllez, L.; Santacruz Várela, A.; Castillo González, F.; Cárdenas Soriano, E. y Delgado Alvarado, A. 2007. Relación entre vigor inicial, rendimiento y sus componentes en poblaciones de maíz chalqueño. Agric. Téc. en Méx. 33:5-16. [ Links ]
Richards, R. A. and Passioura J. B. 1981. Seminal root morphology and water use of wheat II. Genetic variation. Crop Sci. 21:253-255. [ Links ]
Román-Aviles, B.; Snapp, S. S. and Kelly, J. D. 2004. Assessing root traits associated with root rot resistance in common bean. Field Crops Res. 86:147-156. [ Links ]
Rubio, G. and Lynch, J.P. 2007. Compensation among root classes of Phaseolus vulgaris L. Plant and Soil 290:307-321. [ Links ]
Salleo, S.; Lo, G.M.A.; De Paoli, D. and Zippo, M. 1996. Xylem recovery from cavitation-induced embolism in young plants of Laurusnobilis: a possible mechanism. New Phytologist 132:47-56. [ Links ]
Statistical Analysis System (SAS Institute Inc). 2008. The SAS System Reléase 9.1 for Windows, SAS Institute. [ Links ]
Valadez-Gutiérrez, J.; Mendoza-Onofre, L. E.; Córdova-Téllez, L.; Vaquera-Huerta, H.; Mendoza-Castillo, Ma. del C. y García de los Santos, G. 2007. Tamaños de semilla, substancias vigorizantes y pruebas de vigor en sorgos tolerantes al frío. Agrociencia 41:169-179. [ Links ]
VanBela, A. J. E.; Furcha, A. C. U.; Hafkea, J. B.; Knoblauch, M. and Patrick, J. W. 2011. (Questions)n on phloem biology. 2. Mass flow, molecular hopping, distribution patterns and macromolecular signaling. Plant Science ''Article in Press'' Available online 24 May 2011. [ Links ]
Wilcox, D.; Dove, B.; Me. David, D. and Greer, D. 2002. Image tool for Windows Versión 3.0. The University of Texas, Health Science Center. San Antonio, Texas, U.S.A. [ Links ]
Wilkinson, S. 2004. Water use efficieney and chemical signaling. In: Bacon, M.A. (ed.). Water use efficieney in plant biology. Blackwell Publishing, Great Britain. p. 75-112. [ Links ]
Zimmermann, U.; Schneider, H.; Wegner, L. H. and Haase, A. 2004. Water ascent in tall trees: does evolution of land plants rely on a highly metastable state? New Phytologist 162: 575-615. [ Links ]
]]>