
Potencial ostrícola del brazo oeste de Bahía San Quintín: Biomasa actual y estimación preliminar de la capacidad de carga
Oyster culture potential in the west arm of San Quintín Bay: Current biomass and preliminary estimate of the carrying capacity
Zaul García-Esquivel1*, Marco Aurelio González-Gómez1, Francisco Ley-Lou2 y Adán Mejía-Trejo2
1 Instituto de Investigaciones Oceanológicas, Universidad Autónoma de Baja California. Km. 103 Carretera Tijuana-Ensenada Ensenada 22860, Baja California, México. *E-mail: sgarcia@uabc.mx
]]> 2 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California. Km. 103 Carretera Tijuana-Ensenada Ensenada 22860, Baja California, México.
Recibido en julio de 2002;
Aceptado en julio de 2003.
Resumen
De 1995 a 1998 se cuantificó la biomasa del ostión del Pacífico, Crassostrea gigas, cultivado en dos sitios del brazo oeste de Bahía San Quintín (BSQ). Adicionalmente se evaluaron in situ (1999-2000) las tasas de filtración e ingestión con el fin de estimar la capacidad de carga ostrícola del brazo oeste. Al final de nueve meses la producción por sarta varió entre 0.8 y 2.8 kg ostión, registrándose los valores más bajos en un año de intensa mortalidad (1998) y en las sartas con menor densidad de siembra inicial. Se estimó un máximo de 483 kg ostión estante-1 año-1, pero este valor debe reducirse hasta un 30% si únicamente se consideran ostiones de talla comercial (≥ 90 mm). El porcentaje de intercambio del agua entre el brazo oeste y el mar adyacente (6% al 76% del volumen total, referido al NMM) y la concentración media de seston total (2.5 a 14.3 mg L-1) estuvieron fuertemente afectados por las mareas. No obstante, la tasa de ingestión neta de ostiones adultos (2.6 a 3.2 mg mat. org ostión-1 h-1) fue regulada y tendió a mantenerse independiente de la concentración de seston. Se sugiere que estableciendo un límite del 25% de reducción del alimento disponible para el ostión cultivado, la capacidad de carga ostrícola en el brazo oeste de BSQ oscila entre 1913 y 5974 t. Se sugiere que la reducción en las tasas de crecimiento de C. gigas observadas en 2000 podrían deberse a cambios en la cantidad/calidad del alimento disponible en BSQ y/o a competencia por alimento con otros filtroalimentadores.
Palabras clave: Bahía San Quintín, capacidad de carga, ostión, Crassostrea gigas, ingestión.
Abstract
]]> Biomass production of cultured Pacific oysters, Crassostrea gigas, from the west arm of San Quintín Bay (SQB) was quantified from 1995 to 1998. Filtration and ingestion rates were also mesured in situ (1999-2000) to estimate the oyster carrying capacity of the west arm. At the end of nine months oyster production varied between 0.8 and 2.8 kg oyster per string. Lower values were recorded for strings with a low initial density and during a year of high oyster mortality (1998). A maximum of 483 kg oyster rack-1 yr-1 was estimated, but this value should decrease by 30% if only commercial-sized oysters (≥ 90 mm) are considered. Percent water exchange (6% to 76% of total volume, referred to MSL) and seston levels (2.5 to 14.3 mg L-1) were highly dependent on tides. However, adult oysters regulated their net ingestion rates (2.6 to 3.2 mg organic matter oyster-1 h-1), regardless of seston concentrations. It is suggested that establishing a 25% depletion limit for the available food, the carrying capacity of the west arm of SQB ranges from 1913 to 5974 t, and that the observed reduction of growth rates in cultured oysters during 2000 may have resulted from changes in the quantity/quality of the food available in SQB and/or competition for food with other filter-feeders.Key words: San Quintín Bay, carrying capacity, oyster, Crassostrea gigas, ingestion rate.
Introducción
Bahía San Quintín (BSQ) es uno de los cuerpos costeros más productivos y estudiados de la costa noroccidental de México. Desde 1979 se desarrollaron exitosamente en BSQ cutivos piloto del ostión japonés Crassostrea gigas y, de acuerdo con cifras oficiales, su producción comercial aumentó desde 100 hasta 900 t en la década de los ochenta, cifra que no ha sido rebasada hasta ahora (Depto. Acuacultura, SAGARPA, Ensenada, BC).
Desde 1987 toda la producción de C. gigas en BSQ se ha basado en la fijación remota de larva pedivéliger, obtenida principalmente de laboratorios estadounidenses (Héctor González, com. pers., Empresa Integradora Acuacultores de San Quintín). La metodología y estructuras de cultivo utilizadas fueron descritas detalladamente por Polanco et al. (1988). Brevemente, la fijación de larva pedivéliger se realiza dentro de tanques de concreto provistos con aireación, utilizando como sustrato conchas de ostión contenidas en costales de malla. Una vez fijada la larva, los costales se trasladan a los canales del brazo oeste de Bahía San Quintín (Bahía Falsa, BF), donde permanecen por un período de, aproximadamente, dos meses. Posteriormente se fabrican sartas de 1.2 m con siete conchas madre, las cuales son suspendidas en estructuras rectangulares (estantes) hechas de plástico PVC y localizadas en la zona intermareal, en donde permanecen hasta el momento de la cosecha (Polanco et al., 1988). A pesar de que el ostricultivo es una actividad comercial desde 1980, hasta ahora no está documentada la biomasa que, en promedio, se produce por estante. Tampoco se conoce la relación entre las distintas variables morfométricas del ostión cultivado en la laguna, aun cuando existe la posibilidad de realizar estimaciones rápidas de biomasa a partir de variables fácilmente medibles tales como la longitud de la concha.
Actualmente 21 empresas ostrícolas mantienen aproximadamente 9000 estantes de cultivo distribuídos a lo largo de las 730 ha concesionadas o permisionadas para la ostricultura, en el brazo oeste (Héctor González, com. pers., Acuacultores de San Quintín). Por lo anterior, es necesario estimar el impacto potencial que ejerce el ostión cultivado sobre el alimento disponible en esa porción de Bahía San Quintín. Dicha estimación no sólo requiere de información sobre la biomasa cultivada, sino también del volumen de renovación del agua y las tasas fisiológicas de los organismos (Carver y Mallet, 1990). Varios autores han realizado estimaciones del impacto de los filtroalimentadores sobre el alimento disponible en lagunas costeras, tomando como base la concentración de partículas orgánicas presentes (seston orgánico, clorofila a) y las tasas de filtración/ ingestión de los organismos (Cabanas et al., 1979; Hily et al., 1991; Carver y Mallet, 1990; Uribe y Blanco, 2002). También se ha incorporado directamente la velocidad de las corrientes en las estimaciones de la cantidad de alimento disponible (Cabanas et al., 1979), o indirectamente, incluyendo en las ecuaciones el prisma de mareas (Carver y Mallet, 1990) o el tiempo de residencia del agua (Uribe y Blanco, 2002).
El presente estudio tuvo como objetivo general estimar el potencial de carga ostrícola en Bahía Falsa, a partir de las tasas de ingestión del ostión cultivado y del alimento disponible. Para lo anterior, (a) se cuantificó in situ la mortalidad y la biomasa de ostión producida por sarta y por estante en la parte interna y externa del brazo BF; (b) se identificaron las relaciones morfométricas de los ostiones cultivados en ambos sitios de BF; (c) se estimó el alimento disponible y las tasas de ingestión in situ de ostiones adultos, a partir del método de bio-depósitos; y (d) se estimó el potencial ostrícola del cuerpo de agua a partir de a, b y c, y del cálculo del prisma de mareas.
Materiales y métodos
]]> Sitio de estudioBahía San Quintín tiene una superficie aproximada de 42 km2 referida al nivel medio del mar (Chávez-de-Nishkawa y Alvarez-Borrego, 1974). La comunicación al mar ocurre a través de una sola boca (fig. 1), y sólo los canales principales que corren a lo largo de ambos brazos presentan profundidades máximas entre 5 y 15 m, mientras que el resto de BSQ tiene una profundidad media de 2 m (Martori-Oxamendi, 1989). El factor más importante de la hidrodinámica es la marea astronómica de tipo semidiurno, la cual genera corrientes en la boca de hasta 1 m s-1 (Ocampo-Torres, 1980). El transporte de propiedades fisicoquímicas entre los brazos oeste (BF) y este (BSQ) es menor del 10%. El cultivo de ostión se limita BF (fig. 1), donde los estantes de cultivo quedan expuestos al aire un promedio de 15% del tiempo, durante las mareas bajas (García-Esquivel et al., 2000). Tradicionalmente, los estantes (6 x 1.2 m, largo x ancho) se ordenan formando líneas contínuas, con una separación de 2 m entre estantes de la misma línea y con aproximadamente 40 m entre cada línea. Por lo tanto, la superficie efectiva ocupada por un estante es de aproximadamente 160 m2 (6 x 20 m). Los tiempos de residencia estimados para BF a partir de balances de sal varían entre 4 y 6 días en los meses de primavera y verano, o alrededor de 12 días en invierno (Juárez-Villarreal, 1982; Hernández-Ayón et al. , este volumen). El brazo BF funciona típicamente como un sistema exportador neto de fósoro inorgánico e importador de nitrógeno inorgánico y carbono orgánico con respecto al oceáno; no obstante, en años El Niño mantiene una producción neta de nitrógeno inorgánico (Hernández-Ayón et al., este volumen). Las surgencias parecen ser un fenómeno intermitentemente en la costa adyacente a BSQ y la principal causa de variabilidad temporal de biomasa fitoplanctónica (Lara-Lara et al. , 1980). Por lo anterior, las variaciones de clorofila y temperatura en BF siguen el mismo patrón que las mareas, siendo esto más evidente en la zona cercana a la boca (Hernández-Ayón et al., este volumen).
Mortalidad, rendimiento y relaciones morfométricas
El diseño experimental y la metodología para evaluar el crecimiento y la mortalidad del ostión cultivado en BSQ durante 1995-1996 fueron descritos por García-Esquivel et al. (2000). Brevemente, 5 sartas dedicadas para mortalidad (1 sarta/estante) y 60 sartas para crecimiento (12 sartas/estante) fueron marcadas y distribuídas tanto en la cabecera (sitio 1) como en la boca de BF (sitio 6, fig. 1). En cada sitio se usaron dos cohortes de ostión (larvas fijadas en abril y julio de 1995). Mensualmente y durante nueve meses se evaluó la mortalidad acumulativa in situ, en las sartas dedicadas para mortalidad mediante el conteo total de organismos vivos y muertos. Adicionalmente, cada mes se recolectaron cinco sartas por estante (sartas de crecimiento) con el fin de cuantificar la bio-masa y determinar las relaciones morfométricas del ostión. En el laboratorio se desprendieron los organismos de cada sarta, y se seleccionaron al azar 15 organismos/sarta para medir la longitud máxima de la concha y el peso entero vivo. Los ostiones se desconcharon y se estimó el peso seco de sus tejidos secándolos a 100°C por 24 h. Con los datos obtenidos se generaron gráficas de distribución de tallas y se establecieron las relaciones morfométricas del ostión para cada sitio de cultivo. La biomasa total presente en cada sarta se estimó multiplicando el número de organismos vivos de una talla determinada presentes en la sarta por el peso entero vivo estimado a partir de las relaciones alométricas. El porcentaje de mortalidad acumulativa (M) se calculó de acuerdo a la siguiente ecuación (García-Esquivel et al., 2000):
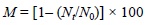
donde N, = número de ostiones vivos al tiempo t, y N0 = número inicial de ostiones vivos por sarta.
En 1998 se presentaron mortalidades masivas de ostión en BSQ, por lo que en noviembre de ese año se realizó una evaluación puntual de la mortalidad de una cohorte de ostión en 6 sitios de BF y uno adicional del brazo este (fig. 1). Los sitios de muestreo se seleccionaron con base a un registro de fechas de fijación y siembra mantenido por los ostricultores. En los lugares de muestreo se recolectaron cinco sartas con ostiones adultos (1 sarta/estante), en las cuales se cuantificó el total de organismos vivos y muertos. Asimismo, en cada sarta se midió (dependiendo de la densidad) la longitud de 30 organismos vivos y un número similar de organismos muertos, y se calcularon las fechas aproximadas de mortalidad del ostión de la siguiente manera: el incremento en longitud del ostión de BF es lineal durante los primeros nueve meses post-fijación (García-Esquivel et al., 2000), por lo que se supuso que en 1998 los organismos del sitio 1 y 6 mantuvieron las mismas tasas de crecimiento lineal que en 1995-1996 (0.23 y 0.36 mm d-1, respectivamente); una vez que se comprobó la coincidencia entre la longitud estimada (a partir de las tasas de crecimiento de 1995-1996) y la longitud de los ostiones vivos presentes en los sitios 1 y 6, se dividió la longitud de los organismos muertos entre la tasa de crecimiento "esperada" (0.23 mm d-1 y 0.36 mm d-1). Para el resto de los sitios se calculó la tasa "esperada" como sigue: (0.23 mm d-1 + 0.36 mm d-1)/2. El dato resultante representa el número total de días que los organismos permanecieron vivos desde el momento de la fijación, por lo que combinado con la fecha de siembra se pudo estimar la fecha probable de mortalidad de los mismos para cada sitio. La biomasa por sarta se estimó combinando los datos de distribución de frecuencias (1998) y relaciones alométricas.
Para determinar si el evento de mortalidad masiva persitía en 2000, en enero de ese año se repartieron cinco sartas en la boca y cinco en la cabecera de BF (1 sarta por estante) con juveniles de 20.6 ± 1.2 mm de longitud. Aproximadamente cada dos meses se realizaron mediciones de longitud y densidad de organismos vivos/muertos en cada una de las sartas marcadas, con lo cual se calculó la tasa de crecimiento y la supervivencia del ostión en cada sitio.
]]> Tasas de ingestión y efecto del ostión sobre el alimento disponibleEn julio y octubre de 1999, y en enero y abril de 2000 se realizaron series de tiempo de 6 a 8 h de duración con el fin de evaluar las tasas de ingestión y biodepositación de C. gigas en BSQ. Las determinaciones fueron realizadas en tierra firme, a una distancia aproximada de 5 m con respecto al canal principal (fig. 1). Se utilizó el sistema de charolas europeo (Hawkins et al. 1996; Iglesias et al., 1998; Bayne et al., 1999). Brevemente, este consistió en utilizar cuatro charolas de plástico con tres divisiones (celdas) individuales (fig. 2). Las charolas se mantuvieron con flujo abierto (150 a 200 mL min-1) mediante bombeo del agua de mar desde una profundidad subsuperficial de 0.5 m del canal principal. En cada celda se colocó un ostión y un divisor de plástico en el sentido antero-posterior de los ostiones con el fin de separar las heces y seudoheces producidas (fig. 2).

En cada fecha se utilizaron 12 ostiones de talla comercial (93 ± 14 mm) previamente aclimatados por 24 h en las charolas experimentales. Al final de ese período se limpiaron las celdas y se inició la colecta de biodepósitos. Las heces y seudoheces se colectaron cada 1.5 h (cada hora en julio) utilizando una manguera de plástico que funcionó como sifón. Las muestras (200 mL) fueron filtradas a través de filtros precombustionados de fibra de vidrio GF/C, enjuagadas con formato de amonio al 2.75% (para eliminar sales) y congeladas para su posterior procesamiento en el laboratorio. También cada hora se hicieron recolectas por triplicado del agua de entrada utilizando una llave de paso extra, y las muestras fueron filtradas en la forma indicada anteriormente. Al final de los experimentos se midió la longitud de los ejemplares con un vernier y se disectó el tejido, el cual se secó a 100°C durante 48 h para determinar el peso seco. Los filtros fueron secados a 90°C por 48 h para obtener el peso seco (material particulado total, TPM) de las heces (f), seudoheces (ps) y seston (s). Posteriormente se combustionaron en una mufla a 450°C durante 4 h para obtener el peso de las cenizas (PIM, material inorgánico particulado). El material orgánico particulado (POM) de cada muestra se obtuvo por diferencia entre el peso seco y su correspondiente PIM.
Las tasas de filtración (FR, mg POM h-1) e ingestión orgánica neta (NOIR, mg POM h-1) se calcularon para cada fecha de acuerdo a las siguientes ecuaciones (Hawkins et al., 1996, Bayne et al., 1999):

donde t = período de medición (h). Para eliminar el efecto de la talla (peso), las tasas fisiológicas se estandarizaron mediante la ecuación alométrica (Bayne et al., 1999):

donde Ys es la tasa fisiológica (FR, NOIR) estandarizada, Ws es el peso de un organismo estándar, We es el peso observado del ostión experimental, Ye la tasa fisiológica observada y b es el exponente para la tasa de filtración. En este estudio se usó un peso estándar de 3.5 g tejido seco (equivalente al peso promedio de los organismos experimentales) y un valor de b de 0.439 (Bougrier et al., 1995).
]]> La superficie y profundidad del brazo BF se estimó en base a una carta batimétrica (Dirección General de Oceanografía Naval, Secretaría de Marina, México), corregida con mediciones de profundidad in situ durante el presente estudio. Las características de la marea se obtuvieron de datos históricos previamente reportados para BSQ (Ocampo-Torres, 1980). El prisma de mareas se calculó para BF como la diferencia de volumen entre la marea alta y baja (Dyer, 1997). El porcentaje de intercambio de agua se calculó dividiendo el volumen de agua de salida entre el volumen total de la laguna (18.7 x 106 m3), referido al nivel medio del mar (Carver y Mallet, 1990).El alimento disponible en BF (Ad, g POM/m3) se calculó multiplicando el prisma de mareas por la concentración de POM. Finalmente, la biomasa de ostiones de una talla dada que puede soportar BF (Bo, Kg) se calculó utilizando la aproximación de Carver y Mallet (1990), modificada con porcentajes que permiten ajustes en la disponibilidad de alimento:

donde 25% = máximo porcentaje de alimento permisible para el ostión, suponiendo que este valor no declinaría significativamente la oferta alimenticia en BF (Uribe y Blanco, 2002); 85% = porcentaje de ajuste para la NOIR del ostión, debido a que el 15% del tiempo (mareas bajas) no consumen alimento; Wwt = peso entero vivo del ostión (g) y 1000 = factor usado para expresar la biomasa en Kg.
Resultados
Biomasa y mortalidad
El patrón de dispersión de tallas fue similar en las sartas experimentales (fig. 3a, b) y comerciales (fig. 3c), con un patrón de distribución normal (prueba de Kolmorogov-Smirnov, P > 0.20) y una amplia dispersión. Al momento de la cosecha (12 meses post-fijación) las sartas comerciales de la boca contenían el 67% de los organismos con tallas > 90 mm, mientras que solo el 21% de los ostiones cultivados en la cabecera alcanzaron esas tallas durante el mismo período (fig. 3c). Se encontró una relación alométrica de tipo potencial entre la longitud y el peso entero vivo o el peso del tejido seco del ostión (fig. 4a, c), y una relación lineal entre el peso entero vivo y el peso del tejido seco (fig. 4b). Estos patrones fueron similares en los ostiones cultivados en la boca y en la cabecera de BF, pero las magnitudes de las constantes fueron distintas. El exponente alométrico fue mayor en los ostiones de la boca (b = 2.68) que en los de la cabecera (b = 2.49 a 2.58). Asimismo, la pendiente de la relación longitud vs. peso vivo fue 1.5 veces mayor en los ostiones de la boca (m = 0.037) que en los de la cabecera (m = 0.025).

Los ostiones cultivados en la boca de BF alcanzaron la talla comercial media (90 mm) de los 9 a los 12 meses post-fijación, y fueron entre 1.3 y 1.9 veces más grandes que los de la cabecera, respectivamente (tabla 2). La producción por sarta disminuyó drásticamente en 1998, en comparación con 1995-1996 (tabla 2). En la boca de BF el promedio de organismos vivos por sarta disminuyó de 24 (febrero) y 54 (abril) en 1996, hasta 10 ostiones en 1998 (tabla 2). Asimismo, en la cabecera el número de organismos por sarta disminuyó de 66 (febrero) y 48 (abril) en 1996, hasta 9 ostiones en 1998. En términos de biomasa, ésta varió entre 0.81 y 2.81 kg/sarta en la boca de BF (tabla 2), equivalente a 89 y 309 kg peso entero vivo/estante, respectivamente. Al combinar la información del número de estantes instalados (9000) en BF, el promedio máximo de ostiones producidos por sarta (54 ostiones) y la biomasa a los 12 meses post-fijación (talla media de 10 cm) se obtiene un potencial productivo de 483 kg ostión estante-1 año-1 o 4347 t año-1. El patrón espacial en las tasas de crecimiento se mantuvo similar entre 1995 y 2000, con valores más altos en la boca que en la cabecera de BF (fig. 5). No obstante, en 2000 las tasas de crecimiento (0.18 y 0.08 mm d-1 para la boca y la cabecera de BF, respectivamente) disminuyeron hasta un 50%-60% con respecto a 1995-96. Asimismo, la mortalidad acumulada de ostiones mayores de 2 cm fue mayor en 2000 (21-29%, fig. 5) que en 1995 (0-6%).

Efecto del ostión sobre el alimento disponible
La concentración media de POM durante las fechas de medición de TF y NOIR varió entre 0.7 y 2.5 mg L-1 (tabla 3), con un promedio general de 1.9 mg L-1. Los porcentajes más altos de POM con respecto al seston total se observaron en octubre y abril (25.5% y 24%, respectivamente) y los más bajos (17.8%) en enero (tabla 3). A pesar de estas variaciones los ostiones regularon sus tasas de filtración y mantuvieron la tasa de ingestión orgánica neta (NOIR) relativamente constante en todos los muestreos (2.6 a 3.9 mg POM ostión-1 h-1), con un promedio general de 3.15 mg POM ostión-1 h-1 (tabla 3).
En el presente trabajo se estimó que el brazo BF tiene una profundidad media de 2.1 m; una superficie de 7.5 x 106 m2, y un volumen de 18.7 x 106 m3, referidos al nivel medio del mar. La influencia de las mareas se vió reflejada en el volumen total de intercambio entre BF y el oceano, con valores mínimos de 6% y máximos de 76% a lo largo de un ciclo quincenal. El prisma de mareas (P) estimado para el ciclo quincenal fue de 9.35 x 106 m3, mientras que los valores estimados separadamente para el período de mareas vivas y muertas fueron de 15.36 x 106 m3 y 4.92 x 106 m3, respectivamente. Utilizando la concentración media de POM (1.9 mg L-1) y la NOIR media (3.15 mg POM ostión-1 h-1 = 75 mg d-1) observada en ostiones de talla comercial (9 cm, 52.2 g peso entero vivo), se estimaron valores de B0 = 5974, 1913 y 3636 t de ostión para mareas vivas, muertas y ciclo quincenal, respectivamente.
Discusión
Rendimiento y mortalidad
]]> Los resultados del presente estudio coinciden con lo reportado previamente (García-Esquivel et al., 2000) en relación al tiempo en que el ostión alcanza la talla media comercial en BSQ. No obstante, esta es la primera vez que se muestra la variabilidad de las tallas del ostión cosechado en ese lugar, y los resultados sugieren que es necesario aumentar el tiempo de cultivo hasta 12 meses (boca) o más de 2 años (cabecera) para poder obtener una eficiencia del 70% de ostiones con talla comercial (90 mm). La gran variabilidad de las tallas al momento de la cosecha es una consecuencia natural del método de cultivo (sartas suspendidas), el cual requiere un mínimo de mantenimiento a lo largo de todo el período de engorda. A pesar de esa ventaja, el cultivo comercial de ostión en BF presenta serias desventajas técnicas que se reflejaron en el presente estudio, y que podrían incidir en el rendimiento y los costos de producción e incluyen: (1) control deficiente de densidades al momento de fijar la larva (García-Esquivel et al., 2000), lo cual puede resultar en la subutilización del sustrato y de la superficie de cultivo; (2) aumento del tiempo de cultivo para obtener un porcentaje adecuado (~70%) de ostiones de talla comercial; (3) volúmenes reducidos de un producto de talla uniforme. En conjunto, estas desventajas representan un costo económico que necesita ser reevaluado. Un estudio comparativo llevado a cabo en la década de los ochenta sugirió que el estante era el arte de cultivo más rentable para BF, seguido de las canastas y balsas (Ramos-Amézquita, 1987). No obstante, las estimaciones del autor se basaron en el número total de organismos, por lo que es necesario un nuevo análisis en función del porcentaje de organismos que alcanzan la talla comercial al momento de la cosecha. Por otra parte, los valores consistentemente bajos de las constantes alométricas y lineales coinciden con los bajos índices de condición reportados previamente para los ostiones cultivados en la cabecera (García-Esquivel et al., 2000). Lo anterior indica que los ostiones de la boca de BF no solo crecen más rápido, sino que contienen más carne (tejido) que los ostiones de talla similar cultivados en la cabecera de BF, por lo que se recomienda utilizar las ecuaciones correspondientes para realizar comparaciones de rendimiento sitio-específicas basadas en la talla del organismo.Las mortalidades masivas de ostión ocurridas en BSQ a partir de 1998 son las de mayor intensidad que se tengan registradas hasta ahora, y se tradujeron en una caída significativa de la producción total. Estudios previos indican que la mortalidad "normal" del ostión cultivado en BSQ es cercana a cero para organismos mayores de 3 cm de longitud (García-Esquivel, 2000). Los valores de mortalidad observados en 1998 y 2000, así como las tallas en que ésta ocurrió (tabla 1) y su persistencia actual (Héctor González, Acuacultores de San Quintín, com. pers.) sugieren que este es un fenómeno extraordinario para BSQ cuya causa aún no ha sido identificada. Es de notar que la mayor mortalidad se presentó hacia el centro de BF (fig. 1, tabla 1) y coincidió con la zona donde se introdujeron alrededor de 1000 t de almeja Chione sp. durante el período de estudio, proveniente del Golfo de California (Depto. Acuacultura, SAGARPA, Ensenada, BC). Hasta ahora se han sugerido varios factores como posibles responsables de la mortalidad en BF, incluyendo: (a) carga de patógenos nocivos asociados a Chione sp. (García-Esquivel et al. 1999); (b) heterozigocidad reducida del ostión cultivado, que se traduciría en una baja capacidad para amortiguar los cambios ambientales de temperatura (Correa et al., este volumen); (c) cambios en la composición taxonómica y/o abundancia del fitoplancton, inducidos por El Niño (García-Esquivel et al., 1999); y (d) competencia por alimento ejercida por Chione sp. A pesar de que se ha demostrado que el ostión cultivado en BSQ presenta una heterozigocidad reducida con respecto a poblaciones silvestres (Correa et al., este volumen), hasta ahora no existe ningún reporte científico donde se haya probado alguna de las hipótesis mencionadas arriba, por lo que es necesario plantear una estrategia integral para evaluar la magnitud y persistencia de la mortalidad de ostión en BSQ, así como para identificar el o los factores responsables del fenómeno. También se reportaron mortalidades inusuales de juveniles-adultos de Crassostrea gigas durante 1998-2000 en Sonora (J. Isidro Flores, Gobierno del Estado de Sonora, México, com. pers.) y en la Bahía de Tomales, California (Martin Strain, Point Reyes Organic Oyster Co.; John Finger, Hog Island Oyster Co., com. pers.). Datos recientes reportados para la costa oeste de Estados Unidos (EUA) sugieren que existen diferencias ploidia-específicas y sitio-específicas en la magnitud de la mortalidad de C. gigas, con mayor mortalidad asociada a ostiones triploides (Cheney et al., 2000). Asimismo, las diferencias sitio-específicas en la costa oeste de EUA parecen deberse no solo a la presencia/ausencia de microalgas tóxicas, sino también al estrés causado por episodios cortos de altas temperaturas y bajas concentraciones de oxígeno que cambian con el ciclo de mareas y la estación de año (Cheney et al., 2000). Más recientemente se reportó que el origen/stock de progenitores también influyó en las mortalidades de C. gigas registradas en Puget Sound y Tomales Bay (Cheney et al., 2001).
Efecto del ostión sobre el alimento disponible
Idealmente el cálculo de la capacidad de carga de una laguna costera debe considerar la evaluación in situ de varios factores hidrodinámicos, geoquímicos y biológicos que cambian en forma simultánea a lo largo del tiempo. Cuando la hidrodinámica es gobernada por corrientes de marea es posible estimar la influencia de los filtroalimentadores sobre el alimento disponible, una vez que se conocen el volumen de renovación del agua y la actividad fisiológica de los organismos (Ryther, 1969; Cabanas et al., 1979; Carver y Mallet, 1990). El supuesto básico es que el alimento disponible en el volumen de entrada es independiente del alimento que sale en el ciclo mareal previo (Carver y Mallet, 1990).
En el presente estudio la variación en la concentración de seston tuvo una influencia mínima sobre la tasa de ingestión neta (NOIR) de C. gigas (tabla 3). Esto contrasta con las observaciones de Barillé et al. (1997), quienes encontraron que a concentraciones in situ de seston menores de 25 mg L-1 la tasa de ingestión de C. gigas fue proporcional a la carga de partículas, pero a concentraciones entre 25 y 100 mg L-1 se mantuvo constante. Las discrepancias probablemente se deban a la plasticidad fisiológica de C. gigas para adaptarse a distintos ambientes, ya que las variaciones en la concentración media de seston reportadas por los autores durante un ciclo de mareas vivas y muertas para la Bahía de Marens Oleron (25 a 400 mg L-1) contrastan sensiblemente con las de BSQ (tabla 3). Los datos obtenidos en el presente estudio sugieren que el promedio generalizado de tasas de ingestión utilizado fue apropiado para los cálculos sobre la capacidad de carga. Asimismo, el establecimiento de una cuota máxima para el ostión del 25% del alimento disponible en BF es justificable si tomamos en cuenta que éste no debe disminuir por debajo de 50%, y que existen otros pastoreadores importantes en el medio (e.g., zooplancton) cuyo impacto aún no ha sido evaluado. Con este criterio, Uribe y Blanco (2002) establecieron una cuota de 8% del alimento disponible en Bahía Tongoy, Chile, para estimar el capacidad de carga de la almeja Argopecten purpuratus en esa bahía. Se sabe que el microzooplancton es capaz de reducir hasta en un 50% el alimento disponible en cuerpos costeros (Halvorse et al., 2001; Susuki et al., 2002). Es de notar que debido a que no se incluyó el tiempo de residencia del agua en el presente estudio, es deseable utilizar los valores de biomasa ostrícola estimados para mareas muertas (1913 t peso entero vivo), ya que esto reduciría el riesgo de subestimar el impacto de los filtroalimentadores. Dicho valor es cercano a las 2000 t de ostión que estimaron Camacho-Ibar et al. (2003) para BF a partir de balances de carbono, suponiendo una eficiencia de transferencia trófica del 10%.
Con todo lo anterior, el presente estudio sugiere que el potencial de producción de las artes de cultivo instaladas en BF (4300 t) está en el límite o por encima del límite de la capacidad de carga de BF. La disminución de hasta un 50% en la tasa de crecimiento de C. gigas observada en 2000 con respecto a 1995-1996 (tabla 3) también coincide con esta sugerencia. No obstante, la producción de ostión registrada durante el período de estudio no sobrepasó las 600 t año-1 (Depto. Acuacultura, SAGARPA) por lo que la reducción de la tasa de crecimiento no podría ser explicada sólamente por una capacidad de carga ostrícola rebasada, a menos de que la biomasa de otros filtroalimentadores no esté siendo considerada correctamente, o que la calidad del alimento disponible para el ostión haya cambiado. En este sentido, se desconoce de manera precisa la biomasa de almeja Chione sp. que se introdujo en la zona de cultivo durante el período de estudio, y no existen datos sobre la calidad del fitoplancton (tipo y abundancia de especies) en ese lapso, por lo que cualquier conclusión sobre la influencia de uno u otro factor sería especulativa. Por lo anterior, se recomienda mantener un monitoreo cercano de las tasas de crecimiento de C. gigas, ejercer un mayor control sobre el manejo de moluscos en BSQ, y extender la medición de tasas de pastoreo en los filtroalimentadores más representativos del cuerpo de agua con el fin de identificar el impacto de esos grupos sobre la capacidad de carga de la laguna.
Agradecimientos
Este trabajo fue parcialmente financiado por los proyectos SIMAC (ref. 980106020) y UABC (ref. 4047), así como por la empresa integradora Acuacultores de San Quintín S.A. Se agradece especialmente el apoyo logístico recibido por las empresas Acuícola San Quintín S.A., Ostiones Guerrero S.A. y J.C. Cota. Conrado Rivera, Salvador Galindo, Martín Hernández y Ricardo Vidal apoyaron en los trabajos de campo durante algunas partes del estudio. El trabajo mejoró sustancialmente con las sugerencia de 3 revisores anónimos.
]]> Referencias
Barillé, L., Prou, J., Héral, M. and Razet, D. (1997). Effects of high natural seston concentrations on the feeding, selection, and absorption of the oyster Crassostrea gigas (Thunberg). J. Exp. Mar. Biol. Ecol., 212: 149-172. [ Links ]
Bayne, B.L., Hedgecock, D., McGoldrick, D. and Rees, R. (1999). Feeding behaviour and metabolic efficiency contribute to growth heterosis in Pacific oysters [Crassostrea gigas (Thunberg)]. J. Exp. Mar. Biol. Ecol., 233: 115-130. [ Links ]
Bougrier, S.P., Geairon, P., Deslous-Pauli, J.M., Bacher, C. and Jonquires, G. (1995). Allometric relationships and effect of temperature on clearance and oxygen consumption rates of Crassostrea gigas (Thunberg). Aquaculture, 134: 143-154. [ Links ]
Cabanas, J.M., González, J.J., Marino, J., Perez, A. y Román, G. (1979). Estudio del mejillón y de su epifauna en los cultivos flotantes de la Ría de Arosa. III. Observaciones previas sobre la retención de partículas y la biodepositación de una batea. Bol. Inst. Esp. Oceanogr., 5: 44-52. [ Links ]
Camacho-Ibar, V.F., Carriquiry, J.D. and Smith, S.V. (2003). Non-conservative P and N fluxes and net ecosystem production in San Quintin Bay, Mexico. Estuaries (aceptado). [ Links ]
Carver, C.E. and Mallet, A.L. (1990). Estimating the carrying capacity of a coastal inlet for mussel culture. Aquaculture, 88: 3953. [ Links ]
Cheney, D.P., MacDonald, B.F. and Elston, R. (2000). Summer mortality of Pacific oysters, Crassostrea gigas (Thunberg): Initial findings on multiple environmental stressors in Puget Sound, Washington, 1998. J. Shellfish Res., 19: 353-359. [ Links ]
Chávez de Nishiwaka, A.G. y S. Alvarez Borrego. 1974. Hidrología de la Bahía de San Quintín, Baja California, en invierno y primavera. Cienc. Mar., 1(2):31-62. [ Links ]
Cheney, D.P., Elston, R., MacDonald, B., Kinnan, K. and Suhrbier, A. (2001). Summer mortality of the Pacific oyster Crassostrea gigas: Influences of culture methods, site conditions, and stock selection. J. Shellfish Res., 20: 542 (abstract). [ Links ]
Correa, F., Collins, E., Oceguera, A., Domínguez, D. y Cordero, B. (2004). Variación alozímica del ostión japonés, Crassostrea gigas en la Bahía de San Quintín, BC, México. Cienc. Mar. 30(1A): 99-107. [ Links ]
Dyer, K.R. (1997). Estuaries. 2nd ed., John Wiley, New York, 195 pp. [ Links ]
García-Esquivel, Z., Galindo-Bect, M.S., González-Gómez, M.A., Hernández-Ayón, M., Ley-Lou, F. y Vidal-Talamantes, R.V. (1999). Mortandad de ostión en Bahía San Quintín durante 1998: Una evaluación puntual. Reporte Técnico, Instituto de Investigaciones Oceanológicas, Universidad Autónoma de Baja California, 22 pp. [ Links ]
García-Esquivel, Z., González-Gómez, M.A., Galindo-Bect, S. and Hernández-Ayón, M. (2000). Microgeographic differences in growth, mortality and biochemical composition of the Pacific oyster (Crassostrea gigas) from San Quintín Bay, northwest Mexico. J. Shellfish Res., 19: 789-798. [ Links ]
Halvorsen, E., Pedersen, O.P., Slagstad, D., Tande, K.S., Fileman, E.S. and Batten, S.D. (2001). Microzooplankton and meso-zooplankton in an upwelling filament off Galicia: Modeling and sensitivity analysis of the linkages and their impact on the carbon dynamics. Prog. Oceanogr., 51: 499-513. [ Links ]
Hawkins, A.J.S., Smith, R.F.M., Bayne, B.L. and Heral, M. (1996). Novel observations underlying the fast growth of suspension-feeding shellfish in turbid environments: Mytilus edulis. Mar. Ecol. Prog. Ser., 131: 179-190. [ Links ]
Hernández-Ayón, J. M., Galindo-Bect, M. S., Camacho-Ibar, V.F., García-Esquivel, Z., González-Gómez, M.A. y Ley-Lou, F. (2004). Dinámica de los nutrientes en el brazo oeste de Bahía San Quintín, Baja California, México, durante y después de El Niño 1997/1998. Cienc. Mar. 30(1A): 129-142. [ Links ]
Hily, C. (1991). Is the activity of benthic suspension feeders a factor controlling water quality in the Bay of Brest? Mar. Ecol. Prog. Ser., 69: 179-188. [ Links ]
Iglesias, J.I., Urrutia, M.B., Navarro, E. and Ibarrolla, I. (1998). Measuring, feeding and absorption in suspension-feeding bivalves: An appraisal of the biodeposition method. J. Exp. Mar. Biol. Ecol., 219: 71-86. [ Links ]
Juárez-Villarreal, M.M. (1982). Modelo de dispersion para un estuario ramificado y su aplicación a Bahía San Quintín. Tesis de maestría, Centro de Investigación Científica y de Educación Superior de Ensenada, BC, México. 79 pp. [ Links ]
Lara-Lara, J.R., Alvarez-Borrego, S. and Small, L.F. (1980). Variability and tidal exchange of ecological properties in a coastal lagoon. Est. Coast. Shelf Sci., 11: 613-637. [ Links ]
Martori-Oxamendi, J.I. (1989). Variabilidad de la circulacion y sus causas en Bahia San Quintín, BC. Tesis de maestría, Centro de Investigación Científica y de Educación Superior de Ensenada, BC, México, 110 pp. [ Links ]
Ocampo-Torres, F.J. (1980). Análisis de mareas y predicción de velocidad mediante un modelo unidimensional en Bahía San Quintín, BC.. Tesis de licenciatura, Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada, BC, México. [ Links ]
Polanco, J.E., Mimbela, S.R., Beléndez, M.L., Flores, M.A. y Reinoso-Alvarez, A.L. (1988). Situación actual del cultivo de ostión japonés (Crassostrea gigas) en el noroeste de México. En: Situación Actual de las Principales Pesquerías Mexicanas. Secretaría de Pesca, México, pp. 219-248. [ Links ]
Ramos-Amézquita, H.R. (1987). Determinación del arte más rentable para el cultivo a nivel comercial del ostión japonés (Crassostrea gigas), en la Bahía de San Quintín. Tesis de licenciatura, Universidad Autónoma de Baja California, Ensenada, BC, México, 157 pp. [ Links ]
Ryther, J.H. (1969). The potential of the estuary for shellfish production. Proc. Natl. Shellfisheries Assoc., 59: 18-22. [ Links ]
Susuki, K., Tsuda, A., Kiyosawa, H., Takeda, S., Nishioka, J., Saino, T., Takahashi, M. and Wong, C.S. (2002). Grazing impact of microzooplankton on a diatom bloom in a mesocosm as estimated by pigment-specific dilution technique. J. Exp. Mar. Biol. Ecol., 271: 99-120. [ Links ]
Uribe, E. y Blanco, J.L. (2002). Capacidad de los sitema acuáticos para el sostenimiento del cultivo de pectínidos: El caso de Argopecten purpuratus en la bahía Tongoy, Chile. En: A.N. Maeda-Martínez (ed.), Los Moluscos Pectínidos de Iberoamérica: Ciencia y Acuicultura. CYTED, pp. 233-248. [ Links ]
]]>