
Interaction of water deprivation and water delivery correlated stimuli in continuous and interval schedules of reinforcement
Emilio Ribes–Iñesta y Edgar Montes1
Universidad de Guadalajara–CEIC
1 Dirigir correspondencia al primer autor
al Centro de Estudios e Investigaciones en Comportamiento, ]]>
Francisco de Quevedo 180, CP 44130 Guadalajara, México.
Correo electrónico: ribes@cencar.udg.mx.
Recibido: 15 de mayo de 2009
Revisado: 17 de julio de 2009
Aceptado: 24 de julio de 2009
Resumen
Se realizó un experimento con ocho ratas para evaluar la interacción de la ausencia/presencia de privación de agua en programas de reforzamiento continuo y de intervalo fijo y variable, con y sin estímulos correlacionados con la entrega de agua. Se emplearon dos grupos de cada rata, cuya única diferencia fue la condición inicial con o sin presentación de estímulos correlacionados. Los resultados mostraron que todas las ratas respondieron en las dos fases iniciales con reforzamiento continuo e intervalo fijo/intervalo variable de agua, a pesar de no estar privadas. Se encontró también que la presencia/ausencia de los estímulos correlacionados con la entrega de agua no tuvo ningún efecto a lo largo del experimento. La frecuencia de respuesta de apretar la palanca varió de acuerdo con la ausencia/presencia de privación, independientemente del programa y frecuencia de entregas de agua, observándose la misma cantidad de respuestas bajo reforzamiento continuo que bajo intervalo fijo/intervalo variable. Se observó que durante la exposición al programa de intervalo fijo/intervalo variable con privación ocurrieron más respuestas de muestreo al bebedero sin agua que respuestas de apretar la palanca. Los resultados se examinan en términos de la funcionalidad de las diversas secuencias molares observadas respecto de la obtención y consumo del agua.
Palabras clave: privación, reforzamiento continuo, intervalo fijo, respuestas de muestreo, ratas.
]]>Abstract
An experiment with eight rats was conducted to examine the interaction among three variables: deprivation level (no deprivation or high deprivation), type of reinforcement schedule (continuous reinforcement, fixed interval or variable interval), and the presence or absence of stimuli correlated with water delivery. Two groups of rats, which differed only in the initial condition to which they were exposed (presence or absence of stimuli), were used. Each and every rat responded in the initial phases with continuous reinforcement and interval schedules, despite not being deprived of water. The presence or absence of the stimuli correlated with water delivery had no effect throughout the experiment. Bar pressing varied as a function of deprivation level, regardless of the reinforcement schedule and of the frequency of water deliveries: comparable response rates were observed under continuous reinforcement and under interval schedules. When exposed to interval schedules while water deprived, there were more probes to the empty water dispenser than bar presses. Results are analyzed in terms of the functionality of the varied molar sequences observed with respect to water procurement and consumption.
Keywords: deprivation, continuous reinforcement, fixed interval, probes, rats.
La teoría del condicionamiento operante ha prestado poca atención conceptual a la privación (y/o eliminación de eventos "aversivos"). La privación de agua, comida u otro objeto o consumible, se ha considerado una operación necesaria para potenciar la acción de determinados reforzadores. Michael (1982) sugirió el término "operaciones de establecimiento" para referirse a las manipulaciones previas a una respuesta operante que hacen que la presentación o la eliminación de un objeto de estímulo sea reforzante (positivo o negativo, respectivamente). Sin embargo, agrupar ambos tipos de operación bajo un mismo término, no modifica sustancialmente su condición marginal respecto de conceptos o categorías con propiedades lógicas dentro de la teoría. Esta actitud contrasta con la importancia teórica que dichas operaciones guardaron en la teoría clásica del aprendizaje en relación a los conceptos de pulsión y de incentivo, y su participación en la determinación cuantitativa y cualitativa del comportamiento (Hull, 1952; Spence, 1960; Mowrer, 1960; Logan, 1960).
Skinner (1938) desarrolló su sistema analizando, al igual que Pavlov (1927), el reflejo alimentario. Desde un inicio, Skinner identificó el "hambre" con la diversidad de conductas relacionadas con comer, e identificó a la pulsión con una operación experimental, la privación de comida, cuya única función era la de permitir la exploración sistemática de los cambios en la conducta de comer en un organismo individual en tiempo real. Así, en sus primeros estudios (Skinner, 1930, 1932) mostró que la frecuencia de presión de palanca en la rata albina (como medida del reflejo alimentario) seguía una función monotónica negativamente acelerada respecto del tiempo de privación, en el que la privación se reducía progresivamente con la ingesta de pelletas en tiempo.
Son pocos los estudios que han evaluado el efecto sistemático de la privación de agua o comida como una variable importante por sí misma. En algunos casos, constituyen observaciones de que la tasa de respuesta en programas intermitentes de reforzamiento sostenidos no se ve afectada por decrementos en la privación de comida en palomas. En otros casos, son estudios que reportan un incremento de la tasa de respuesta en programas múltiples de intervalo variable como efecto del incremento de la privación (Clark, 1958; Hodos, 1959). Otros estudios examinaron la interacción de la privación de alimento con el efecto supresor del choque eléctrico como estímulo punitivo de conducta reforzada con comida en programas de razón fija. Dichos estudios (Azrin, 1960; Azrin, Holz & Hake, 1963) encontraron que la supresión de respuesta, por administración continua de choque eléctrico, en los programas de razón fija con comida era menor mientras mayor era la privación de comida. Destaca el estudio de Carlton (1961) en que se manipuló el retiro de la privación en distintos programas de reforzamiento en operación: Rfc (reforzamiento continuo), IF (intervalo fijo), IV (intervalo variable), RF (razón fija) y RDB (reforzamiento diferencial de tasas bajas). En dicho estudio se encontró que los programas intermitentes de reforzamiento mostraban mayor decremento en la frecuencia de respuesta que el programa de reforzamiento continuo al suspenderse la privación.
Schoenfeld, Antonitis y Bersh (1950) encontraron que el nivel operante incondicional de apretar la palanca aumentó como función de incrementos en la privación de comida. Este dato es importante porque muestra que la privación afecta a una respuesta funcionalmente no establecida en una situación determinada. En este caso, la respuesta de presionar la palanca varió cuantitativamente con la privación de comida, antes del condicionamiento. Otros estudios han reportado niveles operantes incondicionales bajo privación de comida, pero sin manipular cambios en la privación. Destacan las observaciones de Skinner (1938) y las de Ribes y colaboradores (Ribes, Daza, López & Martínez, 1978; Ribes y López, 1979; Polanco, López & Ribes, 1979), quienes sometieron el nivel operante a extinción antes de automoldear a las ratas. Sin embargo, no existen estudios sobre la adquisición de respuestas operantes sin privación, y el efecto posterior de la privación sobre la ejecución.
Se planeó un estudio para evaluar la ejecución en programas de reforzamiento continuo primero, y de intervalo fijo y variable, inicialmente en ausencia de privación de agua y posteriormente con privación de agua. Dado que el efecto de la privación de agua puede depender de la discriminabilidad de su entrega, se manipuló la presencia y ausencia de los estímulos correlacionados explícitamente con su entrega (apagado de la luz general, encendido de la luz del bebedero y sonido del mecanismo del dispensador de líquido).
]]>MÉTODO
Sujetos
Se emplearon ochos ratas macho Wistar albinas de 5 meses de edad, experimentalmente ingenuas. Las ratas tuvieron libre acceso a comida sólida (Nutricubos) en sus cajas habitación. Las sesiones se realizaron durante seis días a la semana, de las 10 a las 14 horas.
Aparatos
Se utilizaron dos cámaras de condicionamiento operante para ratas MED modelo ENV–008, de 24.3 por 29.5 por 29.5 cms, ubicadas en cubículos atenuadores de sonidos. En cada cámara, en el centro de una de las paredes se colocó un bebedero con dispensador de líquido a 2 cms de la rejilla del piso, además de una palanca operativa ubicada a 3.5 cms a la derecha del bebedero. El dispensador de líquido (modelo ENV–252–LIM) registró el consumo de agua en gramos por cada ensayo mediante una báscula. La báscula estaba colocada en una base que sostenía el contenedor de agua. Debido a la operación de este mecanismo, el bebedero presentaba el agua con 3 s de demora respecto de la respuesta, con un intervalo de 3 s de retorno del bebedero a la báscula, en los cuales las respuestas que ocurrían eran registradas pero no tenían ningún efecto. Cada entrega duraba 3 s, y consistía en la disponibilidad inmediata de una gota de agua de 0.01 cm3 en la boquilla del bebedero. Sin embargo, la rata podía obtener un máximo adicional de dos gotas si continuaba manipulando la boquilla del bebedero con la lengua. En el interior del bebedero, se ubicaba un sensor de luz que se interrumpía al introducir la rata la cabeza para beber, y registraba cada ingreso de la cabeza en el bebedero. Un foco blanco de 28 V suministró la iluminación general de la cámara. El foco se ubicaba en la esquina derecha de la pared operativa, a una distancia de 27.5 cm de la rejilla del piso. En cada entrega de líquido se iluminó por 3 s un foco blanco de 28V colocado en el interior del bebedero.
Procedimiento
A lo largo del experimento, las ratas fueron sometidas a condiciones de privación y no privación de agua. Cuando las ratas se sometieron a privación, se les retiró el agua de sus cajas habitación durante 22 horas diariamente, teniendo libre acceso al agua por una hora después de cada sesión. En la condición de no privación, las ratas tuvieron libre acceso al agua en su caja habitación. Todas las sesiones experimentales tuvieron una hora de duración.
Las ratas también fueron expuestas a condiciones en las que se entregaba el agua con y sin los estímulos correlacionados habitualmente (ECA): el ruido del sistema de entrega de agua y la luz blanca en el bebedero. Durante la condición con ausencia de los estímulos correlacionados, al entregarse el agua, la luz general permaneció encendida y la luz del bebedero apagada. Un ruido blanco se presentó durante toda la sesión durante esta condición, y un motor de 127 v permaneció encendido produciendo una vibración sobre la rejilla similar a la que tenía lugar cuando se entregaba normalmente el agua, con el fin de atenuar la estimulación característica producto de los mecanismos del bebedero durante las entregas del agua. En la condición con presencia de los estímulos correlacionados, en cada entrega de agua se apagó la luz general y se encendió durante 3 s la luz blanca en el bebedero, acompañada, sin ningún enmascaramiento, de los estímulos producidos por el mecanismo de entrega de agua.
Las ratas se distribuyeron en dos grupos de cuatro ratas cada uno. Las sesiones de moldeamiento en el Grupo 1 se realizaron con ausencia de ECA, mientras que en el Grupo 2 se realizaron con el procedimiento normal con ECA, en correspondencia con la condición experimental inicial de ausencia/ presencia de ECA para cada grupo. Ambos grupos fueron moldeados a presionar la palanca mediante el procedimiento manual, hasta que cada rata obtuvo 100 entregas de agua en tres sesiones consecutivas. Durante el moldeamiento todas las ratas estuvieron privadas de agua. Las ratas R1 y R6 murieron a los 120 y 200 sesiones de iniciado el experimento, por lo que sólo se presentan los datos correspondientes a dichas sesiones para esas ratas.
]]> La tabla 1 describe las distintas fases del estudio, que constó de cuatro fases experimentales y una de redeterminación. En cada fase, las ratas fueron expuestas a una secuencia A/B/A de 20 sesiones por cada condición, con entregas de agua con ECA o sin ECA. En el Grupo 1, la secuencia inició con la presencia de ECA, mientras que el Grupo 2 inició sin la presencia de ECA. Durante las primeras dos fases, las ratas respondieron a un programa de reforzamiento continuo (Rfc), bajo condiciones de saciedad en la Fase 1 y de privación de agua en la Fase 2. En las siguientes dos fases (Fases 3 y 4), las ratas respondieron a un programa de intervalo fijo (IF) 60 s y a un programa intervalo variable (IV) 60 s, bajo saciedad en la Fase 3, y con privación de agua en la Fase 4. En estas dos últimas fases, se presentó el programa de intervalo fijo de la sesión 1 a la 6 y de la 14 a la 20, con la interpolación del programa de intervalo variable de la sesión 7 a la 13, en cada una de las tres condiciones (con y sin ECA). En la fase de redeterminación, cada grupo se expuso a la condición inicial del experimento.
RESULTADOS
La figura 1 muestra, en escala logarítmica, el número de respuestas de presionar la palanca y de entregas de agua por sesión para cada rata del Grupo 1, que inició el experimento sin privación y con la presencia de ECA. En primer lugar, no se observaron efectos de la ausencia/presencia de los ECA en ninguna de las fases experimentales y la redeterminación. En cambio, sí se observaron efectos de la existencia o carencia de privación de agua, y del programa de reforzamiento operativo. En la Fase 1 con un programa de Rfc y sin privación de agua, todas las ratas respondieron con un rango de 1 a 100 respuestas. Sin embargo, dado que algunas respuestas ocurrieron en el intervalo de entrega del agua, el número de entregas de agua producido fue menor que el de respuestas, con un rango de 1 a 60. En la Fase 2 con un programa de Rfc y con privación de agua, todas las ratas mostraron un incremento en el número de respuestas y de entregas de agua obtenidas. El rango de respuestas osciló de 110 a 800, mientras que el de entregas agua fluctuó entre 100 y 125. En la Fase 3 con un programa de IF 60 s (IV interpolado) y sin privación, el número de respuestas disminuyó, con un rango de 1 a 100 respuestas, y un rango de entregas de agua de 1 a 12. No se observó ningún efecto del IV interpolado entre las sesiones 7 y 13 de la condición con ECA de esta fase. En la Fase 4 con un programa de IF 60s (IV interpolado) y con privación de agua, se produjo un aumento en el número de respuestas, con un rango entre 100 y 1000 respuestas, y un rango de 12 a 60 entregas de agua. Finalmente, en la fase 5, de redeterminación de la primera condición de la Fase 1, se observó un rango de respuestas de 9 a 101, con un rango de 4 a 90 entregas de agua. En las fases 2 y 4, con privación de agua, varió muy poco el número de entregas de agua entre sesiones, con escasas excepciones. En cambio, en las otras tres fases se observó una gran variación en el número de entregas de agua entre sesiones.

La figura 2 muestra, en escala logarítmica, el número de respuestas de presión de palanca y entregas de agua por sesión para cada rata del Grupo 2, que inició el experimento sin privación y sin presencia de ECA. Los resultados son similares a los del Grupo 1, dado que no se observaron efectos sistemáticos de la ausencia o presencia de los ECA. De igual manera, el número de respuestas y entregas de agua fue significativamente mayor durante las fases 2 y 4 con privación de agua, en las que se observó también una distribución más estable de las entregas de agua que en las otras tres fases.
]]>
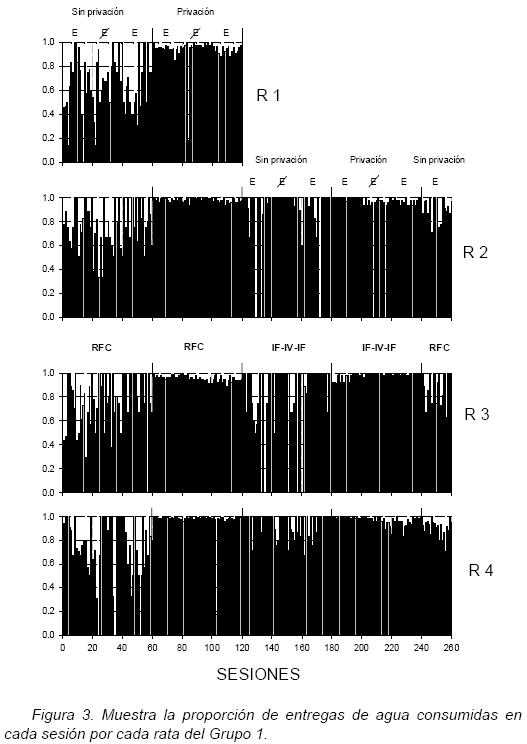
Las figuras 3 y 4 muestran la proporción de entregas de agua consumidas en cada sesión por cada rata del experimento. Puede observarse que durante las dos fases (2 y 4) con privación de agua y la última fase de redeterminación, las ratas consumieron una alta proporción de las entregas de agua, con contadas excepciones. En cambio, en las dos fases (1 y 3) sin privación de agua, se consumió una proporción menor de entregas, especialmente bajo el programa Rfc en el Grupo 1.

La figura 5 muestra el número promedio de entregas de agua, muestreos en el bebedero sin agua y respuestas de presionar la palanca en bloques de 20 sesiones para cada rata de este experimento. Se observó que en las fases sin privación de agua, incluyendo la de redeterminación, el número promedio de ocurrencia de cualquiera de los tres eventos medidos no fue mayor de 20, excepcionalmente, por bloque de sesiones. En cambio, en las dos fases con privación de agua se observaron efectos sistemáticos semejantes en ambos grupos. En la Fase 2 con privación, bajo un programa Rfc, se observó un promedio aproximado de 200 ocurrencias de entregas de agua y de muestreos del bebedero sin agua, y un promedio de entre 300 y 400 respuestas de presionar la palanca en el Grupo 1. En el Grupo 2 fue ligeramente mayor el promedio de muestreos del bebedero que el de entregas de agua, mientras que el de respuestas de presionar la palanca aumentó hasta 600 respuestas en algunas ratas. En la Fase 4 con privación, bajo un programa IF 60 s (IV interpolado), el promedio de entregas agua fue de 20 por bloque de sesiones. Sin embargo, el número de muestreos del bebedero, cuando no aproximado al de respuestas de presionar la palanca, fue superior alcanzando en ocasiones valores de 800 por bloque en ambos grupos.

La figura 6 muestra la mediana y percentiles del número de secuencias de palanquear–palanquear (PP), muestrear–muestrear (MM), palanquear–muestrear (PM), y muestrear–palanquear (MP) de todas las ratas de los grupos 1 y 2 durante la exposición al programa de IF (IV interpolado) bajo privación de agua. Al analizar las secuencias, se pudo incluir una misma respuesta de muestreo o de palanqueo como instancia de dos secuencias, como en el caso de una secuencia mayor PPMP, que incluiría tres secuencias: PP, PM y MP. Estas secuencias se muestran en intervalos sucesivos de 10 s después de cada entrega de agua. Se incluyen sólo los datos de las 6 ratas que concluyeron el experimento. En las secuencias PP se obtuvo una mediana cercana a cero en el primer subintervalo y un incremento progresivo, semejante a una aceleración positiva, en los dos últimos subintervalos con una mediana aproximada de 20 respuestas terminales. En las secuencias PM se obtuvo una mediana terminal de 80 en el último subintervalo, con un incremento progresivo, positivamente acelerado a partir del cuarto subintervalo. En las secuencias MP el nivel inicial en el primer subintervalo mostró una mediana superior a las secuencias previas, con un decremento en el segundo subintervalo y un incremento positivamente acelerado a partir del tercer subintervalo hasta alcanzar una mediana terminal aproximada de 85 ocurrencias. En las secuencias MM, se observó una mediana inicial en el primer subintervalo alrededor de 65 ocurrencias, un decremento en los siguientes dos subintervalos, y una incremento lineal en los tres últimos subintervalos con una mediana terminal de 50 ocurrencias.
]]>
En el apéndice se muestran algunos registros acumulativos de sesiones iniciales y terminales de cada fase de la Rata 4, que son semejantes a los de las demás ratas en este experimento. En estos registros se observó la superposición de las respuestas de palanqueo y de muestreo, o bien su separación. La superposición normalmente se dio en ambos programas de entrega de agua. En los casos en que las respuestas mostraron patrones separados, las respuestas de muestreo pudieron darse por abajo o por arriba de las de palanqueo; sin embargo, en ambos casos, los patrones fueron paralelos en forma en los general, y puede notarse que las respuestas de muestreo coincidieron regularmente con las de palanqueo, especialmente en los programas de IF. La ocurrencia de mayor número de respuestas de muestreo que de palanqueo en los IF no ocurrió siempre en las primeras sesiones, pero sí se observó consistentemente en las últimas sesiones. En el caso del Rfc, las respuestas de palanqueo ocurrieron en "trenes" de mayor longitud al inicio de las sesiones, y de menor longitud con largos periodos de no respuesta al final. Este patrón ocurrió independientemente de que se observaran frecuencias elevadas o bajas de respuesta y, a pesar de los periodos de no respuesta, no se dieron en dichos intervalos respuestas de muestreo. En la primera sesión de la última fase de redeterminación con Rfc sin privación, siempre se observaron patrones de pausa–carrera, semejantes a las ejecuciones bajo programas de razón fija de valores intermedios, pero con un número más elevado de respuestas concurrentes de muestreo.
DISCUSIÓN
Un resultado general observado en todas las ratas de este experimento es que la presencia o ausencia de estímulos correlacionados con la entrega de agua (ECA) no tuvieron ningún efecto en el número de respuestas de presionar la palanca. En un estudio previo (Ribes, Carvajal, Valdés, Torres & Mayoral, en prensa) se observaron mayores frecuencias de respuesta en ausencia que en presencia de los ECA durante la ejecución en programas de Rfc, IF y RF. Sin embargo, en dicho estudio, las ratas fueron expuestas directamente a los programas de IF y RF en la fase inicial de adquisición, y posteriormente a un periodo de sondeo bajo reforzamiento continuo. Su efecto en dicho estudio podría deberse a la interrupción momentánea de la ejecución en curso y, por ello, producir una disminución en la frecuencia de respuesta. Es plausible suponer que, en la medida en que los ECA acompañan consistentemente la entrega del agua, podrían desarrollar funciones discriminativas y probablemente de "reforzamiento condicional" (Skinner, 1938). Sin embargo, en el presente experimento no sólo no se observó ningún efecto de su presencia/ ausencia, sino que el incremento intraciclo de las respuestas de muestreo al bebedero sin agua en su ausencia, sugieren que los ECA no ejercieron ninguna función discriminativa respecto del momento de entrega del agua. El mismo argumento podría aplicarse a las supuestas propiedades inhibitorias o discriminativas de no reforzamiento atribuidas a la ocurrencia del reforzador en los programas de intervalo fijo (Staddon, 1970). En el presente estudio se observó un incremento, y no un decremento, de las respuestas de muestreo al bebedero en los primeros 10 s posteriores a la entrega y consumo del agua, lo que contradice dicha hipótesis, más aún cuando el efecto se produjo tanto en condiciones de privación como en su ausencia.
Se observó que todas las ratas de este experimento respondieron en la palanca desde la primera sesión del programa inicial de Rfc con agua, a pesar de que no se encontraban privadas de líquido. Este es quizá el primer reporte en la literatura de adquisición de una ejecución operante sin privación, aunque durante las tres sesiones de moldeamiento las ratas fueron privadas. Las ratas obtuvieron cuando menos una entrega de agua por sesión y hubo sesiones con 30 ó 50 entregas, una proporción de las cuales, nunca mayor a 40%, en ocasiones no fue consumida durante la primera fase de adquisición. En la última fase de redeterminación, también sin privación, las ratas mostraron un número comparativamente más alto de respuestas que en la primera fase y consumieron prácticamente toda el agua obtenida. Al exponer a las ratas al Rfc con privación de agua, aumentó el número de respuestas con un rango de 200 a 1000 respuestas por sesión. La exposición posterior de las ratas a un programa IF (interpolado IV) sin privación y con privación produjo efectos semejantes, sin observarse cambios durante la operación del programa de IV. Estos resultados parecen corregir cuando menos los observados por Carlton (1961), quien encontró que el retiro de la privación produce mayor decremento en las ejecuciones con programas de intervalo, razón y diferencial de tasas bajas que en los de reforzamiento continuo. Carlton manipuló la privación, eliminándola, una vez establecida la ejecución en dichos programas, mientras que en este experimento la ejecución en los programas de Rfc e IF (IV interpolado) se adquirió en ausencia de privación.
Los resultados de este experimento sugieren que el número de respuestas está determinado por la privación–no privación, más que por la frecuencia o número de entregas de agua obtenidas. Durante las dos fases de no privación, el número de entregas de agua y de respuestas emitidas fue semejante bajo los programas de Crf y de IF (IV interpolado). Resulta sorprendente, sin embargo, que durante la exposición a un Rfc o a un IF (IV interpolado) bajo condiciones de privación, el número (o frecuencia) de respuestas fuera semejante a pesar de que el número de entregas de agua fue mayor durante el programa de Rfc. Este hallazgo no coincide con el observado por Clark (1958), que encontró que el incremento de la tasa de respuesta por la privación interactuaba multiplicativamente con la frecuencia del reforzamiento del programa. También debe destacarse que, a pesar de que bajo el Rfc cada entrega de respuesta interrumpía la respuesta de presionar la palanca, todas las ratas emitieron mayor número de respuestas que el número de entregas de agua recibidas. Este efecto, como ya se mencionó, puede deberse a que las ratas continuaron respondiendo en la palanca durante el periodo de entrega del agua y de acción del mecanismo atenuador del sonido de su entrega. Pero dado que este efecto se produjo tanto en presencia como en ausencia de los ECA, la interpretación más plausible es que las ratas siguieron respondiendo durante el periodo de entrega en el que no obtenían agua adicional. De este modo, puede decirse que las ratas se impusieron un programa de intermitencia en la entrega del agua, equivalente a una razón fija o variable 2 ó 3, a pesar de que solo se requería una respuesta por entrega de agua. Estos resultados son semejantes a los encontrados por Notterman y Mintz (1965), quienes encontraron que, al reforzar diferencialmente de manera continua el esfuerzo realizado al presionar la palanca, las ratas se imponían una contingencia intermitente al emitir respuestas subumbral del requisito.
El número de respuestas de presionar la palanca y de muestreos en el bebedero fue semejante bajo ambos programas, en ausencia de privación, no así bajo condiciones de privación. Bajo no privación no se observaron diferencias en la ejecución en Rfc o IF (IV interpolado). Con privación, en el Rfc el número de presiones de palanca fue mayor que el de muestreos del bebedero, lo que indica que durante la entrega del agua las ratas siguieron presionando la palanca, y los ingresos al bebedero se limitaron probablemente a muestreos inmediatamente después del consumo de agua. En cambio, durante el IF (IV interpolado) se incrementaron los muestreos del bebedero. Este incremento fue bitónico, como se muestra por el número elevado de secuencias MP y MM en todas las ratas en el primer subintervalo posterior a la entrega de agua. Primero hubo un incremento de muestreos, aquí no interrumpido por una nueva entrega en caso de una presión de la palanca (como ocurriría en el Rfc). Segundo, hubo una caída de los muestreos en los siguientes dos ciclos de 10 s produciéndose una aceleración positiva que concluyó en el sexto ciclo y la entrega de agua nuevamente, después de presionar la palanca. En cambio, las respuestas de presionar la palanca se incrementaron a partir del cuarto o quinto subintervalo con un patrón positivamente acelerado, en menor número las respuestas de muestreo. Esto sugiere que la privación potenció en mayor grado la respuesta de muestreo del bebedero que la de presionar la palanca. El patrón de respuestas de muestreo del bebedero podría considerarse semejante a los de otras conductas "adjuntivas" (Falk, 1967). Si tal efecto, como se observó en este experimento, ocurre solamente bajo condiciones de algún tipo de privación, no sería un fenómeno vinculado al programa, como se ha conceptuado tradicionalmente. Por el contrario, se trataría de un fenómeno estrictamente motivacional, que dependería más bien de que las respuestas "adjuntivas" interfieran o sean compatibles con la obtención del objeto motivacional pertinente en la situación.
Recientemente, Bruner y Ávila (2002) sugirieron que las conductas adjuntivas ocurren porque son reforzadas (concurrente y/o adventiciamente), al igual que las respuestas operantes o "instrumentales" prescritas por el programa. Sin embargo, nosotros agregaríamos que para que esto ocurra se requiere de tres factores. Uno, que el programa de reforzamiento explícito prescriba intervalos no funcionales para cualquier tipo de ejecución. Otro, que la naturaleza de la respuesta adjuntiva no sea funcionalmente incompatible con la respuesta operante prescrita por el programa. Finalmente, que los "reforzadores" vinculados a la respuesta prescrita y a la adjuntiva no sean antagónicos. Esto explicaría que en este experimento los muestreos al bebedero hayan ocurrido en mayor número durante el IF (IV interpolado) que en el Rfc, en cuya presencia los muestreos ocurrieron en igual número o por abajo que las respuestas de presionar la palanca. En el Rfc, las respuestas de muestreo habrían interferido con la obtención de agua, tal como lo reportó Falk (1996). La polidipsia desaparece con la exposición a un programa de Rfc, mientras que probablemente ocurriría lo contrario en un programa de TF de valores cortos (equivalentes al intervalo entre reforzadores de un Rfc), ya fuera de comida o, inclusive, de agua.
]]> Innis, Simmelhag y Staddon (1983) encontraron que al exponer a palomas a programas de tiempo fijo de intervalos largos (60 a 300 s), no se observó conducta interina sistemática dirigida al comedero, como suele ocurrir bajo intervalos cortos (5 s). Para dar cuenta de ello, propusieron, siguiendo a Craig (1918), una explicación basada en un solo estado motivacional, que distingue las conductas del animal en dos categorías: apetitivas y consumatorias. Ambas conductas son controladas por el mismo estado motivacional, pero ocurren en momentos diferentes: la conducta consumatoria en presencia del objeto privado y la conducta apetitiva en su ausencia. Sin embargo, esta propuesta poco agrega al sentido común: se consume en presencia del consumible, y se muestran conductas de "espera", "búsqueda" o "preparación" en su ausencia.Se puede realizar un análisis alternativo en términos de las propiedades funcionales de las distintas formas de respuesta en el contexto de un patrón molar de comportamiento. En principio, pueden identificarse distintos tipos funcionales de respuesta en un patrón, dada una contingencia motivacionalmente estructurada: 1) la respuesta contingente (RC) a la presencia del objeto de estímulo motivacionalmente dominante (EIC, reforzador, ocurrencia de la respuesta prepotente ante cierta situación, etc.); 2) las respuestas requeridas (RR) para que la respuesta contingente pueda ocurrir (presionar la palanca, estar próximo al comedero, etc.); las respuestas compatibles (RCO) con las RR y RC (como son los desplazamientos circundantes al comedero, conductas de acicalamiento próximas al objeto motivante, respuestas de orientación que facilitan las respuestas de aproximación, etc.); 4) las respuestas incompatibles (RIN) con las RR y RC (desplazarse en sentido contrario al comedero, orientarse en oposición a las señales pertinentes, ingresar al bebedero cuando antes se requiere apretar la palanca, etc.); y, Analmente, las respuestas irrelevantes (RI), cuya ocurrencia no afecta a las RC y RI.
En este experimento, el programa en operación determinó las propiedades funcionales de las respuestas de muestreo del bebedero. En ambos casos, la respuesta contingente fue consumir el agua, mientras que la respuesta requerida fue la secuencia de presionar la palanca e ingresar al bebedero. Sin embargo, en el Rfc, ingresar al bebedero sin presionar la palanca fue una respuesta incompatible, pues impedía que cada respuesta en la palanca produjera una gota de agua. En cambio, en el IF (interpolado IV), las respuestas de muestreo del bebedero no eran incompatibles siguieran o no a una presión de la palanca hasta el momento de la disponibilidad de la entrega a partir de los 60 s posteriores a la entrega previa. Eso explicaría: a) el incremento inicial en el primer ciclo de 10 s de la conducta de muestreo en la ausencia de respuestas ante la palanca, b) después de una caída en el segundo y/o tercer subintervalos, el incremento positivamente acelerado de los tres tipos de secuencias con componente de muestreo, y c) el mayor aumento de la ocurrencia de las dos secuencias con componente palanca–muestreo, MP y PM, en el último subintervalo vinculado a la entrega del agua. El muestreo inducido parece ocurrir sólo cuando no es incompatible con la respuesta requerida y la respuesta contingente al objeto de estímulo motivante. En este experimento, el mayor número de secuencias MM ocurrieron, precisamente, en el primer subintervalo inmediato posterior a la entrega de agua. De igual manera, en ausencia de privación, al no existir un solo objeto motivacionalmente dominante, los límites entre respuestas requeridas, compatibles, incompatibles e irrelevantes se vuelven borrosos. Al diluirse los límites funcionales entre tipos de respuestas, se incrementa la variación (como carencia de dirección) en la conducta, tal como lo reportó Carlton (1962), al estudiar el efecto del retiro de la privación en la ejecución bajo programas de razón y de intervalo.
En resumen, en este experimento se encontraron resultados que, cuando menos, pueden calificarse como atípicos y de difícil interpretación por la teoría del condicionamiento operante: la adquisición de respuestas operantes bajo condiciones de saciedad, la carencia de propiedades discriminativas de "no reforzamiento" de los estímulos correlacionados con la entrega del agua y de la propia entrega del agua, la determinación del número de respuestas por la condición privación/no privación independientemente del programa y frecuencia de reforzamiento, la ocurrencia de muestreos del bebedero "adjuntivos" bajo programas de IF, la posibilidad de que las conductas adjuntivas no sean inducidas sino solamente toleradas o permitidas por el programa, y la obtención de frecuencias elevadas de respuesta bajo un programa de Rfc. La fidelidad a una tradición empirista recomienda, cuando menos, la necesidad de reflexionar sobre estos resultados y los conceptos y "principios" fundamentales.
REFERENCIAS
Azrin, N. H. (1960). Effects of punishment intensity during variable–interval reinforcement. Journal of the Experimental Analysis of Behavior, 3, 123–142. [ Links ]
Azrin, N. H., Holz, W. C. & Hake, D. F. (1963). Fixed–ratio Punishment. Journal of the Experimental Analysis of Behavior, 6, 141–148. [ Links ]
]]>Bruner, C. A. & Ávila, R. (2002). Adquisición y mantenimiento del palanqueo sin privación explícita del reforzador. Revista Mexicana de Análisis de la conducta, 28, 107–130. [ Links ]
Carlton, P. L. (1961). The interacting effects of deprivation and reinforcement schedule. Journal of the Experimental Analysis of Behavior, 4, 379–381. [ Links ]
Carlton, P. L. (1962). Effects on deprivation and reinforcement–magnitude of response variability. Journal of the Experimental Analysis of Behavior, 5, 481–486. [ Links ]
Clark, F. (1958). The effect of deprivation and frequency of reinforcement in variable–interval responding. Journal of the Experimental Analysis of Behavior, 1, 221–228. [ Links ]
Craig, W. (1918). Appetites and aversions as constituents of instincts. Biological Bulletin, 34, 91–107. [ Links ]
]]>Falk, J. L. (1966). The motivational properties of schedule–induced polydipsia. Journal of the Experimental Analysis of Behavior, 9, 19–25. [ Links ]
Falk, J. L. (1967). Control of schedule–induced polydipsia: Type, size, and spacing of meals. Journal of the Experimental Analysis of Behavior, 10, 199–206. [ Links ]
Ferster, C. B. & Skinner, B. F. (1957). Schedules of reinforcement. Englewood Cliffs, NJ: Prentice–Hall. [ Links ]
Hodos, W. (1959). The effects of several conditions of motivation on two types of reinforcement. Journal of the Experimental Analysis of Behavior, 2, 256. [ Links ]
Hull, C. L. (1952). A behavior system. New Haven: Yale University Press. [ Links ]
]]>Innis, N. K., Simmelhag–Grant, V. L. & Staddon, J. E. R. ( ) Behavior Induced by periodic food delivery. Journal of the Experimental Analysis of Behavior, 39, 309–322. [ Links ]
Logan, F. A. (1960). Incentive. New Haven. Yale University Press. [ Links ]
Michael, J. (1982). Distinguishing between discriminative and motivational functions of stimuli. Journal of the Experimental Analysis of Behavior, 37, 149–155. [ Links ]
Mowrer, O. H. (1960). Learning theory and behavior. Nueva York: Wiley. [ Links ]
Notterman, J. P. & Mintz, D. E. (1965). Dynamics of response. Nueva York: Wiley. [ Links ]
]]>Pavlov, I. P. (1927). Conditioned reflexes. Oxford: Oxford University Press. [ Links ]
Polanco, R., López, F. & Ribes, E. (1979). Efectos de la probabilidad de alimento contingente y no contingente en un programa temporal señalado. Revista Mexicana de Análisis de la conducta, 5, 171–184. [ Links ]
Ribes, E., Daza, B.C., López, F. & Martínez, P. (1978). Efectos del reforzamiento no contingente y demorado en dos respuestas concurrentes. Revista Mexicana de Análisis de la conducta, 4, 141–153. [ Links ]
Ribes, E. & López, F. (1979). La adquisición de operantes concurrentes bajo un programa señalado de reforzamiento definido temporalmente. Revista Mexicana de Análisis de la conducta, 5, 41–55. [ Links ]
Ribes, E., Carvajal, Y., Valdez, U., Torres, C. & Mayoral, A. (en prensa). Acquisition of bar–pressing under interval and ratio–schedules in the presence and absence of stimuli correlated with water delivery. European Journal of Behavior Analysis . [ Links ]
]]>Schoenfeld, W. N., Antonitis, J. H. & Bersh, P. J. (1950). Unconditioned response rate of the white rat in a bar pressing apparatus. Journal of comparative and Physiological Psychology, 43, 41–48. [ Links ]
Skinner, B. F. (1930). On the conditions of elicitation of certain eating reflexes. Proceedings of the National Academy of science, Washington, 16, 433–438. [ Links ]
Skinner, B. F. (1932). Drive and reflex strength. Journal of General Psychology, 6, 22–37. [ Links ]
Skinner, B. F. (1938). The behavior of organisms. Nueva York: Appleton Century Crofts. [ Links ]
Spence, K. W. (1960). Behavior theory and learning. Englewood Cliffs, NJ: Prentice–Hall. [ Links ]
]]>Staddon, J. E. R. (1970). Effect of reinforcement duration on fixed–interval responding. Journal of the Experimental Analysis of Behavior, 13, 9–11. [ Links ]
]]>