
0.4) were excluded from the analyses. This relative stability suggested that CR was regulated by a physiological feedback such as the fraction of poorly digestible organic matter.]]>
0.4) se excluyeron de los análisis. La estabilidad relativa de TA permite sugerir que esta variable está determinada por una retroalimentación fisiológica, que en el caso particular de este experimento estuvo determinada por el contenido de materia orgánica de baja digestibilidad de la dieta.]]>
Tasa de aclaramiento del mejillón Mytilus galloprovincialis. II. Respuesta a variables no correlacionadas del seston (cantidad, calidad y contenido de clorofila)*
Clearance rate of the mussel Mytilus galloprovincialis. II. Response to uncorrelated seston variables (quantity, quality, and chlorophyll content)**
R Filgueira, MJ Fernández–Reiriz*, U Labarta
Consejo Superior de Investigaciones Científicas (CSIC), Instituto de Investigaciones Marinas, C/ Eduardo Cabello 6, 36208 Vigo, Spain. * E–mail: mjreiriz@iim.csic.es
]]> Recibido en septiembre de 2009
ABSTRACT
Clearance rate (CR) response of Mytilus galloprovincialis was determined over 32 experiments in which the trophic conditions of its natural habitat, the Galician rías (Spain), were reproduced. Experimental diets were prepared by mixing pulverized sediment, Tahitian Isochrysis aff. galbana, and pulverized Spartina sp., avoiding multicollinearity among the seston variables used to characterize the diets: total particulate matter (TPM), quality (f = particulate organic matter (POM)/TPM), and chlorophyll content. This approach allowed us to identify the seston variables that have the most important effect on CR, as well as develop a mathematical model of CR response to the environmental trophic characteristics, without worrying about the statistical problems associated with multicollinearity among explanatory variables. Each of the three seston variables had a significant effect on CR response. Maximum CR was observed at a seston concentration of 1.2 mg L–1. The CR was inversely correlated with the fraction of low digestible organic matter (fdet = POMSpartina sp./TPM), and was relatively consistent when poorly digestible diets (fdet > 0.4) were excluded from the analyses. This relative stability suggested that CR was regulated by a physiological feedback such as the fraction of poorly digestible organic matter.
Key words: Mytilus galloprovincialis, clearance rate, chlorophyll, seston quantity and quality.
RESUMEN
Se realizaron 32 experimentos en los que se determinó la tasa de aclaramiento (TA) en Mytilus galloprovincialis ante una matriz de dietas que simulaba el rango de condiciones tróficas de su hábitat natural, las rías gallegas (España). Las dietas experimentales se prepararon empleando sedimento pulverizado, Isochrysis aff. galbana (variedad tahitiana) y Spartina sp. pulverizada, que permitieron la preparación de las dietas evitando la colinealidad entre las variables del seston empleadas para su caracterización: material particulado total (MPT), calidad (f = materia orgánica particulada (MOP)/MPT) y contenido de clorofila. Esta aproximación permitió determinar las variables del seston que afectan en mayor medida a la TA y establecer su patrón de respuesta en función de las características tróficas del medio evitando problemas estadísticos asociados con la multicolinealidad de las variables explicativas. Los resultados pusieron de manifiesto que las tres variables explicativas ejercieron un efecto significativo en la regulación de TA. Los valores máximos de aclaramiento se observaron con una concentración de seston en torno a 1.2 mg L–1. Se observó una reducción de la TA relacionada con el incremento de la proporción de materia orgánica de baja digestibilidad (fdet = MOPSpartina sp/MPT); sin embargo, la TA se mostró relativamente estable cuando las dietas de baja digestibilidad (fdet > 0.4) se excluyeron de los análisis. La estabilidad relativa de TA permite sugerir que esta variable está determinada por una retroalimentación fisiológica, que en el caso particular de este experimento estuvo determinada por el contenido de materia orgánica de baja digestibilidad de la dieta.
Palabras clave: Mytilus galloprovincialis, tasa de aclaramiento, clorofila, cantidad y calidad del seston.
]]>INTRODUCCIÓN
Los modelos de sistemas ecológicos representan herramientas poderosas en la toma de decisiones ya que simulan la organización, la función y los cambios del ecosistema. Un aspecto importante en el desarrollo de modelos para el cultivo de mejillones es la parametrización de la captación de energía en diferentes condiciones tróficas. La captación de energía no depende exclusivamente de las tasas de filtración y retención de partículas de alimento en las branquias, estimada mediante la tasa de aclaramiento (TA). Bajo altas concentraciones de seston, la selección preingestiva y la producción de pseudoheces son mecanismos que regulan la ingestión de alimento (Bayne et al. 1993, Ward y Shumway 2004); sin embargo, a bajas concentraciones de seston, como sucede en las rías de Galicia (materia particulada total < 3 mg L–1) donde el fitoplancton constituye la mayor proporción de seston (Figueiras et al. 2002), los procesos selectivos podrían ser de menor importancia, por lo que en estos sistemas la captación de energía dependería esencialmente de la TA.
La regulación de TA en Mytilus sp. expuesto a diferentes dietas ha sido estudiada ampliamente tanto bajo condiciones controladas de laboratorio como in situ (ver revisión por Bayne et al. 1993). En los estudios de laboratorio, intervalos predefinidos de variables ambientales permiten la determinación de ecuaciones matemáticas que describen la respuesta fisiológica. A pesar de ser un área de investigación muy productiva, la información sobre el efecto de diferentes variables del seston sobre la TA aún no es concluyente debido a deficiencias ya sea metodológicas o experimentales. La deficiencia más importante en los estudios que han utilizado la regresión múltiple para analizar la respuesta de TA a diferentes variables del seston es que éstos no consideran la multicolinealidad que ocurre cuando algunas de las variables explicativas están muy correlacionadas entre sí, provocando un impacto muy negativo sobre la calidad y la estabilidad del modelo de regresión. Cuando se presenta la multicolinealidad, las conclusiones del modelo de regresión en cuanto al efecto significativo de las variables correlacionadas en la variable de respuesta pueden ser espurias (Neter y Wasserman 1974) ya que no se puede distinguir entre el efecto de una variable explicativa y el efecto de la variable correlacionada. La manera más simple de detectar la multicolinealidad es realizar análisis de correlación entre pares de variables explicativas; si dos de estas variables están altamente correlacionadas, sólo una debería de usarse para explicar la variación en la variable de respuesta, ya que en esencia una transmite toda la información de la otra. A pesar de la importancia de considerar la multicolinealidad al determinar la significancia estadística, generalmente no se toman en cuenta las relaciones entre las variables explicativas en el diseño experimental. Después de una revisión exhaustiva de la literatura, consideramos que el presente trabajo es el primero en hacerlo.
En el presente análisis se estudió la TA del mejillón Mytilus galloprovincialis bajo un espectro de dietas que simulaba las condiciones tróficas de su hábitat natural (las rías gallegas). Las dietas experimentales se prepararon mezclando sedimento pulverizado, Isochrysis galbana y una matriz orgánica compuesta de Spartina sp. pulverizada. El uso de estos componentes permitió diseñar las dietas a priori para evitar la correlación entre las siguientes variables que caracterizan la dieta: material particulado total (MPT), calidad (f = material orgánico particulado (MOP)/MPT) y contenido de clorofila (Chla). Cada una de estas variables juega un papel importante en el desarrollo de los mejillones (Freites et al. 2003) y son empleadas frecuentemente en modelos ecofisiológicos (ver tabla 1). Este enfoque permitió (1) la identificación de las variables del seston que afectan la TA en mayor medida y (2) una descripción matemática de la respuesta de TA según las características tróficas.
MATERIAL Y MÉTODOS
Recolección y aclimatación de mejillones
Se recolectaron ejemplares de M. galloprovincialis (Lamarck 1819) de una batea de cultivo en la Ría de Arousa (Galicia, NO de España), y se seleccionaron individuos de 59 ± 2.5 mm de longitud a los que se retiraron los epibiontes. Los mejillones se mantuvieron según el procedimiento descrito por Filgueira et al. (2009).
Dietas experimentales
]]> Las dietas se prepararon mezclando proporciones variables de sedimento pulverizado, Isochrysis aff. galbana (variedad tahitiana) y Spartina sp. pulverizada y previamente liofilizada (tabla 2). I. galbana se cultivó en tanques de 50 L en condiciones controladas, el sedimento se recolectó del lecho marino debajo de las bateas, y Spartina se recolectó a lo largo de la costa cortando el tallo de plantas vivas. El detritus de los dos últimos componentes se secó, pulverizó y tamizó con una malla de nilón de 50 µm de luz. El agua de mar en el tanque de almacenamiento fue filtrada a través de un sistema de cartucho (CUNO Super Micro–Wynd de 10, 5 y 1 µm; CUNO Betapure de 0.5 µm), con un tamaño de poro efectivo de 0.5 µm, y tratada con luz UV. Al inicio del experimento las dietas se prepararon mezclando los diferentes componentes en un tanque con aireación intensa para mantener la homogeneidad y evitar la sedimentación de partículas.Las mediciones de MPT, MOP y Chla se tomaron por triplicado para cada dieta siguiendo el procedimiento descrito por Filgueira et al. (2009). Las características (Chla, MPT y f) de las 32 dietas (tabla 2) cubrieron el intervalo típico observado en las rías gallegas. Las condiciones tróficas de estas rías y las usadas en este estudio se muestran en la tabla 3. Los valores de MPT, MOP y f de las rías son similares a los aquí analizados, aunque MPT puede ser mayor durante tormentas esporádicas. El intervalo de clorofila analizado cubrió 89.4% de los valores observados durante un estudio de nueve años (833 muestras, PROVIGO 1996). En el presente trabajo no se consideraron las concentraciones de clorofila excepcionalmente altas que ocurren durante los florecimientos fitoplanctónicos.
Medición y estandarización de la tasa de aclaramiento
Para estimar TA, para cada dieta se usaron diez cámaras individuales con mejillones y una cámara control sin ellos. En cada tratamiento se usaron diferentes mejillones. La TA se estimó con el método de cámara de flujo continuo siguiendo la metodología descrita por Filgueira et al. (2006). Los experimentos se realizaron durante 4 h, realizando las mediciones de TA después de 2, 3 y 4 h. Los mejillones que desovaron durante los experimentos fueron descartados de los análisis.
En todos los casos la TA se estandarizó a una longitud de mejillón de 60 mm mediante la siguiente fórmula:
donde TAstd es la tasa de aclaramiento estandarizada, TAexp es la tasa de aclaramiento experimental, Lstd es la longitud de estandarización, Lexp es la longitud del individuo experimental y b es el exponente que relaciona la TA con la longitud. Se utilizó un valor de 1.72 como exponente de estandarización respecto a la longitud (Filgueira et al. 2008).
Protocolo para seleccionar el mejor modelo de regresión: TA vs MPT, f, Chla
Se consideraron MPT, f, Chla, su combinación lineal, términos cuadráticos y términos de interacción de primer orden como variables independientes potenciales en el modelo de regresión que mejor explicó la variación de TA. La estimación y selección del modelo de regresión se realizó siguiendo el siguiente protocolo. Primero se calculó la varianza explicada por cada variable independiente y todas las posibles combinaciones de las variables independientes mediante regresión múltiple (Best Subsets Procedure en Statistica 6.0). Después se seleccionaron como mejores modelos potenciales aquellos modelos de regresión que (1) explicaban gran parte de la varianza y (2) presentaban parámetros estadísticamente significativos al aplicar una estimación no lineal (procedimiento de mínimos cuadrados y método de Levenberg–Marquardt en Statistica 6.0). Por último, se utilizó el critero de información de Akaike (AIC; Motulsky y Christopoulos 2004) para seleccionar el modelo de regresión más adecuado de entre los modelos potenciales seleccionados. Se utilizó este enfoque para calcular un coeficiente de comparación (AIC) para cada regresión considerando el número de casos, los parámetros y la suma de cuadrados de los residuales de regresión.
]]> RESULTADOS
Dietas experimentales
En la tabla 2 se muestran las características de las 32 dietas experimentales. El coeficiente de correlación de Pearson (r) no presentó una correlación estadísticamente significativa entre las variables (rChla x MPT = –0.288, P = 0.110; rChla X f = 0.252, P = 0.163; rMPT x f = –0.235, P = 0.196). Además, los factores de inflación de la varianza mostraron valores cercanos a uno (VIFChla = 1.13, VIFMPT = 1.12, VIFf = 1.10), lo que indica que la multicolinealidad fue insignificante.
Modelos de regresión: TA vs MPT, f, Chla
El modelo de regresión no lineal que mejor explicó los resultados experimentales incluyó los términos lineales de f, MPT y Chla, así como el término cuadrático de MPT y la interacción entre f y Chla. La superficie de respuesta se puede describir con la siguiente ecuación:
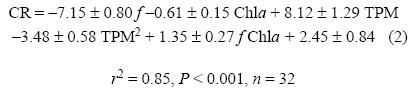
Se aplicó el mismo protocolo estadístico a los análisis de pares entre MPT y Chla, MPT y f, y Chla y f, los cuales son casos particulares del anterior, pero aportan nuevas pistas en cuanto al papel de las variables independientes y permiten una visualización tridimensional de los modelos de regresión.
En la figura 1 se muestra la respuesta de TA en función del contenido orgánico o calidad del seston (f) y MPT. El modelo de regresión no lineal que mejor explicó los resultados se describe con la ecuación:

Al igual que en la ecuación 2, el término cuadrático indicó un MPT óptimo que maximiza TA; sin embargo, el término de interacción (MPT x f) sugirió que el valor óptimo depende de f. La relación entre el MPT óptimo y f que maximiza TA se estimó mediante cálculo diferencial:
]]>
En la figura 2 se muestra la variación de TA en función tanto de MPT como de Chla. La regresión no lineal que mejor explicó los resultados dependió exclusivamente de MPT. La superficie de respuesta puede describirse con la siguiente ecuación:

De acuerdo con esta ecuación, los valores máximos de TA se obtuvieron con 1.15 ± 0.40 mg MPT L–1.
En la figura 3 se muestra la variación de TA en función de Chla y f. La regresión no lineal que mejor explicó los resultados se describe con la siguiente ecuación:

La TA decreció marcadamente al incrementarse el valor de f cuando Chla era baja (fig. 3). Esto evidencia la posible existencia de un umbral de clorofila que determina el comportamiento de los mejillones. Para M. edulis se ha establecido este umbral entre 0.5 y 1.0 µg Chla L–1 (Dolmer 2000, Pascoe et al. 2009). De acuerdo con los resultados del presente trabajo, para M. galloprovincialis este umbral podría establecerse entre 1.18 y 1.75 µg Chla L–1.
Este umbral sugirió la posibilidad de segregar las dietas en dos grupos según la Chla. Tal segregación mostró un cambio en la regresión que mejor explicaba TA cuando MPT, f y Chla fueron considerados simultáneamente (ec. 2). Para las dietas con Chla por encima del umbral, la mejor regresión dependió sólo de MPT. La superficie de respuesta está dada por:

La relación cuadrática entre TA y MPT mostró que los valores máximos de TA se obtuvieron con un MPT óptimo de 1.26 ± 0.40 mg L–1.
]]> Para las dietas con Chla por debajo del umbral, la regresión que mejor explicó la variación de TA fue más simple que la regresión que incluyó todas las dietas simultáneamente (ec. 2). De hecho, los términos de clorofila no fueron significativos en la nueva regresión:
La relación cuadrática entre TA y MPT mostró un MPT óptimo de 1.26 ± 0.59 mg L–1 que maximizó la TA.
El análisis de pares identificó un cambio fuerte en la respuesta de TA, dependiente de Chla, lo cual ha sido corroborado en otros estudios (Dolmer 2000, Pascoe et al. 2009). La segregación de las dietas experimentales según el umbral de clorofila (1.18–1.75 µg Chla L–1) mejoró el ajuste de los modelos de regresión incrementando la varianza explicada. Primero, las dietas con Chla por encima del umbral provocaron una respuesta consistente de TA (3.2–4.3 L h–1), lo que podría explicarse en términos de MPT (ec. 7), con valores máximos de TA a 1.26 mg MPT L–1. Segundo, las dietas con Chla por debajo del umbral provocaron respuestas muy diferentes en TA (de 1.0 a 5.0 L h–1), lo que podría explicarse en términos de MPT y la calidad de la dieta, con una relación inversa entre TA y f, y valores máximos de TA a 1.25 mg MPT L–1 (ec. 8). Por tanto, aunque Chla no esté explícita en los modelos de regresión, si está involucrada en la regulación de TA como una variable umbral importante.
DISCUSIÓN
A pesar de la cuantiosa literatura alusiva a la respuesta de TA en los mejillones, la información sobre el efecto de diferentes variables del seston en su regulación aún no es concluyente. Una falla importante en investigaciones previas ha sido no considerar la multicolinealidad. Las conclusiones de los experimentos que presentan multicolinealidad son ambiguos y los efectos de las variables explicativas en la respuesta de TA muy probablemente son espurios. Aquí el principal objetivo metodológico fue diseñar un conjunto de dietas para evitar la multicolinealidad entre las variables explicativas consideradas (MPT, f y Chla). El análisis mostró que estas variables no estuvieron correlacionadas y los factores de inflación de la varianza indicaron que la multicolinealidad era insignificante. Después de una extensa revisión de la literatura, creemos que este estudio es el primero en considerar la multicolinealidad en el diseño experimental, fortaleciendo así las conclusiones.
Varios estudios han establecido que los bivalvos regulan su TA para optimizar la asimilación de energía (Bayne et al. 1993, Velasco y Navarro 2002). En el presente trabajo se analizó esta respuesta fisiológica mediante relaciones funcionales entre TA y diferentes variables ambientales. La relación cuadrática observada entre TA y MPT (ecs. 7, 8) indica que los valores máximos de TA se presentan a concentraciones de seston de 1.25 mg L–1. Kiørboe et al. (1981) observaron un patrón similar en M. edulis, aunque no es posible hacer una comparación de las concentraciones de MPT asociadas con los valores máximos de TA ya que la composición dietética y los métodos de caracterización empleados fueron diferentes. Otros estudios han encontrado un amplio espectro de relaciones entre TA y MPT en M. edulis, incluyendo relaciones inversas (Kiørboe et al. 1980, Clausen y Riisgard 1996) y directas (Newell y Shumway 1993, Hawkins et al. 1996). La disparidad entre estudios puede estar relacionada con la concentración de seston o la correlación entre las variables explicativas.
Nuestros resultados sugieren que Chla podría considerarse una variable umbral que cambia la respuesta de TA dependiendo de si la concentración en la dieta está por encima o por debajo del umbral. Las dietas con Chla >1.18–1.75 µg L–1 provocan una respuesta consistente de TA que depende exclusivamente de MPT (ec. 7). Por debajo de este umbral se observa una respuesta de TA muy diferente (variando de 1.0 a 5.0 L h–1) que podría ser explicada por MPT y la calidad en contenido orgánico de la dieta (ec. 8). En otros estudios se ha notado la respuesta de TA a dietas con baja Chla. Varios autores observaron una reducción total de la actividad de filtración en M. edulis a 0.5–1 | g Chla L–1 (ver revisión de Dolmer 2000). Hawkins et al. (1999) y James et al. (2001), trabajando con Perna canaliculus, y Hawkins et al. (2001), con Chlamys farreri, observaron una reducción en la actividad de filtración, pero sin cesarla. Pascoe et al. (2009) observaron en M. edulis una reducción tanto en TA como en el número de animales alimentándose, reducción que también fue observada recientemente por Strohmeier et al. (2009) en M. edulis y Pecten maximus. Filgueira et al. (2009), en un experimento de 9 h, observaron una TA constante y alta en M. galloprovincialis durante las primeras 3 h. Estos resultados difieren del presente trabajo en que aquí los mejillones que fueron alimentados con dietas con alto contenido orgánico presentaron una reducción significativa de TA (ec. 8) al principio del experimento. Se ha encontrado una relación inversa entre TA y f para M. edulis (Bayne et al. 1993) y M. chilensis (Velasco y Navarro 2002), aunque con dietas con mayor contenido de seston. En algunos casos el incremento de TA en respuesta a dietas con bajo contenido orgánico ha ido acompañado por la selección de partículas en los palpos labiales y, consecuentemente, la eficiencia en la ingestión de materia orgánica se ha incrementado (Bayne et al. 1993, Hawkins et al. 1996). Sin embargo, en este experimento no se observó la producción de pseudoheces.
La relación inversa entre la calidad en contenido orgánico de la dieta y la TA se ha interpretado como una respuesta fisiológica para mantener una tasa constante de ingestión orgánica durante periodos de baja concentración de seston total (Bayne et al. 1987). En este caso, la respuesta de TA a las dietas por debajo del umbral de clorofila de 1.18–1.75 µg L–1 no permite que los animales alcancen una tasa constante de ingestión orgánica (ec. 8), por lo que la cantidad de materia orgánica ingerida no parece estar involucrada en la regulación de TA. Por otro lado, Willows (1992) desarrolló un modelo teórico que supone que la TA es regulada por la capacidad del sistema digestivo. Por tanto, el balance entre el tiempo de residencia en el intestino y la tasa de ingestión podría ser un regulador clave de la TA. Con base en el modelo de Willows, Fernández–Reiriz et al. (2007) sugirieron que en ecosistemas con baja concentración de seston como las rías gallegas (MOP < 3 mg L–1), el volumen estomacal de los mejillones se mantiene por debajo de su capacidad máxima, estimulando valores consistentemente altos de TA; sin embargo, sus resultados y los aquí presentados no sustentan las suposiciones del modelo de Willows ya que se observaron valores bajos de TA en mejillones cuyas dietas contenían bajas concentraciones de seston, lo que indica que TA no está determinada exclusivamente por la capacidad estomacal.
]]> Una de las variables que podría ser importante en la regulación de TA es la digestibilidad de los componentes alimenticios. Según Hawkins et al. (1999), la regulación de TA en los mejillones podría estar relacionada con el contenido de materia orgánica fitoplanctónica en su dieta, lo que indica mayor digestibilidad de la materia orgánica rica en clorofila. Con base en esta hipótesis, se separó el contenido orgánico (f) en tres fracciones que reflejaran los diferentes componentes usados para preparar las dietas: fmicro (MOPmicroalgas/MPT), fsed (MOPsedimento/MPT) y fdet (MOPdetritus/MPT). Estos tres componentes de f junto con MPT y Chla fueron considerados variables explicativas y se aplicó el mismo protocolo descrito en la sección de material y métodos para seleccionar la mejor regresión, mostrando que TA depende exclusivamente de fdet:
Esta ecuación sugiere que los valores máximos de TA se presentan en ausencia de Spartina sp., y que TA se reduce significativamente cuando la materia orgánica de Spartina sp. aumenta en relación con la concentración de seston. Esta respuesta podría reflejar la baja digestibilidad de Spartina sp. en comparación con la otra fuente principal de materia orgánica, I. galbana. Aunque se ha demostrado que los organismos filtradores pueden asimilar materia orgánica vegetal, la presencia de componentes de baja digestibilidad (celulosa y lignina) dificulta este proceso (Duggins y Eckman 1997). Esta reducida digestibilidad podría estar relacionada con compuestos polifenólicos (Tugwell y Branch 1992), los cuales han mostrado tener un impacto negativo significativo en la TA de Crassotrea gigas y Mytilus trossulus (Levinton et al. 2002). Por tanto, la reducción en TA observada en este estudio al exponer los mejillones fueron a dietas con alta calidad en contenido orgánico (f) y baja Chla (debajo del umbral de 1.18–1.75 µg L–1) podría deberse a la baja digestibilidad del componente principal de estas dietas, Spartina sp. Ward et al. (1998) observaron que C. gigas alimentado con dietas mixtas presentó una TA significativamente menor con partículas de Spartina alterniflora que con células de Rhodomonas lens, y sugirieron que las diferencias probablemente se debían a la menor eficiencia de retención de las partículas de S. alterniflora más que a diferencias en la digestibilidad de cada fuente. No obstante, otros estudios han observado valores similares para Spartina y otros componentes dietéticos en bivalvos homorrábdicos (M. trossulus, Ward et al. 1998; Dreissena polymorpha, Bastviken et al. 1998) como M. galloprovincialis. Según Ward et al. (1998), M. trossulus tiene la capacidad de rechazar selectivamente el detritus de S. alterniflora sobre el de R. lens. En este proceso no se presentó regulación de TA y la selección se realizó en los palpos labiales, con la consecuente formación de pseudo–heces. Con las dietas experimentales usadas en el presente estudio los mejillones no produjeron pseudoheces, lo que indica la poca importancia de los procesos de selección en el balance energético en condiciones experimentales representativas de las rías gallegas.
La reducción de TA observada aquí para M. galloprovincialis dependiendo de su dieta podría interpretarse como un proceso de regulación fisiológica (Hawkins et al. 1996, Levinton et al. 2002, Pascoe et al. 2009). Tras la segregación del contenido orgánico en tres fracciones según su origen (sedimento pulverizado, I. galbana y Spartina sp.), se puede sugerir que la reducción de TA al alimentar los mejillones con dietas de alto contenido orgánico con Chla debajo del umbral (1.18–1.75 µg L–1) es producto de un alto porcentaje de materia orgánica de baja digestibilidad (fdet, ec. 9). Así, al excluir las dietas con fdet > 0.4 se observa una TA constante de 3.9 ± 0.42 L h–1 (coeficiente de variación de Pearson = 10.7%). Este valor es similar al encontrado por Filgueira et al. (2009) con tiempo y dietas experimentales similares y el mismo intervalo de clorofila, pero usando exclusivamente I. galbana y sedimento pulverizado. Este promedio también es similar a lo encontrado en los experimentos de campo realizados por Navarro et al. (1991), Iglesias et al. (1996), Babarro et al. (2000) y Petersen et al. (2008) con M. galloprovincialis en las rías de Galicia.
En conclusión, la respuesta de TA en M. galloprovincialis en condiciones experimentales representativas de las rías gallegas está relacionada sólo con la cantidad de partículas de baja digestibilidad en la dieta (ec. 9). La presencia de Spartina sp. causa una reducción significativa de la TA, mientras que la respuesta es relativamente consistente cuando se excluyen las dietas con fdet> 0.4 de los análisis (coeficiente de variación de Pearson = 10.7%). Los valores consistentes de TA sugieren que el umbral de clorofila de 1.18–1.75 µg L–1 es un artificio estadístico causado por las dietas con fdet> 0.4. Esto podría explicar lo observado por Fernández–Reiriz et al. (2007) en el sentido de que la regulación de TA no se debe a la disponibilidad de seston o a los niveles de clorofila, sino más bien demuestra una retroalimentación fisiológica que no depende de la capacidad del sistema digestivo. En el presente trabajo, esta retroalimentación podría atribuirse a un alto contenido de materia orgánica de baja digestibilidad en la dieta; sin embargo, se requieren ulteriores investigaciones para probar esta hipótesis, incluyendo estudios del balance digestivo y la eficiencia de absorción.
AGRADECIMIENTOS
Este estudio fue apoyado por PROINSA (proyecto CSIC 2004448), la Xunta de Galicia (PGDIT03RMA13E) y la UE (MABENE EVK3–2001–00144). El primer autor fue apoyado por CSIC–Xunta de Galicia y Caixanova con una beca para entrenamiento científico. Se agradece a L Nieto, B González y R Salgueiro (CSIC–España) su asistencia técnica en el laboratorio.
REFERENCIAS
]]>Babarro JMF, Fernández–Reiriz MJ, Labarta U. 2000. Feeding behaviour of seed mussel Mytilus galloprovincialis: Environmental parameters and seed origin. J. Shellfish Res. 19: 195–201. [ Links ]
Bastviken DTE, Caraco NF, Cole JJ. 1998. Experimental measurements of zebra mussel (Dreissena polymorpha) impacts on phytoplankton community composition. Freshwat. Biol. 39: 375–386. [ Links ]
Bayne BL, Hawkins AJS, Navarro E. 1987. Feeding and digestion by the mussel Mytilus edulis L (Bivalvia: Mollusca) in mixture of silt and algal cells at low concentrations. J. Exp. Mar. Biol. Ecol. 111: 1–22. [ Links ]
Bayne BL, Iglesias JIP, Hawkins AJS, Navarro E, Héral M, Deslous–Paoli JM. 1993. Feeding behaviour of the mussel Mytilus edulis: Responses to variations in quantity and organic content of the seston. J. Mar. Biol. Assoc. UK 73: 813–829. [ Links ]
Clausen IB, Riisgard HU. 1996. Growth, filtration and respiration in the mussel Mytilus edulis: No evidence for physiological regulation of the filter–pump to nutritional needs. Mar. Ecol. Prog. Ser. 141: 37–45. [ Links ]
]]>Dolmer P. 2000. Feeding activity of mussels Mytilus edulis related to near–bed currents and phytoplankton biomass. J. Sea Res. 44: 221–231. [ Links ]
Duggins DO, Eckman JE. 1997. Is kelp detritus a good food for suspension feeders? Effects of kelp species, age and secondary metabolites. Mar. Biol. 128: 489–495. [ Links ]
Fernández–Reiriz MJ, Duarte P, Labarta U. 2007. Modelos de comportamiento alimentario en el mejillón de las rías de Galicia. In: Figueras A (ed.), Biología y Cultivo del Mejillón (Mytilus galloprovincialis) en Galicia. Biblioteca de Ciencias (CSIC), Madrid, pp. 197–223. [ Links ]
Figueiras FG, Labarta U, Fernández–Reiriz MJ. 2002. Coastal upwelling, primary production and mussel growth in the Rías Baixas of Galicia. Hydrobiologia 484: 121–131. [ Links ]
Filgueira R, Labarta U, Fernández–Reiriz MJ. 2006. Flow–through chamber method for clearance rate measurements in bivalves. Design and validation of individual chambers and mesocosm. Limnol. Oceanogr. Methods 4: 284–292. [ Links ]
]]>Filgueira R, Labarta U, Fernández–Reiriz MJ. 2008. Effect of condition index on allometric relationships of clearance rate in Mytilus galloprovincialis Lamarck, 1819. Rev. Biol. Mar. Oceanogr. 43: 391–398. [ Links ]
Filgueira R, Fernández–Reiriz MJ, Labarta U. 2009. Clearance rate of the mussel Mytilus galloprovincialis. I. Response to extreme chlorophyll ranges. Cienc. Mar. 35: 405–417. [ Links ]
Freites L, Fernández–Reiriz MJ, Labarta U. 2003. Biochemical composition and energy content of the mussel Mytilus galloprovincialis of subtidal and rocky shore origin: Influence of environmental variables and source of mussel seed. Cienc. Mar. 29: 603–619. [ Links ]
Gardner JPA. 2002. Effects of seston variability on the clearance rate and absorption efficiency of the mussels Aulacomya maoriana, Mytilus galloprovincialis and Perna canaliculus from New Zealand. J. Exp. Mar. Biol. Ecol. 268: 83–101. [ Links ]
Hawkins AJS, Smith RFM, Bayne BL, Héral M. 1996. Novel observations underlying the fast growth of suspension–feeding shellfish in turbid environments: Mytilus edulis. Mar. Ecol. Prog. Ser. 131: 179–190. [ Links ]
]]>Hawkins AJS, James MR, Rickman RW, Hatton S, Weatherhead M. 1999. Modelling of suspension–feeding and growth in the green–lipped mussel Perna canaliculus exposed to natural and experimental variations of seston availability in the Marlborough Sounds, New Zealand. Mar. Ecol. Prog. Ser. 191: 217–232. [ Links ]
Hawkins AJS, Fang JG, Pascoe PL, Zhang JH, Zhang XL, Zhu MY. 2001. Modelling short–term responsive adjustments in particle clearance rate among bivalve suspension–feeders: Separate unimodal effects of seston volume and composition in the scallop Chlamys farreri. J. Exp. Mar. Biol. Ecol. 262: 61–73. [ Links ]
Iglesias JIP, Pérez–Camacho A, Navarro E, Labarta U, Beiras R, Hawkins AJS, Widdows J. 1996. Microgeographic variability in feeding, absorption, and condition of mussels (Mytilus galloprovincialis L): A transplant experiment. J. Shellfish Res. 15: 673–680. [ Links ]
James MR, Weatherhead MA, Ross AH. 2001. Size–specific clearance, excretion, and respiration rates, and phytoplankton selectivity for the mussel Perna canaliculus at low levels of natural food. NZ J. Mar. Freshwat. Res. 35: 73–86. [ Links ]
Kiørboe T, Møhlenberg F, Nøhr O. 1980. Feeding, particle selection and carbon absorption in Mytilus edulis in different mixtures of algae and resuspended bottom material. Ophelia 19: 193–205. [ Links ]
]]>Kiørboe T, Møhlenberg F, Nøhr O. 1981. Effect of suspended bottom material on growth and energetics in Mytilus edulis. Mar. Biol. 61: 283–288. [ Links ]
Levinton JS, Ward JE, Shumway SE. 2002. Feeding responses of the bivalves Crassostrea gigas and Mytilus trossulus to chemical composition of fresh and aged kelp detritus. Mar. Biol. 141: 367–376. [ Links ]
Motulsky H, Christopoulos A. 2004. Fitting Models to Biological Data Using Linear and Nonlinear Regression. Oxford Univ. Press, Oxford, 351 pp. [ Links ]
Navarro E, Iglesias JIP, Pérez–Camacho A, Labarta U, Beiras R. 1991. The physiological energetics of mussels (Mytilus galloprovincialis L) from different cultivation rafts in the Ría de Arosa (Galicia, NW Spain). Aquaculture 94: 197–212. [ Links ]
Neter J, Wasserman W. 1974. Applied Linear Statistical Models. Richard D Irwin, Homewood, Illinois, 842 pp. [ Links ]
]]>Newell CR, Shumway SE. 1993. Grazing of natural particulates by bivalve molluscs: Spatial and temporal perspective. In: Dame RD (ed.), Bivalve Filter Feeders in Estuarine and Coastal Ecosystem Processes. Springer–Verlag, Berlin, pp. 85–148. [ Links ]
Pascoe PL, Parry HE, Hawkins AJS. 2009. Observations on the measurement and interpretation of clearance rate variations in suspension–feeding bivalve shellfish. Aquat. Biol. doi: 10.3354/ab00123. [ Links ]
Petersen JK, Nielsen TG, van Duren L, Maar M. 2008. Depletion of plankton in a raft culture of Mytilus galloprovincialis in Ría de Vigo, NW Spain. I. Phytoplankton. Aquat. Biol. 4: 113–125. [ Links ]
PROVIGO. 1996. Nueve Años de Muestreo Hidrográfico en la Ría de Vigo (Enero 1987–Enero 1996). Fundación PROVIGO, Vigo, 47 pp. [ Links ]
Strohmeier T, Strand Ø, Cranford P. 2009. Clearance rates of the great scallop (Pecten maximus) and blue mussel (Mytilus edulis) at low natural seston concentrations. Mar. Biol. 156: 1781–1795. [ Links ]
]]>Tugwell S, Branch GM. 1992. Effects of herbivore gut surfactants on kelp polyphenol defenses. Ecology 73: 205–215. [ Links ]
Velasco LA, Navarro JM. 2002. Feeding physiology of infaunal (Mulinia edulis) and epifaunal (Mytilus chilensis) bivalves under a wide range of concentrations and qualities of seston. Mar. Ecol. Prog. Ser. 240: 143–155. [ Links ]
Velasco LA, Navarro JM. 2005. Feeding physiology of two bivalves under laboratory and field conditions in response to variable food concentrations. Mar. Ecol. Prog. Ser. 291: 115–124. [ Links ]
Ward JE, Shumway SE. 2004. Separating the grain from the chaff: Particle selection in suspension– and deposit–feeding bivalves. J. Exp. Mar. Biol. Ecol. 300: 83–130. [ Links ]
Ward JE, Levinton JS, Shumway SE, T Cucci. 1998. Particle sorting in bivalves: In vivo determination of the pallial organs of selection. Mar. Biol. 131: 283–292. [ Links ]
]]>Willows RI. 1992. Optimal digestive investment: A model for filter feeders experiencing variable diets. Limnol. Oceanogr. 37: 829–847. [ Links ]
* Traducido al español por Christine Harris.
** Versión bilingüe (Inglés–Español) en formato PDF
]]>