Se estudió el comportamiento temporal de la enfermedad pudrición blanca, causada por el hongo Sclerotium cepivorum, en tres lotes de cultivo de cebolla, ubicados en el Rancho Agua Nueva, municipio de Juventino Rosas, Guanajuato, con densidades de inóculo (DI) de 0.021, 0.052 y 0.44 esclerocios g–1 de suelo, catalogadas como baja, media y alta densidad. Las primeras plantas con síntomas se observaron 30 d después del transplante (ddt) y las últimas a los 160 ddt. Para este tiempo la incidencia acumulada fue 51.93, 67.75 y 82.9% para las DI baja, media y alta. La relación entre las curvas del progreso de la enfermedad y la fenología del cultivo mostraron que a mayor densidad de inóculo la incidencia máxima de la enfermedad (ymax) se presenta en etapas más tempranas. El progreso temporal de la pudrición blanca fue descrita por el modelo de crecimiento Gompertz cuando la DI fue 0.021 y 0.052 esclerocios g–1 de suelo, y por el modelo Monomolecular cuando la DI fue 0.44 esclerocios g–1 de suelo. La comparación de epidemias se realizó tomando en cuenta las tasas de crecimiento homologadas al modelo Gompertz (Rho) y mediante el área bajo la curva del progreso de la enfermedad (ABCPE), mostrándose diferencias significativas (p<0.01). La DI inicial de S. cepivorum en el suelo determina características epidemiológicas importantes como la tasa de incremento de la enfermedad (r), la forma de la curva, el estado fenológico de máxima incidencia (ymax) y la incidencia final de enfermedad (yf ); características útiles para entender y predecir el desarrollo de la enfermedad en el campo y tomar decisiones de manejo.
Palabras clave: Epidemiología, modelos de crecimiento.
Abstract
Temporal behavior of white rot disease, caused by the fungus Sclerotium cepivorum, was assessed on three onion plots, located at Rancho Agua Nueva, municipality of Juventino Rosas, Guanajuato, México. The plots had different soil inoculum density (ID) of the pathogen: 0.021, 0.052, and 0.44 sclerotia g–1 soil, classified as low, medium, and high. The first plants with symptoms were observed 30 d after transplanting (dat), and the last diseased plants were recorded 160 dat. At this time, accumulated disease incidence was 51.93, 67. 75, and 82.9%, corresponding to low, medium, and high ID. The relationship between disease progress curves and crop phenology showed that at the highest soil ID, maximum disease incidence (ymax) occurs at the earliest phenology stages. Temporal progress of white rot at low and medium ID (0.021 and 0.052 sclerotia g–1 soil) was described by Gompertz growth model, while high ID (0.44 sclerotia g"1 soil) was described by the monomolecular model. The comparison of epidemics was performed taking into account the growth rates homologized to the Gompertz model (Rho) and by the area under the disease progress curve (AUDPC) showing significant differences (p<0.01). Initial ID of S. cepivorum in soil determines important epidemiological characteristics like disease increase rate (r), form of disease progress curve, phenological stage of maximum incidence (ymax), and final disease incidence (yf ), useful for understanding and predicting disease development in field and taking decisions on disease management.
Key words: Epidemiology, growth models.
]]>
INTRODUCCIÓN
El Estado de Guanajuato es el principal productor de cebolla (Allium cepa L.) en México, con 26% de la producción (SIAP–SAGARPA, 2002). Sin embargo, el cultivo está fuertemente afectado por la enfermedad pudrición blanca, ocasionada por el hongo Sclerotium cepivorum Berk. Esta enfermedad causa serios daños en los cultivos de cebolla y ajo y en otras especies del género Allium. Se han realizado diversas investigaciones sobre S. cepivorum enfocadas principalmente a su control en el cultivo de la cebolla, pero no suficientes estudios epidemiológicos que ayuden a describir y entender el comportamiento temporal de la enfermedad. Cano (2003)[1] estudió la cantidad de inóculo presente en el suelo y el progreso temporal de epidemias durante dos ciclos de cultivo de ajo con una variedad diferente en cada uno, encontrando que la cantidad de inóculo presente en el suelo tiene un efecto directo en el porcentaje de incidencia de la enfermedad. En el cultivo de la cebolla es también importante realizar estudios epidemiológicos para un mejor entendimiento de la dinámica temporal de la pudrición blanca y su interacción con el hospedante. El objetivo del presente trabajo fue determinar y comparar el desarrollo temporal de la incidencia de la enfermedad pudrición blanca, inducida por S. cepivorum, en el cultivo de la cebolla bajo tres diferentes densidades de inóculo del patógeno en el suelo, en relación con el desarrollo fenológico del cultivo.
MATERIALES Y MÉTODOS
Lugar de estudio
Este trabajo se realizó de diciembre de 1989 a mayo de 1990, en el Estado de Guanajuato, municipio de Juventino Rosas, en el Rancho Agua Nueva. El clima predominante fue semiseco con lluvias en verano, temperatura media anual entre 18 y 22 °C. Se usaron tres lotes de cultivo con 0.021, 0.052 y 0.44 esclerocios g–1 de suelo. La extracción de esclerocios desde el suelo se hizo con la técnica de Papavizas (1972). Estos lotes de clasificaron como de baja, media y alta densidad de inóculo (DI). El trabajo de laboratorio se hizo en el Laboratorio de Ecología de Fitopatógenos del Suelo del Colegio de Postgraduados.
Establecimiento del cultivo
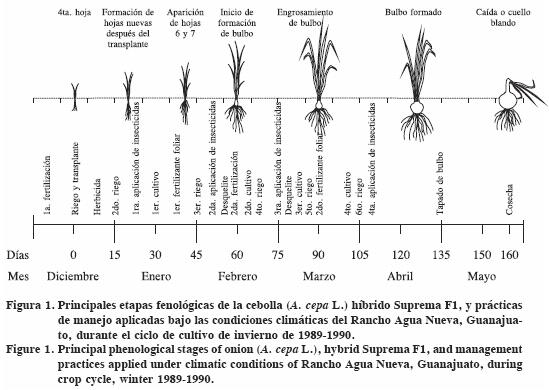
El cultivo se estableció en el ciclo invierno–primavera, de mediados de diciembre a fines de mayo. Dentro de cada lote se trazaron cuadrantes, cada uno con tres surcos con una separación de 0.90 m por 2.0 m de largo (5.4 m2 cuadrante–1). Cada cuadrante se delimitó con estacas e hilo. La superficie que abarcó cada lote fue 2160 m2, 972 m2 y 1890 m2 con 400, 180 y 350 cuadrantes para las DI baja, media y alta. Se sembró el híbrido de cebolla Suprema F1, trasplantado a doble hilera con distancia de 8 a 10 cm entre plantas y 15 entre hileras, dando un promedio de 100 plantas por cuadrante. Las prácticas de manejo fueron las típicas de la región, e incluyeron fertilización, riego, aplicación de herbicidas e insecticidas, cultivo, desquelite, tapado de bulbo y cosecha. También se registraron las etapas fenológicas del cultivo con base en la experiencia en campo, y de acuerdo con Brewster (2001).
Medición de la enfermedad
]]>
La evaluación de la enfermedad se realizó mediante censos a los 30, 45, 65, 81, 102, 124, 138 y 160 d después del transplante (ddt). En cada censo se registró la incidencia de plantas con síntomas de amarillamiento y marchitez de las hojas. Las plantas enfermas se marcaron con banderillas de diferente color por fecha; el registro se llevó por cuadrante.
Variables climáticas
El registro mensual de temperatura ambiental (°C), temperatura del suelo sin cubierta (°C) y precipitación pluvial (mm) se obtuvo de la estación meteorológica del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) ubicado en Celaya, Guanajuato, situada a 0.5 km del lugar de estudio.
Análisis del progreso temporal de la curva de la enfermedad
Los datos de incidencia se usaron para obtener la curva de progreso de la enfermedad acumulada y absoluta en el tiempo; también se ajustaron a modelos de crecimiento en su forma linearizada: logístico, 1n(y /(1– y) = 1n[y0 /(1– y0)] + rLt; Gompertz, –1n(–1n y) = –1n(–1n y0) + rGt; y monomolecular, 1n [1 / (1– y)] = 1n [1 / (1– y0)] + rMt (Campbell y Madden, 1990). Los parámetros de tasa: modelo logístico (rL), modelo Gompertz (rG) y modelo monomolecular (rM), se estimaron con el Modelo Lineal General (GLM) usando el método de mínimos cuadrados y el paquete estadístico SAS. Se eligió el modelo que describió mejor el crecimiento de la enfermedad en los tres diferentes lotes con base en el coeficiente de determinación (R ), el cuadrado medio del error (CME), el error estándar de los estimadores de los parámetros y y sus probabilidades, así como en el comportamiento de los residuales. Para estandarizar las tres epidemias a un mismo modelo y compararlas, se calculó el parámetro de tasa absoluta media ponderada (Rho), con la ecuación Rho (ρ) = r*/2m+2, donde r* adquiere los valores de la tasa de infección aparente del modelo monomolecular, Gompertz y logístico, y m adquiere los valores 0, 1, y 2. Una vez calculada Rho, ésta se sustituyó en la ecuación anterior para calcular r*, la cual correspondió al valor de la tasa de infección aparente del modelo menos frecuente homologada al modelo de mayor frecuencia de ajuste (Campbell y Madden, 1990). Una vez homologadas las tasas de crecimiento de las diferentes epidemias éstas se compararon mediante una prueba de t para determinar diferencias estadísticas significativas al 0.01 (Infante y Zárate, 1990).
También se compararon de las epidemias mediante la estimación del área bajo la curva del progreso de la enfermedad (ABCPE), aplicando el método de integración trapezoidal (Campbell y Madden, 1990) y usando la ecuación: donde n es el número de mediciones de la enfermedad en el tiempo; el término (yiyyi+1) / 2 es el punto medio entre (yiyyi+1) que representa la cantidad de enfermedad en un intervalo de tiempo, correspondiente a la altura de cada rectángulo; y el término (ti+1 – ti) representa el tiempo (d) entre dos evaluaciones de la enfermedad e indica la anchura de cada rectángulo. Los cálculos se hicieron usando el paquete estadístico SAS. Se realizó un análisis de varianza para comparar el ABCPE de las tres epidemias con base en la prueba de Tukey (p<0.05).
RESULTADOS Y DISCUSIÓN
Los datos para el presente trabajo se generaron en 1990, por lo que la información pudiera parecer obsoleta, pero no lo es porque se debe considerar que las epidemias que induce S. cepivorum en el cultivo de cebolla, una vez establecido el patógeno, son fenómenos biológicos que están siempre presentes en el agroecosistema. Son también fenómenos dinámicos caracterizados por su estructura espacial y temporal, descripción a la que se dedica el presente trabajo. Para apreciar el valor de la información generada y analizada en el presente trabajo, así como su actualidad, valdría la pena preguntar. ¿Qué condiciones podrían haber cambiado respecto a la estructura y el comportamiento de las epidemias de S. cepivorum en el Bajío de 1990 a la fecha?
1) Las variedades cultivadas en la región no han cambiado y no cambiarán mientras siga la dependencia de semillas importadas, que son precisamente las más vulnerables a la enfermedad.
]]>
2) Las condiciones climáticas y edáficas conducentes al desarrollo de las epidemias no han cambiado.
3) El manejo agronómico no ha cambiado (monocultivo continuo) y es el que logra la máxima productividad del agroecosistema, aunque conduce también a la expresión máxima de la epidemia.
En la búsqueda de literatura pertinente al tema no se encontró, de 1990 a la fecha, ningún trabajo parecido siquiera al presente, tal vez porque, por ocurrir en el suelo, un cuerpo opaco que no permite ver los fenómenos que en él ocurren, el seguimiento de la dinámica poblacional de S. cepivorum puede hacerse sólo mediante métodos de investigación indirectos (muestreos, extracción, cuantificación) que no son particularmente preferidos por los investigadores.
Desarrollo fenológico de la cebolla
El ciclo de cultivo desde el transplante hasta la cosecha duró 160 d de diciembre a mayo. Se registraron las siguientes etapas fenológicas según Brewster (2001): desarrollo de la cuarta hoja, formación de hojas nuevas, aparición de las hojas seis y siete, inicio de formación de bulbo, engrosamiento de bulbo, bulbo formado y caída de cuello o cuello blando (Figura 1).
Clima
Las medias mensuales de temperatura ambiental (°C), temperatura del suelo sin cubierta (°C), y precipitación (mm) durante el ciclo de cultivo se presentan en el Cuadro 1. Las temperaturas ambiental y del suelo aumentaron durante el ciclo de cultivo de 13.6 a 21.15 °C y de 9.9 a 21 °C. Las mayores precipitaciones se registraron en mayo (72.2 mm) y junio (76.9 mm).
Incidencia de la enfermedad
]]>
En las tres densidades de inóculo (alta, media y baja) las primeras plantas con síntomas de la enfermedad se observaron a los 30 ddt y el último registro de plantas enfermas fue a los 160 ddt en la cosecha.
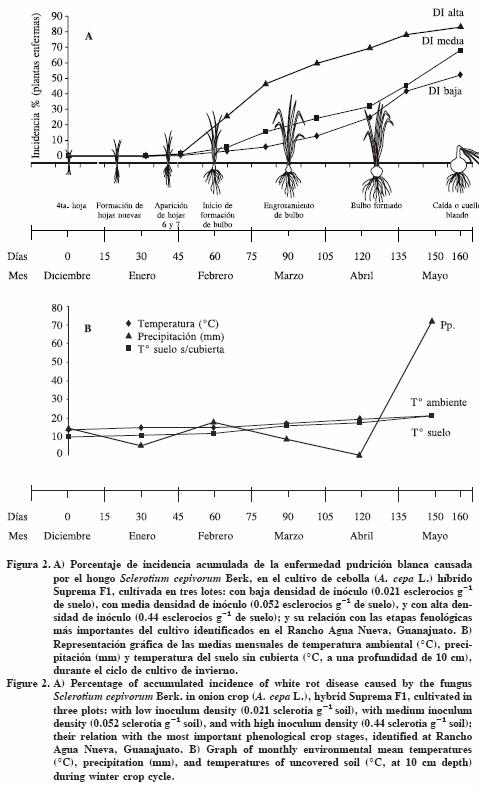
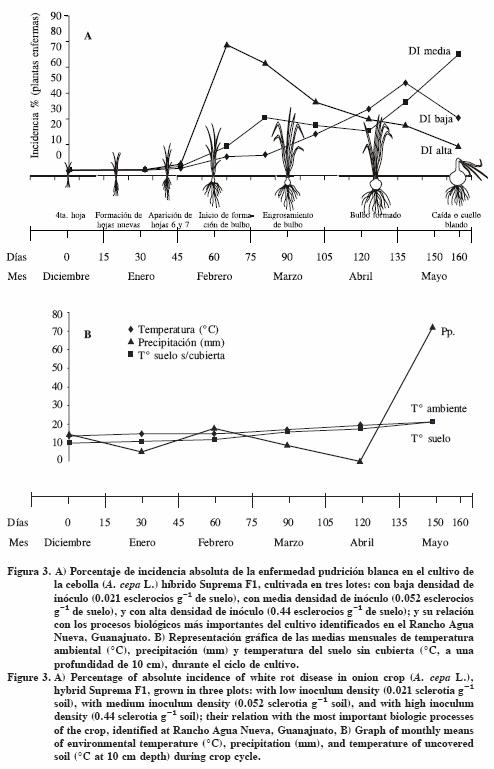
La epidemia en el lote con DI baja se inició a los 30 ddt registrándose una incidencia inicial de 0.03%, a los 45 ddt aumentó la cantidad de enfermedad llegando a 0.36% de incidencia, la cual coincidió con la aparición de las hojas 6 y 7. La cosecha se realizó a los 160 ddt y se registró una incidencia acumulada de 51.93% de plantas enfermas (Cuadro 2; Figura 2). La curva de incidencia absoluta (Figura 3) para esta epidemia indicó que a los 138 ddt hubo la mayor cantidad de plantas enfermas con 11.85% de incidencia, correspondiendo con la etapa de bulbo formado (Cuadro 3; Figura 3).
En el lote con DI media, la epidemia se inició también a los 30 ddt, registrándose una incidencia inicial de 0.087%. De manera similar a lo sucedido en el lote de DI baja, a los 45 ddt se observó un incremento de 0.76% en la cantidad de enfermedad que coincidió con la aparición de las hojas 6 y 7; mientras que a la cosecha (160 ddt) se registró una incidencia acumulada de 67.75% (Cuadro 2; Figura 2). La gráfica de incidencia absoluta en este lote de DI media (Figura 3) reveló dos picos con alta incidencia de plantas enfermas: el primero a los 81 ddt (etapa de engrosamiento de bulbo) con 10.27 % y el otro en la cosecha (160 ddt) con 22.47% de incidencia, coincidiendo con el período fenológico de caída o cuello blando del cultivo (Cuadro 3, Figura 3).
En el lote con DI alta, la epidemia se inició a los 30 ddt con 0.128% de incidencia. A los 45 ddt se observó un incremento de 1.3% en la enfermedad, que coincidió con la aparición de las hojas 6 y 7; a la cosecha (160 ddt) se registró una incidencia acumulada de 82.9% (Cuadro 2; Figura 2). La curva de incidencia absoluta (Figura 3) mostró un incremento de la enfermedad de 24.4% de incidencia entre los 45 y 81 ddt coincidiendo con las etapas de inicio de formación de bulbo y de engrosamiento del bulbo (Cuadro 3; Figura 3). En los tres lotes de cultivo, la epidemia inició a los 30 ddt, en la etapa de formación de hojas nuevas. También se observó una relación directa entre la densidad de inóculo (DI) inicial y la incidencia final acumulada, ya que con DI iniciales de 0.021, 0.052 y 0.44 esclerocios g-1 de suelo las incidencias finales fueron 51.93, 62.75 y 82.9% respectivamente.
La temperatura del suelo es el elemento climático que más influye en la germinación y desarrollo de S. cepivorum, así como de la enfermedad que causa. Crowe y Hall (1980) encontraron que en el campo, la germinación e infección de S. cepivorum ocurre entre los 9 y 24 °C con un óptimo de 15 °C y su viabilidad disminuye en un intervalo de 21 a 30 °C. Adams y Papavizas (1971) mencionan que los esclerocios son la primera fuente de inóculo y éstos infectan en cualquier etapa de crecimiento del hospedante, si se presentan temperaturas de 15 a 25 °C, con un óptimo de 20 °C, mientras que el desarrollo de la infección tiene un intervalo de 10 a 18 °C y un óptimo de 15 °C. En el presente estudio, de diciembre a junio hubo temperaturas de 9 a 21 °C, lo cual fue apropiado para la germinación e infección de S. cepivorum en el hospedante, coincidiendo con lo reportado por los dos autores mencionados. La temperatura de 21 °C al final de su ciclo pudieron participar en la disminución de la viabilidad e infección en el hospedante.
Ajuste de curvas de progreso de la enfermedad
]]>
El ajuste de los datos de incidencia de la enfermedad para las tres densidades de inóculo a los modelos de crecimiento monomolecular, logístico y Gompertz en su forma linearizada, permitió elegir el modelo que mejor describe a las tres epidemias. Los criterios estadísticos considerados para evaluar y seleccionar los modelos se presentan en el Cuadro 4.
En la DI baja, el progreso de la incidencia de la pudrición blanca fue descrito satisfactoriamente por el modelo Gompertz con: R2=98.9, CME=0.007, =0.025, =0.0002. La distribución de residuales fue aleatoria, es decir, los valores predichos del modelo son muy cercanos a los valores reales de incidencia. El modelo de regresión obtenido fue: – 1ln [ –1n (y)] = –2.59 + 0.018 (t), donde –2.59 es el intercepto, 0.018 es la pendiente y t es el tiempo. En la DI media, el modelo que mejor describió los datos de la enfermedad fue el Gompertz con: R2=97.6, CME=0.017, =0.054, =0.0005 y una distribución de residuales aleatoria. El modelo de regresión obtenido fue: – 1n [–1n (y)] =–2.497+ 0020 (t) donde –2.497 es el intercepto, 0.020 es la pendiente y t es el tiempo. En la DI alta, el progreso temporal de la incidencia fue mejor descrito por el modelo mono–molecular con: R2=98.4, CME=0.007, =0.026, =0.0002, así como una distribución de residuales aleatoria. El modelo de regresión obtenido fue: 1n (1/1–y) 0.565 + 0.014)(t), donde –0.565 es el intercepto, 0.014 es la pendiente y t es el tiempo.
En el presente estudio se encontró que las epidemias en los lotes con baja y media DI se ajustaron al modelo Gompertz, que es un modelo sigmoidal. Crowe y Hall (1980) mencionan que las hifas de esclerocios germinados crecen a través del suelo infectando a raíces que están a 1 y 2 cm de distancia. Cuando la densidad de inóculo es baja y el patógeno tiene movilidad limitada, es el crecimiento de las raíces el que incrementa la probabilidad de contacto entre el patógeno y el hospedero, por lo que la curva de crecimiento sigmoidal de la raíz explica la curva de progreso sigmoidal de la enfermedad (Campbell et al., 1984). En el lote con alta DI hubo mayor cantidad de esclerocios y aparentemente una infección explosiva temprana, en las primeras etapas fenológicas, presentándose muerte masiva de plantas, lo cual evitó que hubiera mayor producción de tejido sano disponible para futuras infecciones. Con estos resultados se puede inferir que las diferencias en la DI del patógeno en el suelo van a influir en la forma de la curva de progreso temporal de la epidemia, es decir, determinan el tiempo (o etapa fenológica), la máxima cantidad de enfermedad y la cantidad del daño causado al cultivo.
Las tasas de crecimiento de la enfermedad estimadas para los sitios en estudio con DI baja, media y alta fueron 0.018, 0.02 y 0.014. Dado que los parámetros de cambio se derivaron a partir de diferentes modelos, se homologaron usando el parámetro Rho o tasa absoluta media ponderada, resultando 0.004, 0.005 y 0.007 para DI baja, media y alta. Estos valores permitieron calcular la tasa de infección aparente homologada al modelo de mayor frecuencia de ajuste, en este caso el de Gompertz obteniendo valores de rG de 0.018, 0.020 y 0.028 para los lotes con DI baja, media y alta (Cuadro 5).
Comparación de epidemias
Para comparar las tasas homologadas al modelo Gompertz se usó la prueba de t para comparación de dos parámetros (Infante y Zárate, 1990) (Cuadro 6). Los resultados mostraron que la comparación de las tasas de densidad baja y media, media y alta, baja y alta, fueron estadísticamente distintas (p<0.01), es decir, las tres densidades de inóculo de S. cepivorum seleccionados en este trabajo provocaron epidemias significativamente diferentes. Al parecer, la tasa de crecimiento de la enfermedad depende de la DI existente en el suelo: cuando ésta es mayor, la tasa de crecimiento aumenta.
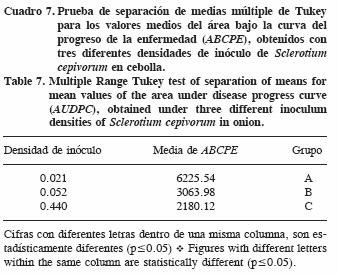
La comparación de las tres epidemias mediante el área bajo la curva del progreso de la enfermedad (ABCPE) indicó que la DI alta tuvo la mayor área (6225.54), seguida por la DI media (3063.98) y la más baja fue para la DI baja (2180.12) (Cuadro 7).
La comparación de epidemias (o curvas del progreso de la enfermedad) es una práctica común en epidemiología. En este caso permitió corroborar la relación indicada de a mayor densidad de inóculo la incidencia de la enfermedad es mayor. Por tanto, se considera que la DI inicial es importante para entender y quizá predecir el desarrollo de la enfermedad dentro de una población de plantas en campo y para tomar decisiones y el establecimiento de estrategias de manejo del patógeno y de la enfermedad. El análisis temporal de las epidemias y sus componentes (planta, patógeno y ambiente) permitió entender que la densidad de inóculo inicial del patógeno altera la intensidad de la epidemia, la tasa del progreso de la enfermedad (r), la forma de la curva del progreso de la enfermedad, el tiempo en que se presenta la cantidad máxima (ymax) de enfermedad y la enfermedad final observada (yf).
]]>
CONCLUSIONES
El inicio de la enfermedad ocurrió en la formación de hojas nuevas después del transplante, independientemente de la DI. La relación entre las curvas del progreso de la enfermedad y la fenología, demostraron que a mayor densidad de inóculo, la máxima incidencia absoluta (ymax) se presenta en etapas más tempranas de desarrollo del cultivo. Las curvas del progreso de la enfermedad se ajustaron mejor, para el caso de la densidad de inóculo baja y media, al modelo Gompertz; y la alta al monomolecular.
La comparación de epidemias basadas en las tasas de crecimiento homologadas al modelo Gompertz, así como en el ABCPE, indicaron diferencias estadísticamente significativas entre las tres diferentes densidades de inóculo, correspondiendo a la densidad de inóculo alta la mayor tasa de crecimiento y la mayor ABCPE.
Las condiciones de temperatura ambiental y del suelo registradas en la zona de estudio son adecuadas para la germinación e infección de S. cepivorum en el hospedante durante el ciclo de cultivo invierno–primavera (diciembre–mayo). El análisis temporal de la enfermedad y sus componentes (planta, patógeno y ambiente) mostró que la DI inicial determina características epidemiológicas importantes como: tasa de progreso (r), intensidad de la enfermedad, tiempo de ymax y cantidad de enfermedad final (yf), en este pato–sistema.
LITERATURA CITADA
Adams, P. B., and G. C. Papavizas. 1971. Effect of inoculum density of Sclerotium cepivorum and some soil environmental factors on disease severity. Phytopathology 61: 1253. [ Links ]
Brewster, J. L. 2001. Las Cebollas y otros Alliums. Editorial Acribia S. A. Zaragoza, España. pp: 21–43. [ Links ]
Campbell, C. L., and L. V. Madden. 1990. Introduction to Plant Disease Epidemiology. Wiley & Sons. USA. 532 p. [ Links ]
Campbell, C. L., R. W. Jacobi, N. T. Powell, and C. E. Main. 1984. Analysis of disease progression and the randomness of occurrence of infected plants during tobacco black shank epidemics. Phytopthology 74: 230–235. [ Links ]
Crowe, F. J., and D. H. Hall. 1980. Vertical distribution of Sclerotia of Sclerotium cepivorum and host root systems relative to white rot of onion and garlic. Phytopathology 70: 70–73. [ Links ]
Infante–Gil, S., y G. P. Zárate de Lara. 1990. Métodos Estadísticos. Ed. Trillas, México. 643 p. [ Links ]
Papavizas, G. C. 1972. Isolation and enumeration of propagules of Sclerotium cepivorum from soil. Phytopathology 62:545–549. [ Links ]
SIAP, SAGARPA. 2002. Avance comparativo de siembras y cosechas a nivel nacional. Cultivo de Cebolla. www.siap.org.mx (consultado el 30 de septiembre de 2002). [ Links ]
NOTAS
1 Cano S., A. 2003. Análisis epidemiológico de la enfermedad "pudrición blanca" inducida por Sclerotium cepivorum Berk. en el cultivo del ajo (Allium cepa L.) en Cortazar, Edo. de Guanajuato. Tesis de M. en C. Instituto de Fitosanidad, Programa de Fitopatología. Colegio de Postgraduados. Montecillo, Edo. de México. 112 p. [ Links ] ]]>1971611253200121-431990532198474230-23519807070-731990643197262545-549SIAPSAGARPA20022003112
 y
y  y sus probabilidades, así como en el comportamiento de los residuales. Para estandarizar las tres epidemias a un mismo modelo y compararlas, se calculó el parámetro de tasa absoluta media ponderada (Rho), con la ecuación Rho (ρ) = r*/2m+2, donde r* adquiere los valores de la tasa de infección aparente del modelo monomolecular, Gompertz y logístico, y m adquiere los valores 0, 1, y 2. Una vez calculada Rho, ésta se sustituyó en la ecuación anterior para calcular r*, la cual correspondió al valor de la tasa de infección aparente del modelo menos frecuente homologada al modelo de mayor frecuencia de ajuste (Campbell y Madden, 1990). Una vez homologadas las tasas de crecimiento de las diferentes epidemias éstas se compararon mediante una prueba de t para determinar diferencias estadísticas significativas al 0.01 (Infante y Zárate, 1990).
y sus probabilidades, así como en el comportamiento de los residuales. Para estandarizar las tres epidemias a un mismo modelo y compararlas, se calculó el parámetro de tasa absoluta media ponderada (Rho), con la ecuación Rho (ρ) = r*/2m+2, donde r* adquiere los valores de la tasa de infección aparente del modelo monomolecular, Gompertz y logístico, y m adquiere los valores 0, 1, y 2. Una vez calculada Rho, ésta se sustituyó en la ecuación anterior para calcular r*, la cual correspondió al valor de la tasa de infección aparente del modelo menos frecuente homologada al modelo de mayor frecuencia de ajuste (Campbell y Madden, 1990). Una vez homologadas las tasas de crecimiento de las diferentes epidemias éstas se compararon mediante una prueba de t para determinar diferencias estadísticas significativas al 0.01 (Infante y Zárate, 1990). donde n es el número de mediciones de la enfermedad en el tiempo; el término (yi y yi+1) / 2 es el punto medio entre (yi y yi+1) que representa la cantidad de enfermedad en un intervalo de tiempo, correspondiente a la altura de cada rectángulo; y el término (ti+1 – ti) representa el tiempo (d) entre dos evaluaciones de la enfermedad e indica la anchura de cada rectángulo. Los cálculos se hicieron usando el paquete estadístico SAS. Se realizó un análisis de varianza para comparar el ABCPE de las tres epidemias con base en la prueba de Tukey (p<0.05).
donde n es el número de mediciones de la enfermedad en el tiempo; el término (yi y yi+1) / 2 es el punto medio entre (yi y yi+1) que representa la cantidad de enfermedad en un intervalo de tiempo, correspondiente a la altura de cada rectángulo; y el término (ti+1 – ti) representa el tiempo (d) entre dos evaluaciones de la enfermedad e indica la anchura de cada rectángulo. Los cálculos se hicieron usando el paquete estadístico SAS. Se realizó un análisis de varianza para comparar el ABCPE de las tres epidemias con base en la prueba de Tukey (p<0.05).