
Siete nuevas especies de Malpighiaceae neotropicales
William R. Anderson †
University of Michigan Herbarium. 3600 Varsity Drive, Ann Arbor, Michigan 48108, U.S.A. Address correspondence to Christiane Anderson: chra@umich.edu
Recibido en febrero de 2014. ]]> Aceptado en mayo de 2014.
ABSTRACT
Seven new species of neotropical Malpighiaceae are described: Aspicarpa steinmannii, Bunchosia breedlovei, and Bunchosia cruciana from Mexico; Bunchosia soejartoi, Lophanthera marcelae, and Mascagnia corymbosa from Colombia; and Heteropterys tiinae from Peru.
Key words: Aspicarpa, Bunchosia, Colombia, Heteropterys, Lophanthera, Malpighiaceae, Mascagnia, Mexico, Peru.
RESUMEN
Se describen como nuevas siete especies de Malpighiaceae neotropicales: Aspicarpa steinmannii, Bunchosia breedlovei y Bunchosia cruciana de México; Bunchosia soejartoi, Lophanthera marcelae y Mascagnia corymbosa de Colombia; y Heteropterys tiinae de Perú.
Palabras clave: Aspicarpa, Bunchosia, Colombia, Heteropterys, Lophanthera, Mascagnia, Malpighiaceae, México, Perú.
]]> Study of neotropical Malpighiaceae continues to reveal undescribed species, especially in large and difficult genera, such as Bunchosia H.B.K. The novelties here described were discerned during a review of Bunchosia in Mexico, a visit to herbaria in Colombia in connection with the forthcoming Catálogo de las Plantas de Colombia, and examination of collections sent as gifts to the University of Michigan Herbarium.
Aspicarpa steinmannii W. R. Anderson, sp. nov. (Fig. 1).
Aspicarpa steinmannii differs from all other species of Aspicarpa by the combination of a shrubby habit with very small leaves lacking stipules, floral bracts equalling the vegetative leaves, chasmogamous flowers borne at the apex of branches, and a samara with the three equal wings.
Much-branched shrub 1 m tall; stems initially densely sericeous, the older stems ca. 2 mm in diameter, partially glabrescent but with some hairs persistent. Leaves mostly opposite or subopposite but sometimes alternate or ternate; lamina of larger leaves 7-15 mm long, 2-4 mm wide, narrowly ovate (elliptical), rounded at base, acute at apex, loosely sericeous on both sides with the very fine hairs 2-3 mm long, straight, appressed or somewhat spreading, the abaxial surface with some hairs persistent, the adaxial surface glabrescent in age; glands absent from surface of lamina but sometimes represented by one columnar or clavate marginal structure on each side, 0.2-0.3 mm long, dark red or purplish, borne slightly above base; lateral veins not visible on either side in dried leaves; petiole 0.5-1 mm long, sericeous, eglandular; stipules not seen. Chasmogamous flowers borne in pairs at tips of vegetative stems; bract subtending each flower like a vegetative leaf but generally smaller, 4-9 mm long, 1.5-2 mm wide; peduncle 3-10 mm long, shorter in flowers, elongating as fruits mature; bracteoles borne at or slightly below apex of peduncle, spreading, resembling miniature leaves 1.5-3 mm long, 0.8-1.5 mm wide, ovate, deciduous in fruit; pedicel 3-7 mm long, shorter in flowers, elongating as fruits mature; cleistogamous flowers not found. Sepals abaxially densely tomentose and adaxially glabrous, incurved in anthesis, the 4 lateral sepals abaxially biglandular, the anterior sepal eglandular. Petals yellow, glabrous. Stamens 5, glabrous, opposite sepals; 3 stamens opposite anterior sepal and 2 posterior-lateral sepals fertile, bearing large anthers; 2 stamens opposite anterior-lateral sepals sterile, consisting only of a short filament without any sort of apical enlargement. Style 1, erect and straight, with a capitate terminal stigma, borne (presumably) on the anterior carpel. Intact fruits not seen; samaras alike, except samara coming from anterior carpel bearing persistent style; samara obcordate in outline, with three subequal wings (two lateral and one dorsal) 8-10 mm long, 3-4 mm wide, loosely hairy with some hairs persistent at maturity, especially on nut and base of wings; wings membranous with reticulate venation, distinct distally, connate proximally; each samara bearing a slender cartilaginous ventral carpophore from base along midline to place of attachment of samara to receptacle.
Type: MEXICO. Jalisco: Mun. Huejuquilla El Alto, sobre la brecha a La Ordeña y Rancho Nuevo, aprox. 1 km al NW de Vallecito (un rancho aprox. a 17 km al W de Huejuquilla El Alto sobre la carretera a San Juan Capistrano, Zacatecas), 22°40'28.9" N, 103°59'43.7" W, 6.XI.2012 (flower/fruit), R. Hernández-Cárdenas 778 with L. Arredondo-Amezcua (holotype: MICH!; isotype: IEB, fide V. Steinmann, pers. comm.).
Additional specimens examined: MEXICO. Jalisco: same locality as type, 17.X.2007 (flower), Steinmann & Ramírez-Amezcua 5939 (MICH); 24.XI.2011 (fruit), Steinmann et al. 7219 (MICH; also IEB, fide V. Steinmann, pers. comm.).
Tropical deciduous forest, 1232 m, collected with flowers and fruits in October and November.
The few collections of this species, all from the same population, bear only a few flowers, in such poor condition that they do not suffice for a complete description, which will have to be added when better material has been collected.
]]> The epithet of this fascinating new species honors Victor W. Steinmann (b. 1969), who first collected it and brought it to my attention. According to the traditional taxonomy of Malpighiaceae (e.g., Niedenzu, 1928), this species would be assigned to the genus Gaudichaudia H.B.K., but the phylogeny of Davis and Anderson (2010) gave convincing evidence that Aspicarpa Rich. and Gaudichaudia will have to be combined, so it seems best to describe this species under Aspicarpa, the older generic name.Aspicarpa steinmannii is immediately shown to belong in the Aspicarpa/Gaudichaudia complex by the following characters: leaves eglandular except for the tiny tail-like marginal outgrowths near the base of the lamina of some leaves; stamens five, opposite sepals; gynoecium with a single style, the stigma terminal and capitate; samaras with well-developed lateral wings; each samara attached to the receptacle by a cartilaginous carpophore. The fact that the mericarps are winged and borne on carpophores is the sole basis one could find for assigning the species to Gaudichaudia rather than Aspicarpa. Within Aspicarpa sens. lat., A. steinmannii is distinguished by the following combination of characters: much-branched shrub with very slender branches and tiny leaves; stipules apparently absent; cleistogamous flowers absent; chasmogamous flowers borne in pairs at tips of vegetative shoots; bracts hardly different from vegetative leaves; dorsal wing of samara identical to the two lateral wings. If one searches carefully through the described species of Aspicarpa and Gaudichaudia, one can find all or most of these characters (except perhaps for the production of flowers in pairs at the tips of vegetative stems), but taken together they are unmatched in any other species.
Bunchosia breedlovei W. R. Anderson, sp. nov.
Bunchosia breedlovei differs from B. veluticarpa by its larger laminas with more numerous lateral veins, eglandular petioles, inflorescences borne in the axils of the earliest deciduous leaves, and patchily glabrescent mature fruits.
Tree 18-24 m tall; stems densely appressed-tomentose or subsericeous with the vesture persistent or unevenly deciduous in first year, glabrescent in second year. Lamina of larger leaves 19-28.3 cm long, 10-16 cm wide, broadly elliptical or obovate, cuneate or rounded at base, flat or slightly revolute at margin, broadly obtuse or rounded and usually abruptly short-acuminate or apiculate at apex, adaxially glabrate at maturity except often appressed-tomentose on midrib and lateral veins near base, abaxially densely and persistently tomentose with the hairs T-shaped with the long crosspiece straight or sinuous, apparently eglandular, the principal lateral veins 10-13 on each side of midrib, very prominent abaxially and sunken adaxially in dried leaves; petiole 9-14 mm long, densely and persistently appressed-tomentose or sericeous, eglandular; stipules 1.5-3 mm long. Inflorescences simple, without vegetative leaves, borne near base of younger stems in axils of deciduous earliest leaves, mostly single but occasionally in pairs, 3.5-10 cm long, containing 6-18 flowers with the flowers decussate or distally in no consistent order, the axes densely and persistently appressed-tomentose or the pedicel glabrescent late in maturation of fruit; bracts 2-2.5 mm long, broadly ovate, abaxially sericeous or distally glabrescent, adaxially glabrous, eglandular; peduncle 0-2.5 mm long in fruit; bracteoles like bracts but smaller, 1-1.5 mm long, one of the pair sometimes bearing a large abaxial gland; pedicel 4.5-6 mm long in fruit. Sepals in fruit about as long as glands or slightly longer, rounded, thin, abaxially proximally sericeous and distally glabrous, adaxially glabrous; calyx glands 8, 3-3.5 mm long in fruit, obovate, 2 partially connate. Petals and stamens not seen. Gynoecium 3-carpellate; ovary densely tomentose, the wall very thick; styles connate their whole length. Largest fruits seen (still immature) 19-23 mm in diameter (dried), spheroidal or broadly 3-angled, densely and persistently tomentose but apparently patchily glabrescent near maturity, the wall granulate.
Type: MEXICO. Chiapas: Mun. Ocozocoautla de Espinoza, 18-20 km N of Ocozocoautla along road to Mal Paso, steep slope with montane rain forest, 800 m, 20.X.1971 (immature fruit), D. E. Breedlove 20958 (holotype: MICH!; isotypes: CHAPA!, DS!, ENCB!, F!, MEXU!, MO!, NY!, TEX!).
Additional specimens examined: MEXICO. Chiapas: Mun. Berriozábal, 13 km N of Berriozábal, fissured limestone ridge, lower montane rain forest, 1000 m, 2.XI.1971 (fruit), Breedlove 21642 (DS, ENCB, LL, MEXU, MICH, MO, NY).
This species is named for Dennis Eugene Breedlove (1939-2012), who, with A. R. Smith and R. F. Thorne, made the only known collections of it. Both were in fruit, so the flowers are not known, but in this genus the flowers are relatively uninformative except for the gynoecium, and the species is so distinctive that I am certain it is undescribed. Bunchosia breedlovei is to be compared to B. veluticarpa W. R. Anderson, a species of Costa Rica. Both taxa are (at least sometimes) tall trees, which is unusual in Bunchosia, and both have a densely tomentose three-carpellate gynoecium. In both the fruit is densely and tightly tomentose during enlargement, with the hairs more or less persistent in B. veluticarpa and apparently unevenly deciduous in B. breedlovei when the fruit is nearly full-sized; this persistent vesture on the enlarging fruits immediately distinguishes B. breedlovei from all other species of Bunchosia known from Mexico. The size of the fruits also distinguishes B. breedlovei from the majority of species in Mexico, which mostly have smaller fruits. Bunchosia veluticarpa differs from B. breedlovei in the following additional ways: the lamina of larger leaves is consistently smaller, especially narrower, 12-20.5 cm long, 5.5-8.5 cm wide; its principal lateral veins are only 7-9(11) on each side of the midrib, moderately raised below and flat or raised (not sunken) above in dried leaves; it has several obvious, well-defined abaxial glands at the base and many others scattered throughout the length of the lamina; the petiole often bears a small bulbous gland beside one or both stipules and a pair of impressed glands between middle and apex; and the inflorescences are mostly borne in the axils of current leaves. The laminas of B. breedlovei show scattered seemingly glandular anomalies hidden by the hairs, but these do not have the clear definition of the abaxial leaf glands usually found in Bunchosia and are probably wounds, not glands.
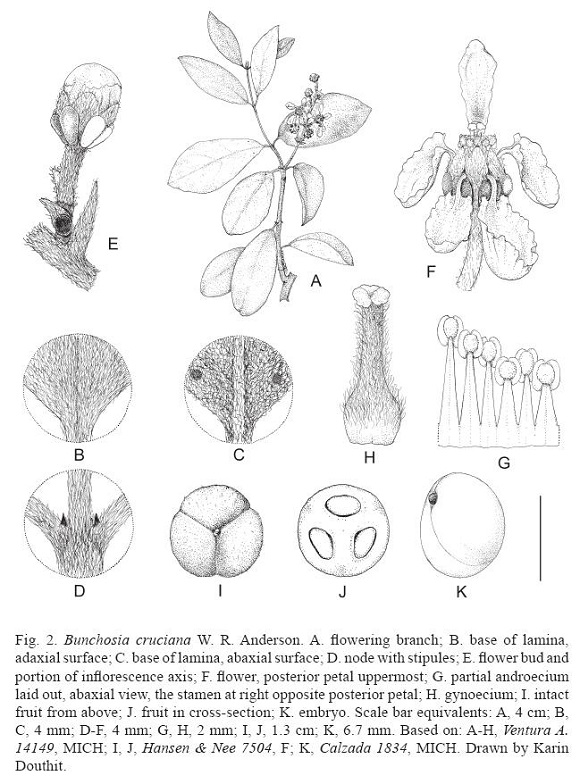
Bunchosia cruciana W. R. Anderson, sp. nov. (Fig. 2)
]]>
Bunchosia cruciana differs from B. biocellata in its smaller, adaxially velutinous laminas and its shorter petioles and inflorescences, from B. montana by its tricarpellate ovary.
Shrub or small tree 2-5(8) m tall; stems densely velutinous with subsessile or short-stalked V- or Y-shaped hairs in first year, glabrate in second year. Lamina of larger leaves 3.5-7.5(11.5) cm long, 2-3.5(6) cm wide, elliptical or somewhat ovate or obovate, cuneate, rounded, or somewhat cordate at base, flat at margin, obtuse or rounded (or rarely abruptly short-acuminate) at apex, adaxially persistently velutinous with the hairs V- or Y-shaped with a short stalk and straight branches, abaxially very densely and persistently tomentose with the hairs T-shaped with the long crosspieces sinuous or somewhat twisted, mostly biglandular abaxially near base with the glands beside midrib or between midrib and margin (very rarely several additional glands present distally in a row set in from margin), the principal lateral veins 5-8 on each side of midrib, prominent abaxially but neither prominent nor sunken adaxially; petiole 2-4(5) mm long, densely and persistently velutinous, eglandular; stipules 0.4-0.6(0.8) mm long. Inflorescences simple, without vegetative leaves, mostly borne on leafy stems but occasionally on leafless stems, in axils of current leaves or after fall of the subtending leaf, 1.5-6 cm long, containing 6-8(12) flowers with the flowers decussate or distally in no consistent order, the axes densely and persistently velutinous or tomentose or occasionally loosely sericeous; bracts 1.5-5 mm long, triangular or ovate, abaxially tomentose, adaxially glabrous, eglandular; peduncle 1-3 mm long; bracteoles like bracts but smaller, 1-2 mm long, one of the pair usually bearing a large abaxial gland; pedicel 3-7 mm long. Sepals 1.5-2.3 mm long beyond glands, 1.3-1.5 mm wide, broadly obtuse or rounded, abaxially tomentose or loosely sericeous over the whole surface or nearly glabrous toward margin and ciliate on margin, adaxially glabrous, appressed in anthesis; glands 8, 1.5-2.5 mm long, obovate, often compressed but distinct or rarely the glands of adjacent sepals connate, glabrous, often slightly detached at apex, the posterior 2 mostly slightly decurrent. Petals yellow, glabrous; lateral petals spreading or reflexed, with the claw 2-2.7 mm long and the limb 3-6 mm long and wide, flat to concave (especially the anterior pair), erose or coarsely toothed, eglandular all around margin; posterior petal spatulate, with the thickened claw erect, 2.5-4 mm long, widening distally without an apical constriction, the limb spreading to reflexed, 2.5-3.5 mm long, 2-3.2 mm wide, obovate, flat to recurved, erose and eglandular. Stamens glabrous; filaments 1.5-3.5 mm long, longest in front of flower and shortest in rear, connate for about 1/4-1/3 of their length; anthers 0.8-1 mm long, pressed against styles, the connective yellowish to brown, not or only slightly glandular-swollen abaxially. Gynoecium 3-carpellate; ovary 1.5-2 mm high, ovoid, tomentose or loosely sericeous; styles 3 but completely connate, tomentose, 1.5-2.2 mm long; stigma hardly to prominently 3-lobed. Fruit orange or red, 13-15 mm in diameter (dried), globose or depressed-globose and 3-lobed, thinly sericeous to glabrescent but even at maturity with some hairs persistent, the wall smooth (i.e., not granulate) but often with reticulate walls of pyrenes showing through the thin flesh.
Type: MEXICO. Veracruz: Mun. "Dos Ríos" (Emiliano Zapata), Palo Gacho, 300 m, 10.VII.1971 (flower/immature fruit), F. Ventura A. 3842 (holotype: MICH!; isotypes: DS!, ENCB!, TEX!).
Additional specimens examined: MEXICO. Veracruz: Mun. Actopan: Raya Manuel Díaz (Sierra Manuel Díaz), 17.IV.1985, Acosta P. 531 (XAL); Laguna de La Mancha, 10.VI.1977, Novelo 353 (MO, XAL); Mun. Apazapan: road from Baños de Carrizal to Emiliano Zapata, 2-6 km SE of Emiliano Zapata, 27.VI.1980, Hansen & Nee 7504 (F, MO, XAL); 3 km SW of Chahuapa on road to Apazapan, 12.VI.1984, Solheim & Cházaro B. 1588 (NY, WIS, XAL); Mun. Coatepec: 2 km antes de Jalcomulco en las faldas del Cerro de Las Palmas, 20.V.1979, Castillo C. & Tapia 699 (ENCB, F, GUADA, UC, XAL); Mun. Emiliano Zapata: 500 m de la desviación a Carrizal por la carreterra Xalapa - Veracruz, 20.VII.1975, Calzada 1834 (F, MEXU, MICH, XAL); Cerro Gordo, 30.IV.1972, Dorantes & Márquez 1493-D (ENCB, MICH); Plan del Río, carretera Veracruz - Xalapa, 8.VII.1977, Fay & Hernández 841 (F, GH, MEXU, NY, TEX, US, XAL); km 368-371 on Jalapa - Veracruz highway, between Cerro Gordo and Plan del Río, 13.II.1943, Gilly et al. 53 (MICH); just N of La Bocana, ca. 2.5 km S of El Aguaje, 30.VI.2001, Provance 3406 (MICH); La Cumbre, about 8 km NW of Rinconada, 23.VI.1984, Solheim 1684 (NY, WIS, XAL); Palo Gacho, 10.VII.1971, Ventura A. 3841 (CAS, ENCB, LL, MICH, TEX); Palo Gacho, 16.VI.1975, Ventura A. 11452 (ENCB, MICH); Plan del Río, 16.VII.1975, Ventura A. 11680 (ENCB, MICH); Mun. La Antigua: 2 km al W de Cardel, 27.VI.1970, González G. 32 (CAS, F, GH, MEXU); Mun. Paso de Ovejas: 2 km antes de Mata Mateo, 15.VII.1984, Avendaño R. & Castillo C. 1804 (UC, XAL); 2 km al NE de Acazonica, 23.VII.1985, Medina A. & Vázquez B. 298 (ENCB, IBUG, XAL); Mun. Puente Nacional: La Ceiba, 12.VI.1975, Ventura A. 11418 (ENCB, MICH); Río Escondido, 22.VI.1977, Ventura A. 14149 (ENCB, MICH); Mun. Xalapa: Rinconada near Jalapa, V.1894, Smith 1564 (NY); Mun. unknown: cerro a 11 km al S de Palma Sola, frente a Villa Rica, 29.VI.1972, Dorantes 1177 (CAS, ENCB, F, MEXU); Buena Vista, 19.VII.1972, Dorantes 1421 (F, LL, MEXU); Barranca de Panoaya, VII.1920, Purpus 8519 (UC); Remulatero, IV.1922, Purpus 8759 (UC).
Tropical deciduous forest or scrub vegetation from sea level to 550 m, collected with flowers from February to July and with fruits from April to July; local name "zapotito".
The specific epithet of Bunchosia cruciana refers to the Mexican state of Veracruz, where this species is endemic. Among Mexican species in which the ovary is tricarpellate and hairy and the enlarging fruit is glabrescent, B. cruciana should be compared to B. biocellata Schltdl., which is widespread in eastern Mexico and whose type also came from Veracruz. The easiest way to distinguish B. biocellata from B. cruciana is that in B. biocellata the laminas are initially sericeous adaxially with sessile, straight, more or less appressed hairs but soon glabrate except often persistently sericeous on the midrib. Its laminas are usually larger [(6)9-17 cm long, (4)5-9 cm wide)], its petioles and stipules are usually longer, its inflorescences are often longer and contain more flowers, and the fruits are often larger. Bunchosia montana A. Juss. resembles B. cruciana but its gynoecium is bicarpellate; it is known from Puebla, Oaxaca, Chiapas, Guatemala, and Honduras, but not from Veracruz.
Bunchosia soejartoi W. R. Anderson, sp. nov. (Fig. 3)
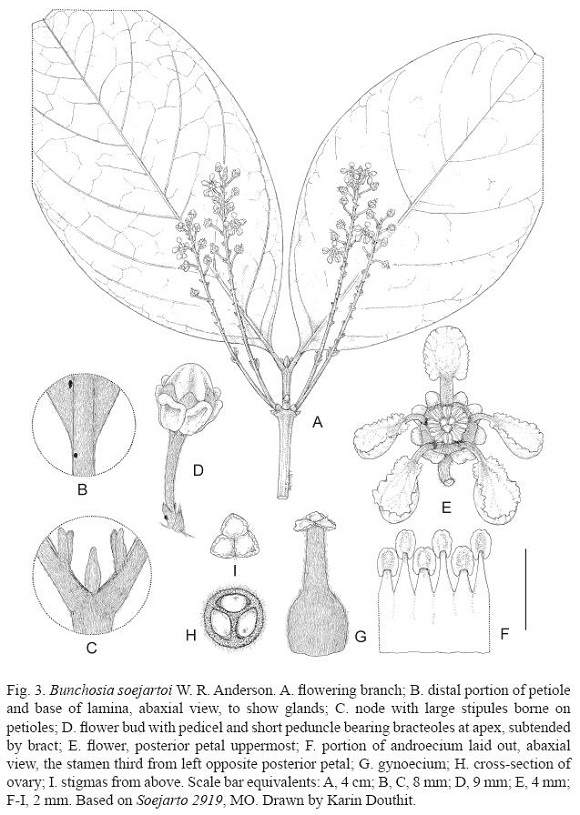
Tree 10 m tall; stems densely and persistently sericeous, glabrescent in later years with the advent of secondary growth. Lamina of larger leaves 16-26 cm long, 11-16 cm wide, broadly elliptical, broadly cuneate at base, rounded and apiculate or shallowly retuse at apex, adaxially glabrate at maturity except persistently loosely sericeous on midrib, abaxially densely and persistently appressed-tomentose with the hairs sessile or short-stalked and the crosspiece ca. 1 mm long and straight to sinuous but not twisted, the hairs numerous but not abundant enough to hide the epidermis, the abaxial glands small (ca. 0.5 mm in diameter) and few (up to 6 per side?), borne beside or near the midrib, the principal lateral veins 8-10 on each side; petiole 11-16 mm long, densely and persistently sericeous, eglandular at base, mostly bearing a pair of glands ca. 0.5 mm in diameter at or slightly below apex; stipules 2.5-3.5 mm long, narrowly triangular, flattened and often incurved distally, abaxially densely sericeous, adaxially thinly sericeous to glabrescent, borne on petiole 2-3 mm above base. Inflorescences borne in pairs on older stems axillary to scars of shed leaves, never in axils of current leaves; pseudoracemes 10-16 cm long with the slender axis sericeous, without vegetative leaves below the flowers, the flowers 16-34 or more, borne proximally in pairs but distally in no consistent arrangement; proximal bracts 3-4 mm long, distal bracts 1.5-2 mm long, triangular, abaxially sericeous, adaxially glabrous, all eglandular; proximal peduncles 2-4 mm long, distal peduncles 1-1.5 mm long, sericeous; bracteoles ca. 1 mm long, ovate, abaxially sericeous, adaxially glabrous, one of each pair bearing 1 abaxial gland and the other eglandular; pedicels 6-10 mm long, sericeous, glabrescent in age. Sepals extending 0.5-1 mm beyond glands, rounded, abaxially sericeous, adaxially glabrous, pressed inward against filaments; glands apparently 8 but probably actually 10 with the anterior 4 connate in 2 broad pairs, 2.5-3 mm long, obovate, compressed, decurrent with half or more of their length on pedicel, recurved at apex. Petals light yellow, glabrous; lateral petals reflexed, the claw 1.5-2 mm long, the limb 3-4.5 mm long, 3-4 mm wide, deeply to shallowly concave, erose, eglandular, the outermost largest and most deeply concave; posterior petal erect, the claw 2 mm long, the limb ca. 2.5 mm long, 2 mm wide, shallowly concave, erose or distally entire, eglandular. Stamens glabrous; filaments 1.5-2.5 mm long, longer opposite sepals than opposite petals, 1/2 or more connate; anthers 0.7-0.8 mm long, alike, the connectives yellow or light brown and slightly glandular-swollen. Gynoecium 3-carpellate, densely sericeous or appressed-tomentose its whole length including the style; ovary 1-1.2 mm high, globose, 3-loculed; style 1 (formed from 3 completely connate), 1.3-1.5 mm long, the 3 large stigmas distinct. Fruits unknown.
Type: COLOMBIA. Antioquia: Mun. Anorí, Corr. Providencia, Buenos Aires trail, along Río Anorí, 500-700 m, advanced and humid secondary forest, 7.VI.1971 (flower), D. D. Soejarto 2919 (holotype: GH!; isotypes: COL!, HUA!, MO!).
Bunchosia soejartoi is named in honor of the botanist Djaja Djendoel Soejarto (b. 1939), collector of the type and only known collection. It should be compared to other species that have a tricarpellate, densely hairy gynoecium. In Colombia, the only species that could be confused with B. soejartoi is B. retusa Triana & Planch., but in the latter the lamina is densely woolly below (not appressed-tomentose), the petioles are only ca. 5 mm long and biglandular at base, and the inflorescences are shorter, up to 10 cm long in flower, and borne singly in the axils of current vegetative leaves. In Central America the most similar species are B. lanieri S. Watson of Belize, Guatemala, and Honduras, and B. stipulacea W. R. Anderson of Panama. The most immediately evident distinction of B. soejartoi from those species is the inflorescences paired on older stems axillary to leaf scars; in the Central American species the inflorescences are single and axillary to current leaves. Bunchosia soejartoi is also distinguished by its rounded, often retuse leaf apices, few leaf glands, often sinuous abaxial leaf hairs, and stipules borne well above the base of the petiole. The flowers of B. soejartoi are smaller in all their parts than those of B. lanieri and B. stipulacea.
Heteropterys tiinae W. R. Anderson, sp. nov. (Fig. 4)
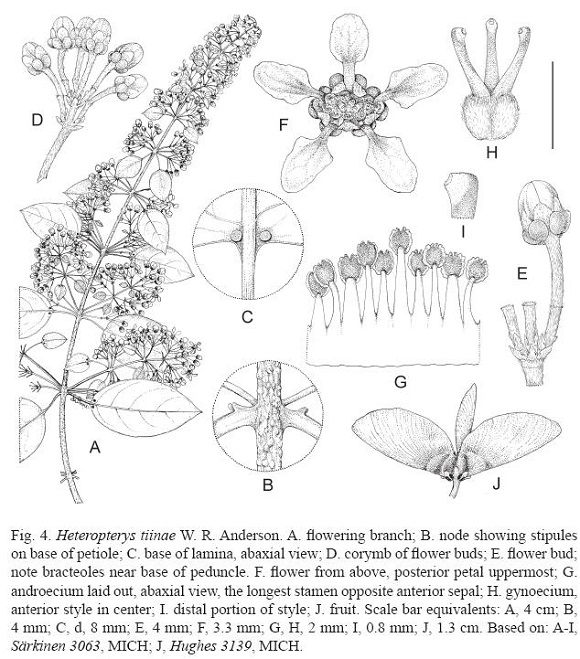
Heteropterys tiinae differs from all other species of Heteropterys by the combination of small glabrous laminas bearing large glands abaxially, epipetiolar stipules, and umbels or corymbs of 4–6 flowers with small greenish petals borne on well-developed peduncles.
Slender woody vine, the stem initially thinly sericeous with very short brown strongly appressed hairs, soon (already in first year) glabrescent and developing many large punctiform lenticels. Lamina of larger leaves 3.5-7.3 cm long, 1.9-3 cm wide, elliptical or narrowly ovate, cuneate to rounded and sometimes short-decurrent at base, flat or slightly revolute at margin, abruptly short-acuminate or obtuse and apiculate at apex, glabrous or bearing a few short appressed hairs abaxially near base, bearing 2 large raised glands on abaxial surface at base and occasionally 1 or 2 additional glands on abaxial surface above base, the glands rarely on decurrent laminar tissue and appearing to be on apex of petiole, the principal lateral veins 4-6 on each side of midrib; petiole of larger leaves 7-14 mm long, very sparsely sericeous to soon glabrate, eglandular except for glands on decurrent lamina (see above); stipules 0.2-0.4 mm long, thick, borne on petiole just above base, glabrous, persistent. Inflorescence axillary and terminal, a pyramidal panicle with flowers ultimately borne in umbels or corymbs of 4-6, the congestion variable with the proximal pair often separated by up to 2 mm from the terminal cluster, the axes thinly golden- or brownish-sericeous; inflorescence axes often 2 or 3 in same axil, arrayed vertically; bracts 0.5-1 mm long, ovate, appressed, eglandular or occasionally bearing a pair of tiny marginal glands near base; peduncle 1.5-3 mm long, always distinctly shorter than pedicel; bracteoles 0.4-0.7 mm long, ovate, appressed, eglandular or not infrequently bearing a pair of tiny marginal glands near base, borne very low on peduncle, up to 0.5 mm above bract; pedicel 3.5-5 mm long. Flowers ca. 8 mm in diameter. Sepals 1.5 mm long beyond glands, 0.8-1.1 mm wide, elliptical or broadly triangular, rounded at apex, appressed in anthesis, abaxially thinly sericeous, adaxially glabrous, the anterior eglandular, the lateral 4 biglandular with the glands green, swollen, rotund or elliptical, 1.1-1.6 mm long, glabrous. Petals exposed in the enlarging bud, yellow-green, glabrous, abaxially smooth or the lateral 4 slightly carinate; lateral petals spreading to reflexed in age, with claw 1-1.4 mm long and limb 2-2.3 mm long, ca. 1.5 mm wide, rounded-quadrangular or ovate, entire or slightly erose; posterior petal similar to lateral petals but with claw thicker and not recurved in age and limb slightly smaller. Stamens glabrous; filaments 1.5-2.2 mm long, connate proximally for 0.5-0.8 mm, longest opposite the anterior sepal and then opposite the 2 anterior-lateral sepals, stoutest opposite the 3 posterior petals; anthers 0.4-0.6 mm long, subequal, the connectives red. Ovary ca. 1 mm high, sericeous with very short tightly appressed hairs; styles 1.3-1.6 mm long, subequal with the anterior slightly longer than the posterior 2, glabrous, the anterior straight and erect or leaning slightly toward the posterior petal, the posterior 2 bowed, all 3 slightly flattened laterally and dorsally rounded-truncate at apex with the stigmas internal. Samaras 13-17 mm long, sparsely and loosely sericeous with the short hairs much more abundant and persistent on the nut than on the wing; dorsal wing 11-15 mm long, 6-8 mm wide; nut 3-5 mm long and wide, without lateral outgrowths.
Type: PERU. Cajamarca: between Jaén and Gotas de Agua, 5°41'61" S, 78°46'66" W, 800 m, roadside in area of disturbed scrubby seasonally dry forest, 13.IV.2008 (flower), T. E. Särkinen 3063 (holotype: MICH!; isotypes: E, MOL, fide C. E. Hughes, pers. comm.).
]]> Additional specimens examined: PERU. Cajamarca: Jaén, ca. 4 km NE of Jaén, road to Gotas de Agua, in low hills W of the Río Marañón, 05°41'43" S, 78°4645" W, 742 m, disturbed dry thorn scrub, 17.V.2011 (fruit), Hughes 3139 (MICH; also MOL, MT, USM, fide C. E. Hughes, pers. comm.).Heteropterys tiinae is named in honor of the Finnish botanist Tiina Elina Särkinen (b. 1981), who collected the type in collaboration with A. Daza, H. Vandrot, and S. D'ugard; the species is known only from the two collections cited here. Heteropterys tiinae is probably referable to the complex of H. tomentosa A. Juss., with the geographically closest species being H. falcifera A. Juss. of Bolivia and H. fulva Cuatrec. of Peru. It is readily distinguished from those species, and from all other species of Heteropterys, by the following combination of characters: small nearly glabrous leaves with large glands on the abaxial surface (not on the margin), epipetiolar stipules, umbels or corymbs of 4-6 flowers, well-developed peduncles with the bracteoles borne near the base, and tiny greenish petals. The flowers are among the smallest in the family. The yellow-green color of the petals is probably unique in Heteropterys H.B.K. and highly unusual in the family. The only other species of Malpighiaceae with greenish petals that I recall is Pterandra guianensis W. R. Anderson (W. Anderson, 1981; C. Anderson, 1997).
Lophanthera marcelae W. R. Anderson, sp. nov.
Lophanthera marcelae differs from all other species of Lophanthera by its bicarpellate ovary.
Small shrub; stems densely and persistently sericeous with initially coppercolored hairs 0.5-1 mm long, straight, and sessile, belatedly glabrescent and developing circular or elliptical lenticels ca. 0.5 mm long. Lamina of larger leaves 13-18.4 cm long, 3-3.9 cm wide, narrowly elliptical, cuneate at base, flat or slightly revolute at margin, long-acuminate at apex with acumen 1.5-2 cm long, adaxially eglandular and glabrous at maturity, abaxially ± persistently sericeous with the hairs originally dense on midrib, numerous on laminar tissue but mostly not touching and not nearly dense enough to conceal epidermis, bearing 3-5 small impressed glands on each side of midrib between midrib and margin in proximal 2/3 of lamina, the principal lateral veins 12-15 or more on each side of midrib, fine and only moderately prominent on both sides of dried lamina, difficult to distinguish from lesser lateral veins; petiole 8-10 mm long, densely and ± persistently sericeous, eglandular (but sometimes glands on decurrent base of lamina seeming to be on apex of petiole); stipules intrapetiolar (borne on adaxial face of petiole at base), 4-6 mm long, narrowly triangular and tapering gradually to apex, membranous, the pair 1/2-2/3 connate, abaxially sericeous, adaxially glabrous, persistent. Inflorescence terminal and erect, 10 cm long (incomplete), bearing 25 cincinni, the cincinni proximally decussate, distally in no consistent order, the axis densely and persistently sericeous like stems; cincinni ascending, each containing 1-2 flowers; bracts 2-3.5 mm long, 0.7-1 mm wide, narrowly triangular, abaxially sericeous, adaxially glabrous, eglandular; primary peduncle (from bract to first joint) 7-9 mm long, loosely sericeous, bearing slightly below its apex 2 bracteoles like the bract but shorter, 1 usually eglandular, the other (occasionally both) with a spatulate apical extension terminating in a discoid gland; pedicel (in bud) 7-8 mm long, loosely sericeous. Sepals (in bud) 2-3 mm long, ca. 2 mm wide, ovate or elliptical, rounded and erose or denticulate at apex, glabrous or bearing a few abaxial hairs, all eglandular or 2 adjacent sepals bearing 1 gland each, the 2 glands adjacent to each other, distally free from the sepals and slightly reflexed. Petals pink, glabrous, nearly entire (minutely denticulate), dorsally smooth. Stamens 10, all fertile, glabrous; filaments distinct or slightly connate only at very base; anthers (in bud) 2-2.5 mm long, ± alike, the connective shorter than locules at apex and swollen at base, the outer locules bearing a very narrow (0.10-0.15 mm wide) membranous longitudinal wing extending from apex at least 2/3 of the distance to base. Receptacle glabrous between androecium and gynoecium. Gynoecium glabrous, bicarpellate; ovary with both locules fertile; styles 2, long and very slender, subulate, terminating in minute apical stigmas, bent in bud. Fruit unknown.
Type: COLOMBIA. Chocó: Región del (Bajo) Río Baudó, 6.II.1967 (immature flower), H. P. Fuchs, L. Zanella & J. H. Torres R. 21771 (holotype: COL!).
Lophanthera marcelae is named in honor of Marcela Celis, a Colombian student of Iridaceae, who was wonderfully helpful to me and my wife, Christiane, when we visited Colombia in 2009 to work on the Malpighiaceae for the Catálogo de las Plantas de Colombia.
Unfortunately, the holotype of Lophanthera marcelae (the single specimen known) bears only flowers in bud, but there can be no doubt that this is a distinctive new species. It is to be compared to the two other species of Lophanthera that have pink, dorsally smooth petals, L. hammelii W. R. Anderson and L. spruceana Nied.; the other three species of the genus have yellow, dorsally carinate petals (Anderson, 1983). The following key distinguishes L. marcelae from the other pink-flowered species.
| 1 Gynoecium densely sericeous; bracteole gland, when present, sessile and covering much of the abaxial surface of the bracteole; stipules 1.5-3 mm long, connate their whole length; northwestern Amazonia (Brazil and Colombia) .................... L. spruceana | |
| 1 Gynoecium glabrous; bracteole gland, when present, terminal and peltate; stipules 3-6 mm long, at least partially connate but often distally distinct. | |
| ]]> 2 Gynoecium tricarpellate; calyx bearing 10 abaxial glands; anther wings 0.2-0.4 mm wide, widest distally; Costa Rica ....................................................... L. hammelii | |
| 2 Gynoecium bicarpellate; calyx bearing 0-2 abaxial glands; anther wings 0.1-0.15 mm wide and of uniform width; Colombia (Chocó)...................................... L. marcelae | |
As the above key implies, it seems likely that L. marcelae is most closely related to L. hammelii; it is worth noting that they are the only trans-Andean species of Lophanthera A. Juss., the other four all being Amazonian (Anderson, 1983). When L. marcelae is better known more differences between them will probably emerge. For example, L. hammelii is a large shrub or tree with a very long, pendent inflorescence, while the type of L. marcelae was a small shrub with a short, apparently erect inflorescence, but without more material it is difficult to know how much significance to attach to that difference. When we have mature flowers and fruits of L. marcelae, they will probably differ in additional characters from those of L. hammelii. Lophanthera marcelae is the only species of the genus to have fewer than three carpels in the gynoecium. Its very narrow anther wings are noteworthy, but it is possible that they enlarge somewhat in anthesis.
It has long been evident from their morphology that Lophanthera and Spachea A. Juss. are similar genera, especially in their gland-bearing bracteoles and dry schizocarpic fruits (Anderson, 1981). Molecular studies (e.g., Davis et al., 2001) have strongly supported their sister status. In most characters Spachea is derived relative to Lophanthera (Anderson et al., website, consulted in July 2013): In Spachea there are glands on both sides of the lamina, whereas in Lophanthera they are only on the abaxial side as in most Malpighiaceae. In Spachea the styles are stout and the stigmas and pollen grains are large, while Lophanthera has subulate styles and tiny stigmas and pollen grains like most byrsonimoid genera. In Spachea the species are functionally dioecious, whereas in Lophanthera all the plants bear functionally bisexual flowers, as in most Malpighiaceae. In one character, however, Lophanthera is derived and Spachea has the presumably ancestral condition — in Lophanthera the outer locules of the anthers bear longitudinal wings, while those of Spachea are smooth as in most Malpighiaceae. I have argued (website, consulted July 2013) that this character justifies the continued recognition of both genera, because if Spachea is considered to be nested within Lophanthera, one has to assume that its smooth anther locules represent a reversal to an ancestral condition. Davis and Anderson (2010) have shown in their new phylogeny that L. hammelii is more closely related to Spachea than to the other species of Lophanthera sampled. This is one of the very few cases in which our molecular results have contradicted our morphological data. I feel that we should continue to recognize Lophanthera, at least until this conflict has received further study. In this context it is intriguing that the anther wings in L. marcelae are so very narrow. They suggest that possibly the anthers of Spachea really are derived from an ancestor that lost the prominent wings found on the anthers of other species of Lophanthera. Thus L. marcelae is a species of considerable phylogenetic interest and every effort should be made to collect good material of it, both to expand our very imperfect knowledge of its morphology and to add sequences from its DNA to our phylogenetic tree.
Mascagnia corymbosa W. R. Anderson, sp. nov. (Fig. 5).

Mascagnia corymbosa differs from M. ovatifolia in its smaller, eglandular laminas and unbranched inflorescences borne in the axils of a mature leaves.
Slender woody vine; stems tightly sericeous to glabrate. Lamina of leaves 6.7-9.3 cm long, 3-5.3 cm wide, ovate or elliptical, cuneate at base, abruptly short-acuminate at apex, nearly glabrous on both sides at maturity but with scattered loosely appressed hairs, especially on midrib, eglandular, the principal lateral veins 5-7 on each side of midrib, the reticulum visible below but not prominent on either side; petiole 8-15 mm long, slender, thinly and tightly sericeous, eglandular; stipules ca. 0.3 mm long and wide, triangular, sericeous, borne on stem at base of petiole, persistent or deciduous. Inflorescence unbranched, borne in axil of full-sized leaf, the stalk ca. 3 cm long, slender, sericeous or glabrescent, distally flattened, bearing a tight corymb 4-6 mm long of 10 or more flowers; bracts 0.9-1.5 mm long, ca. 0.4 mm wide, narrowly triangular, eglandular, pilose, persistent; peduncle (in fruit) 6-10 mm long, slender, loosely sericeous or (especially distally) glabrescent; bracteoles borne at apex of peduncle or 1 slightly below, 0.4-0.5 mm long, ca. 0.2 mm wide, narrowly triangular, eglandular, pilose, persistent; pedicel (in fruit) 11-12 mm long, slender, proximally glabrous, distally sericeous to glabrescent. Flowers not seen, the only known collection bearing mature fruits; 4 sepals abaxially biglandular, the fifth eglandular; styles truncate at apex. Samara orbicular, 2.8-3.3 cm in diameter; lateral wing membranous, continuous at base and apex, not cleft at apex but broadly and shallowly emarginate, nearly glabrate at maturity; dorsal winglet 8-10 mm wide, prolonged upward toward apex of samara and confluent with lateral wing; ventral crest 1-1.5 mm wide, extending from apex of nut upward and confluent with lateral wing; nut inserted above center of lateral wing, persistently thinly sericeous; ventral areole ovate or triangular, 3.5-4 mm high, 2.3-3 mm wide; fruit subtended by a short, obscure and not darkly colored, sparsely pilose 3-lobed disc.
Type: COLOMBIA. Antioquia: Mun. Mutatá, carretera de Bajirá hacia Caucheras, 2 km; bosque primario perturbado, 40 m, 20.III.1983 (fruit), J. Brand & E. Ascanio 315 (holotype: HUA!; isotype: MO!).
]]> This interesting species is named for its unbranched, tightly corymbose inflorescence axillary to a full-sized leaf. Among the species of Mascagnia that have such unbranched axillary inflorescences M. corymbosa is distinguished by the combination of nearly glabrate leaves with both the lamina and the petiole eglandular, very congested clusters of flowers, long slender peduncles and pedicels that are sericeous to glabrescent, eglandular bracteoles, the orbicular samara that is only broadly and shallowly emarginate at the apex, and the pilose disc below the fruit. The other Colombian species of Mascagnia that bear their flowers in unbranched inflorescences axillary to full-sized leaves have the lamina more or less densely and persistently hairy at maturity, at least abaxially. The only other Colombian species with a pilose disc is M. ovatifolia (H.B.K.) Griseb., but in that species the inflorescence is paniculate with the ultimate flower clusters less congested, the leaves are larger with the lamina bearing a pair of glands at the abaxial base, and the disc below the fruit is large and dark red. Mascagnia corymbosa is known only from the type, which is in fruit. I cannot be certain of the original shape of the styles, but some give the impression that they were lyrate. If that is the case, the petals will probably prove to be pink or lilac; I know of no yellow-flowered species of Mascagnia with lyrate posterior styles.
ACKNOWLEDGMENTS
Karin Douthit drew the beautiful illustrations. Tiina Särkinen and Colin E. Hughes kindly sent specimens of Heteropterys tiinae and provided additional information. I am most grateful for the hospitality shown by Colombian botanists during visits to COL and HUA in 2009; Julio Betancur B., Felipe Cardona, Marcela Celis, Herberto A. David, Carlos A. Parra-O., and Lauren Raz were particularly helpful. I thank Christiane Anderson for her comments. Thomas F. Daniel, Emily Lott, and Victor Steinmann reviewed the manuscript and contributed helpful suggestions and corrections. The curators of the following herbaria generously provided gifts and loans of specimens, and/or access to their collections: CAS, CHAPA, COL, DS, ENCB, F, GH, GUADA, HUA, IBUG, LL, MEXU, MICH, MO, NY, TEX, UC, US, WIS, XAL. My research was supported by NSF grant DEB-0543909 to the University of Michigan and an American Philosophical Society Franklin Research Grant.
LITERATURE CITED
Anderson, C. 1997. Revision of Pterandra (Malpighiaceae). Contr. Univ. Michigan Herb. 21: 1-27. [ Links ]
Anderson, W. R. 1981. Malpighiaceae. The botany of the Guayana Highland. Part XI, Malpighiaceae. Mem. New York Bot. Gard. 32: 21-305. [ Links ]
]]>Anderson, W. R. 1983. Lophanthera, a genus of Malpighiaceae new to Central America. Brittonia 35: 37-41. [ Links ]
Anderson, W. R., C. Anderson and C. C. Davis. 2006-. Malpighiaceae. http://herbarium.lsa.umich.edu/malpigh/index.html [accessed 2013] [ Links ]
Davis, C. C. and W. R. Anderson. 2010. A complete generic phylogeny of Malpighiaceae inferred from nucleotide sequence data and morphology. Amer. J. Bot. 97: 2031-2048. [ Links ]
Davis, C. C., W. R. Anderson and M. J. Donoghue. 2001. Phylogeny of Malpighiaceae: Evidence from chloroplast ndhF and trnl-F nucleotide sequences. Amer. J. Bot. 88: 1830-1846. [ Links ]
Gates, B. 1982. Banisteriopsis, Diplopterys (Malpighiaceae). Flora Neotropica Monograph 30: 1-237. [ Links ]
]]>Niedenzu, F. 1928. Malpighiaceae. In: Engler, A. (ed.). Das Pflanzenreich IV. 141: 1-870. [ Links ]
]]>