La testosterona inhibe las respuestas contráctiles del agonista adrenérgico α1, fenilefrina, asociadas con la liberación de calcio intracelular en la aorta de rata
Testosterone inhibits the contractile responses to phenilephrine associated with the release of intracellular calcium in rat aorta
Carlos Castillo,* Enrique F. Castillo, Jorge López y Ruth M. López
Sección de Estudios de Posgrado e Investigación de la Escuela Superior de Medicina, Instituto Politécnico Nacional. México, D.F., México
]]> Recibido en su versión modificada: 07 de junio del 2005
* Correspondencia y solicitud de sobretiros:
Dr. Carlos Castillo.
Escuela Superior de Medicina del Instituto Politécnico Nacional, Plan de San Luis y Díaz Mirón, Col. Casco de Santo Tomás, CP 11340, México, D.F., México.
Tel. +52 (55) 57296260, Fax +52 (55) 53412726,
correo electrónico: drcarloscastillo@esmas.com
Resumen
]]> Utilizando segmentos de aorta de rata sin endotelio inmersos en solución sin Ca2+, evaluamos la capacidad de la testosterona para modificar el efecto contráctil del agonista adrenérgico fenilefrina, así como el incremento en el tono de reposo (ITR) asociado con la entrada capacitativa de calcio por el sarcoplasma. La testosterona [10-5—10-4 M] inhibió significativamente la contracción activada por la fenilefrina [10–6 M] y el ITR. Estos efectos no fueron modificados con cicloheximida [10–5 M] (inhibidor de la síntesis protéica), flutamida [10–5 M] (antagonista de receptores androgénicos), o aminoglutetimida [10-5 M] (inhibidor de la citocromo P450 aromatasa). La testosterona también inhibió las respuestas contráctiles de la serotonina [10–5 M], pero no de la cafeína [10–2 M]. Además, la testosterona inhibió las contracciones del ácido ciclopiazónico [10-6 M] y de la ryanodina [10–5 M] asociadas con el ingreso capacitativo de Ca2+ mediante canales de Ca2+ tipo no L. Estos datos sugieren que la testosterona interfiere con la vía de transducción de los receptores acoplados a proteínas Gq–11, e inhibe la entrada capacitativa de Ca2+ a través de canales de Ca2+ tipo L y tipo no L; los efectos son no genómicos, independientes de receptores androgénicos, y de la conversión testosterona en estrógenos.Palabras clave: Testosterona, liberación de Ca2+ intracelular, agonistas adrenérgicos α1
Summary
Using endothelium–denuded rat aortic rings incubated in Ca2+–free solution, we assessed the ability of testosterone to influence the contractile effect of phenylephrine, and the increase in resting tone (IRT) associated with Ca2+ ability to cross the plasma membrane. The addition of testosterone[10-5—10-4M]5 min before phenylephrine [10-6 M], inhibited both phenylephrine–induced contraction and IRT. These changes were not affected by cycloheximide (10-5 M; a protein synthesis inhibitor of), flutamide (10-5 M; an androgenic receptor antagonist), or by adding aminoglutethimide (10-5 M; an aromatase inhibitor). Testosterone also blocked the contractile response to serotonin [10-5 M] but not to caffeine [10-2 M]. On the other hand, testosterone inhibited the contractile responses to cyclopiazonic acid (10-6 M; a selective Ca2+ –ATPase inhibitor) or ryanodine (10-5 M; an activator of sarcoplasmic reticulum Ca2+ –release channels) associated with capacitative Ca2+ influx through non–L–type Ca2+ channels. These data suggest that by acting on the cellular membrane, testosterone interferes with the signal transduction pathway of Gq–11 protein–coupled receptors, and inhibits capacitative Ca2+ influx through both L–type and non–L–type Ca2+ channels. These effects are non–genomic, non–mediated by the intracellular androgen receptor, and not due to the conversion of testosterone to estrogens.
Key words: Testosterone, intracellular Ca2+ release, adrenergic agonists
Introducción
Múltiples estudios han informado la influencia de la testosterona en el tono vascular. La testosterona ocasiona relajación de diversos lechos vasculares de varias especies; por ejemplo, administrada en concentraciones relativamente altas ha mostrado una actividad relajante aguda en la aorta aislada de la rata,1–4 la aorta y las arterias coronarias aisladas del conejo,5 y en las arterias coronarias del perro in vivo.6 No obstante, también ha sido informado que concentraciones fisiológicas de la testosterona producen un efecto relajante en el lecho arterial mesentérico y en la aorta de la rata.7,8 Todos estos estudios indican que el efecto vaso relajante de la testosterona se manifiesta rápidamente sugiriendo una influencia no genómica, por parte del esteroide en la que han sido implicados mecanismos dependientes e independientes del endotelio.1,4–8 Para explicar el efecto agudo dilatador de la testosterona independiente del endotelio, han sido propuestos mecanismos que involucran la activación de canales de K+ en el músculo liso vascular4,5,7 y, de forma reciente, canales de Ca2+ dependientes de voltaje, de tipo L8 No obstante, otros potenciales mecanismos que expliquen la acción inhibitoria de la actividad contráctil del músculo liso vascular por parte de la testosterona, requieren ser evaluados. Particularmente se consideró como hipótesis que: la testosterona interfiere con los movimientos de Ca2+ intracelular activados por neurotransmisores u hormonas en el músculo liso vascular.
El efecto vasocontráctil causado por los agonistas adrenérgicos α1 depende de su acción sobre los reservorios intracelulares de Ca2+. Los agonistas adrenérgicos av así como otros agonistas (v.gr., serotonina) producen contracciones transitorias de arterias aisladas inmersas en una solución libre de Ca2+, mediante la liberación de Ca2+ de los reservorios intracelulares.9 La liberación de este ión conduce a la disminución o vaciamiento de los depósitos intracelulares de Ca2+ y, como consecuencia de este fenómeno, a la apertura de canales de Ca2+ de la membrana plasmática, lo que permite la entrada de Ca2+ a la célula.10–13 Así, la liberación de Ca2+ intracelular por sí misma, y la consecuente apertura de los canales de Ca2+ de la membrana celular conducen a la respuesta contráctil. La activación de la entrada de Ca2+, que es dependiente del vaciamiento de los reservorios intracelulares de Ca2+ sensibles a agonistas, se denomina entrada capacitativa (u operada por reservorios) de Ca2+.10
]]> El propósito del presente estudio fue determinar si el efecto vasodilatador de la testosterona, independiente del endotelio, se relaciona con su potencial capacidad para interferir con los procesos previamente mencionados, asociados con la actividad de los agonistas adrenérgicos a1 sobre los depósitos intracelulares de Ca2+. Utilizando segmentos anulares sin endotelio de aorta de rata, inmersos en una solución libre de Ca2+ evaluamos los efectos de la administración aguda de la testosterona sobre la respuesta contráctil transitoria de la fenilefrina que depende de los depósitos intracelulares de Ca2+, y el incremento en el tono de reposo (ITR) que se asocia con la entrada capacitativa de Ca2+ a través de la membrana. Además, investigamos farmacológicamente la participación de los receptores androgénicos y de la enzima convertidora de testoterona a estradiol, la (aromatasa), en este efecto agudo de la testosterona.
Material y métodos
Fármacos
Testosterona, hidrocloruro de fenilefrina, hidrocloruro de acetilcolina, flutamidea, hidrocloruro de diltiazem, hidrocloruro de serotonina, cafeína anhidra, ácido ciclopiazónico, ryanodina, DL–aminoglutetimida (Sigma Chemical Co.; St. Louis MO, U.S.A.); cycloheximida (ICN, Costamesa, Cal., E.U.A.) y pentobarbital sódico (Anestesal, Smith Kline Beckman, D.F., México). La testosterona, la ryanodina y la cicloheximida fueron disueltas en etanol (1%). El ácido ciclopiazónico fue disuelto en dimetilsulfóxido (DMSO; 0.1%). Los otros fármacos se disolvieron en agua bidestilada. Las diluciones subsecuentes de todos los fármacos fueron hechas en solución Krebs, y expresadas como concentración final en la cámara tisular. La pureza de los fármacos utilizados fue > 98%.
Las ratas machos Wistar (250–300g) fueron conservadas en la granja hasta su sacrificio. Los animales se mantuvieron con ciclos de luz y oscuridad de 12/12 horas en un cuarto especial a temperatura constante (22 ± 2°C), con agua y comida libremente aprovechables. Los experimentos fueron conducidos con base en protocolos aprobados por el Comité de Uso y Cuidado de Animales de nuestra Institución.
Preparación de los anillos aórticos
Los animales fueron anestesiados con pentobarbital sódico (63 mg/kg–1, i.p.); las aortas torácicas fueron rápidamente extirpadas, colocadas en solución fisiológica, limpiadas y liberadas de tejido conectivo. Las arterias aisladas se cortaron en segmentos anulares (4–5 mm. de longitud) y se colocaron en cámaras para órgano aislado (10 ml) con solución Krebs de la siguiente composición (mM): NaCl 118; KCl 4.7; KH2PO4 1.2; MgSO4 1.2; CaCl2 2.5; NaHCO3 25 y dextrosa 11.7. La solución libre de Ca2+ tuvo la misma composición que la de Krebs, excepto que se eliminó el CaCl2 y se añadió EDTA [0.1 mM]. La solución en los baños fue mantenida a 37°C, pH 7.4 y burbujeada con 95% O2 – 5% CO2. Los experimentos con cafeína se realizaron a 25°C en lugar de 37°C, porque a esta temperatura el efecto contráctil es más evidente.13 Para registrar la tensión isométrica, los anillos fueron montados en dos ganchillos de alambre de acero inoxidable y se fijaron, respectivamente, al fondo de la cámara tisular y a un transductor de desplazamiento de fuerza (Grass FTO3) conectado a un polígrafo (7D Grass; Grass Instrument Co., Quincy MA, U.S.A). Cada anillo aórtico fue sometido a una tensión inicial de 2g (elegida de experimentos preliminares en donde se relacionó la tensión inicial con la respuesta contráctil máxima obtenida con 1x10–6 M de fenilefrina). Treinta minutos después de colocar los anillos en los baños, los tejidos fueron contraídos con fenilefrina [1x10–6 M], y lavados tres veces con solución de Krebs para restaurar la tensión al nivel previo a la contractura; este proceso fue repetido dos veces más (cada 30 minutos) antes de iniciar los experimentos (la duración total de este período de estabilización fue aproximadamente 120 minutos). Todos los experimentos fueron realizados con anillos aórticos sin endotelio funcional. El endotelio fue dañado haciendo girar varias veces la parte interna de los anillos sobre la porción distal de una pinza de disección. La integridad endotelial fue valorada con acetilcolina [1x10–6 M]; los segmentos que no mostraron relajación fueron considerados carentes de endotelio.
]]> Procedimiento experimental
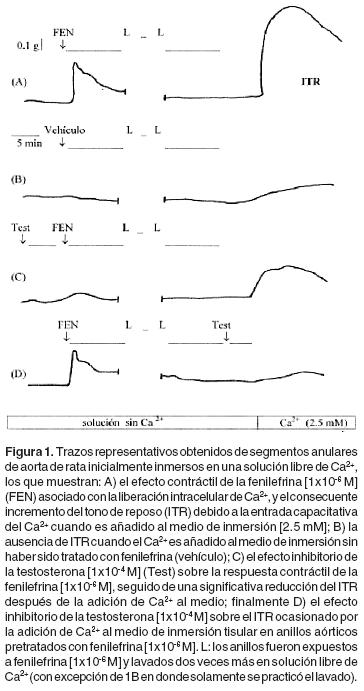
Finalizado el período de estabilización tisular, se administró fenilefrina [1x10–6 M] para contraer a los anillos aórticos en solución Krebs, después los tejidos fueron tratados con solución libre de Ca2+ por 15 minutos; entonces, se administró nuevamente fenilefrina [1x10–6 M], y aproximadamente 10 minutos después, las preparaciones arteriales fueron lavadas; la administración de fenilefrina (y el subsecuente lavado) se repitió dos veces más, al final de lo cual se añadió Ca2+ [2.5 mM] al medio de inmersión. La administración repetida de fenilefrina conduce al vaciamiento de los depósitos intracelulares de Ca2+, y, este fenómeno en turno, permite la entrada de Ca2+ desde el espacio extracelular cuando es adicionado al medio, conduciendo a un incremento en el tono de reposo (ITR)11–13 (Figura 1A). Una vez observado el ITR (aproximadamente 30 minutos), los anillos se lavaron con solución Krebs y permanecieron en ésta por una hora (período de reestabilización) antes de la administración de los fármacos en estudio. En otras dos series de experimentos, la serotonina [1x10–6 M] y la cafeína [1x10–2 M] fueron utilizadas como los agentes contráctiles de los tejidos aórticos en un medio libre de Ca2+, siguiendo el mismo protocolo que con la fenilefrina.
Después del período de reestabilización, la solución Krebs fue reemplazada por solución libre de Ca2+ y, 15 minutos después, se repitió básicamente el protocolo mencionado con anterioridad (Figura 1A), añadiendo testosterona [1x10–6 – 1x10–4 M] 5 minutos antes de la administración de fenilefrina [1x10–6 M] seguida por la adición de Ca2+ (Figura 1C), o 5 minutos antes que el Ca2+ (es decir, después de la administración de fenilefrina; Figura 1D) para evaluar la capacidad del esteroide para modificar las respuestas contráctiles; ya sea aquellas asociadas con la liberación de Ca2+ intracelular producidas por la fenilefrina, o con la entrada de Ca2+extra–celular dependiente del vaciamiento de los reservorios intracelulares de Ca2+. En otra serie de experimentos, se administró diltiazem [1x10–6 M] al medio de incubación libre de Ca2+, 5 minutos después de la administración de fenilefrina y 15 minutos antes de la adición de Ca2+, con el objetivo de evaluar la participación de los canales de Ca2+ de tipo L en la entrada de Ca2+ extracelular durante el ITR.
Tres series independientes de experimentos se realizaron para evaluar, respectivamente, la participación de los receptores androgénicos, de los estrógenos (conversión de testosterona a estrógenos por acción de la aromatasa), y la activación de la síntesis protéica, en los efectos de la testosterona sobre las contracciones obtenidas con la fenilefrina. Después del período de reestabilización, los anillos aórticos bañados en solución libre de Ca2+fueron tratados durante 30 minutos antes de la administración de testosterona [1x10–4 M] con los siguientes compuestos: flutamida [1x10–5 M] (antagonista de receptores androgénicos, cicloheximida [1x10–6 M] (inhibidor de la síntesis proteicay aminoglutetimida [1x10–6 M] (inhibidor de la enzima aromatasa. Cinco minutos después de la administración de la testosterona, se administró la fenilefrina [1x10–6 M].
Como se mencionó en series paralelas de experimentos, las respuestas contráctiles de la serotonina y la cafeína fueron estudiadas. De esta forma, los efectos contráctiles de la serotonina [1x10–5 M] y la cafeína [1x10–2 M] en los anillos aórticos inmersos en solución libre de Ca2+ y previamente tratados (5 min) con testosterona, fueron analizados después del período de reestabilización.
Por último, los efectos de la testosterona sobre las respuestas contráctiles de la ryanodina (activador del canal liberador de Ca2+ del retículo sarcoplásmico) y el ácido ciclopiazónico (inhibidor selectivo de la ATPasa de Ca2+ del retículo sarcoplásmico) en segmentos aórticos, fueron estudiados después del período de estabilización. La adición de ryanodina [1x10–5 M] o ácido ciclopiazónico [1x10–6 M] ocasiona una respuesta contráctil sostenida de lento desarrollo en los anillos aórticos en solución Krebs (con Ca2+), la cual es atribuida a la entrada capacitativa de Ca2+ a través de canales de Ca2+ tipo no L en la aorta de rata.11–13 La capacidad de la testosterona para interferir con tal entrada de Ca2+ fue evaluada estudiando los efectos de la hormona [1x10–4 M]; pretratamiento por 5 min., sobre las respuestas contráctiles al ácido ciclopiazónico [1x10–6 M] y la ryanodina [1 x10–5 M] en los anillos aórticos inmersos en una solución conteniendo Ca2+. En todos los experimentos cada segmento anular aórtico sirvió como su propio control. Una preparación tisular en cada experimento fue utilizada para determinar cambios dependientes del tiempo (control temporal). Los solventes utilizados no tuvieron efecto sobre el tono en reposo o la respuesta contráctil de los agonistas.
Análisis estadístico
]]> Los datos se presentan como la media y el error estándar de la media (media ± e.e.). En todos los casos, n es igual al número de ratas del cual se obtuvieron los segmentos de aorta. Los datos se compararon, consecuentemente, por medio de la prueba t de Student, o análisis de varianza (ANOVA) de un factor seguido por la prueba de Tukey. En todos los casos un valor de P<0.05 fue considerado estadísticamente significativo.
Resultados
Los fármacos fenilefrina [1 x10–6 M] (Figura 1), serotonina [1x10–5 M] y cafeína [1x10–2 M] provocaron efectos contráctiles transitorios en los anillos aórticos sin endotelio inmersos en una solución libre de Ca2+. Los efectos contráctiles prácticamente desaparecieron cuando cada una de las tres drogas fue administrada por tercera vez (los fármacos fueron retirados con solución libre de Ca2+ después de cada administración). La posterior adición de Ca2+ [2.5 mM] condujo al ITR en los casos de la fenilefrina (Figura 1A) y de la serotonina, pero no con la cafeína o las preparaciones de control temporal (las últimas, en presencia de solución libre de Ca2+ no fueron expuestas a los agonistas contráctiles; Figura 1B). En experimentos adicionales, se comprobó que el compartimiento de Ca2+ sensible a la cafeína fue, al menos parcialmente, compartido por la fenilefrina y la serotonina, porque la administración previa de cada una de estas drogas redujo de forma importante la respuesta contráctil de las otra13 (datos no mostrados).
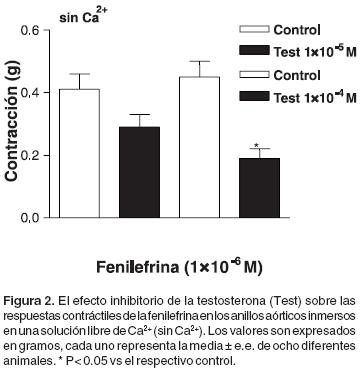
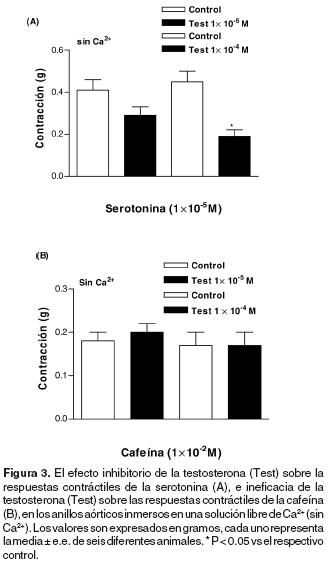
Efecto de la testosterona sobre las respuestas contráctiles de la fenilefrina, serotonina y cafeína que dependen del Ca2+ intracelular. El tratamiento (5 min.) con testosterona [1x10–5y 1x10–4 M] inhibió los efectos contráctiles obtenidos con la fenilefrina [1x10–6 M] en los anillos aórticos sin endotelio inmersos en solución libre de Ca2+ (Figuras 1C y 2). De igual forma, el tratamiento (5 min.) con testosterona [1x10–4 M] inhibió el efecto contráctil transitorio ocasionado por la serotonina [1x10–5 M] (Figura 3A). En cambio, la respuesta contráctil transitoria provocada por la cafeína [10–2 M] no fue modificada por la testosterona [1x10–4 M] (Figura 3B).
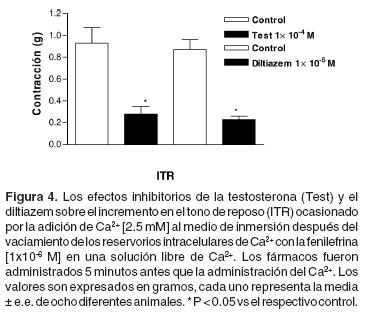
El efecto de la testosterona sobre el incremento en el tono de reposo asociado con la reincorporación de Ca2+ en el medio de incubación. El tratamiento con testosterona [1x10–4 M] por 5 minutos, redujo el ITR (Figuras 1D y 4). También, el tratamiento con diltiazem por 5 minutos [1x10–5 M], inhibió de manera significativa el ITR inducido por la adición de Ca2+ en los anillos aórticos (Figura 4).
]]>
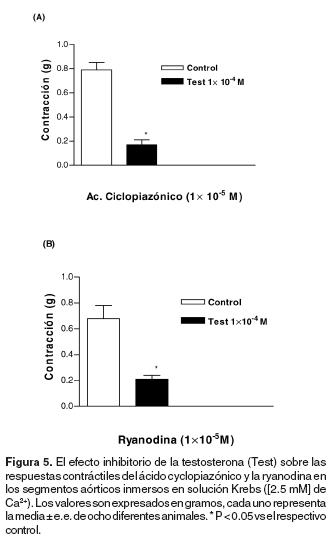
El efecto de la testosterona sobre las respuestas contráctiles del ácido ciclopiazónico y de la ryanodina en los anillos aórticos inmersos en una solución con Ca2+. El ácido ciclopiazónico y la ryanodina produjeron respuestas contráctiles de desarrollo lento y sostenido en los anillos aórticos inmersos en solución Krebs con Ca2+ [2.5 mM] (Figura 5). Los efectos contráctiles de ambas drogas fueron inhibidos significativamente con el tratamiento por 5 minutos con testosterona [1x10–4 M] (Figura 5).
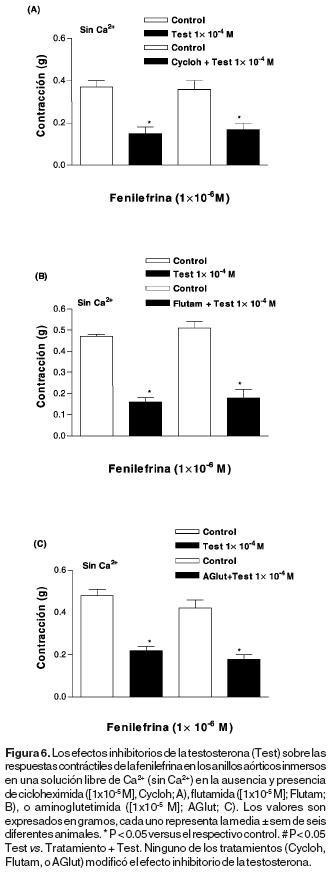
Los efectos de la flutamida, la cicloheximida y la aminoglutetimida sobre la actividad de la testosterona como inhibidor de la respuesta contráctil de la fenilefrina dependiente de Ca2+ intracelular. El tratamiento con cicloheximida [1x10–5 M] (Figura 6A), flutamida [1x10–5 M] (Figura 6B), o aminoglutetimida [1x10–5 M] (Figura 6C) no modificó, en cada caso, el efecto inhibitorio de la testosterona sobre la respuestas contráctiles de la fenilefrina en los anillos aórticos inmersos en una solución libre de Ca2+.
Discusión
]]> Los presentes resultados muestran que la testosterona inhibe la respuesta contráctil transitoria producida por el agonista adrenérgico a1, fenilefrina, en los segmentos aórticos sin endotelio de la rata, inmersos en una solución libre de Ca2+, y el ITR observado por la adición de Ca2+ al medio (después de vaciar los depósitos intracelulares de Ca2+ mediante la administración repetida de fenilefrina). Estos efectos inhibitorios, fueron observados 5 minutos después de la exposición a la hormona. Por la rapidez de su aparición pueden ser interpretados como efectos no genómicos, y en concordancia con esta interpretación, encontramos que el inhibidor de la síntesis proteica, cicloheximida, no modificó los efectos inhibitorios de la testosterona. Es decir, además de la rapidez de la respuesta, el hecho de que la cicloheximida no inhiba el efecto relajante de la testosterona, sugiere que, durante el tiempo en cuestión, no hubo participación de mecanismos genómicos. Así, dos de los factores que se consideran importantes para clasificar una respuesta como no genómica (curso temporal rápido e insensibilidad a los fármacos inhibidores de la síntesis proteica)14,15 han sido identificados en el caso de los efectos inhibitorios de la testosterona sobre las respuestas contráctiles mencionadas. Además, los efectos de la testosterona no fueron modificados por el antagonista selectivo del receptor intracelular androgénico, flutamida,16 ni por el inhibidor de la aromatasa, aminoglutetimida,17 lo cual excluye, por un lado la participación del receptor mencionado, y por otro, la participación del estradiol formado a partir de la testosterona, en tales efectos. Estos resultados son congruentes con estudios previos, en la aorta y la arteria coronaria de conejo,5 y en el lecho arterial mesentérico de la rata,7 los cuales muestran que el efecto vasodilatador de la testosterona no fue afectado por la flutamida ni por la aminoglutetimida. Asimismo, varios estudios recientes indican que la administración de la testosterona ocasiona una vasodilatación rápida (interpretada como no genómica) e independiente del receptor androgénico intracelular en la aorta aislada de rata1,4 y en las arterias coronarias de conductancia y resistencia de perro.6En el efecto vasodilatador de la testosterona han sido propuestos mecanismos dependientes e independientes del endotelio. En la aorta1,4 y el lecho mesentérico de la rata7y en los vasos coronarios epicardios y de resistencia del perro6 la dilatación aguda de la testosterona ha sido atribuida parcialmente al óxido nítrico derivado del endotelio, pero se acepta también la participación de un mecanismo independiente del endotelio. Por otra parte, hay evidencia de que la testosterona ocasiona relajación independiente del endotelio en la aorta y arteria coronaria aislada del conejo,5 en la aorta torácica de la rata2 y en la arteria coronaria porcina.18
De esta forma, nuestros resultados documentan un efecto inhibidor de la actividad contráctil del músculo liso vascular, por la testosterona, que es independiente del endotelio y del receptor androgénico.
Las concentraciones de testosterona utilizadas en el presente estudio fueron elegidas porque son similares (concentraciones micromolares) a aquellas con las que otros investigadores han informado haber obtenido una respuesta vasodilatadora en vasos sanguíneos de variadas especies.1,3–6 Claramente, estas concentraciones son mayores que aquellas consideradas fisiológicas; por ejemplo, 10–35 nM en el adulto humano normal.19 No obstante, hay evidencia de que la administración aguda de bajas concentraciones (nanomolares) de testosterona, ocasiona vaso dilatación del lecho arterial mesentérico y de la aorta de la rata;7,8 lo cual sugiere que concentraciones fisiológicas de testosterona pueden influenciar el tono vascular. También, estudios recientes muestran que dosis bajas de testosterona ocasionan vasorelajación en arterias coronarias humanas.20,21 Entonces, es posible que en relación con la concentración haya considerable variación en la capacidad de la testosterona para influenciar el tono vascular en diferentes lechos vasculares y especies.
En el músculo liso vascular, la activación de los receptores adrenérgicos a1 causa la hidrólisis, mediada por la fosfolipasa C, del fosfolípido de la membrana, fosfatidilinositol 4,5 bifosfato (PIP2), produciendo dos segundos mensajeros: inositol 1,4,5–trifosfato (IP3) y diacilglicerol (DAG).9 El IP3 se enlaza a receptores localizados en el retículo sarcoplásmico y libera calcio,9,22 el cual es considerado responsable de las contracciones transitorias producidas por los agonistas en un medio libre de Ca2+. Por lo tanto, la inhibición por la testosterona de la respuesta contráctil transitoria de la fenilefrina en los anillos aórticos de rata, puede ser el resultado de la interferencia de la hormona en algún lugar de la vía de transducción de la señal de los receptores adrenérgicos a1. Asimismo, la respuesta contráctil de la serotonina en los anillos aórticos inmersos en una solución libre de Ca2+, fue inhibida por la testosterona. Este efecto contráctil es mediado por receptores 5–HT2 25 acoplados a proteínas Gq–11 que movilizan Ca2+ intracelular (vía hidrólisis de fosfatidilinositol) de almacenes comunes a los agonistas adrenérgicos a1 y a la cafeína. De esta manera, la testosterona inhibió la actividad de agonistas que actúan en diferentes receptores, adrenérgicos y serotonérgicos, pero que poseen un mecanismo similar de transducción de la señal. Por lo tanto, la testosterona parece afectar la función de dos poblaciones diferentes de receptores de membrana, o la actividad de los elementos de una vía de transducción común de la señal. No obstante, la testosterona no ejerció una acción directa inhibitoria de la liberación de Ca2+ en el retículo sarcoplásmico, porque no modificó el efecto contráctil de la cafeína en los anillos aórticos inmersos en solución libre de Ca2+. Por lo anterior, el efecto inhibitorio de la testosterona sobre la respuesta contráctil transitoria de la fenilefrina (y de la serotonina) puede relacionarse con los elementos involucrados en la vía de transducción de la señal del receptor adrenérgico a1 (y del receptor 5–HT2), con la exclusión de un efecto inespecífico en el retículo sarcoplásmico.
La testosterona también inhibió el ITR. En la aorta de rata, ha sido establecido que la administración de fármacos agonistas de receptores adrenérgicos a, conduce al vaciamiento de los reservorios intracelulares de Ca2+ y, consecuentemente, a la apertura de canales de Ca2+ de la membrana celular.11–13 Así, los reservorios internos de Ca2+ desempeñan un papel crucial en la respuesta contráctil de las células del músculo liso vascular, porque los almacenes deprivados de Ca2+ provocan la llamada entrada capacitativa, u operada por reservorio, de Ca2+ extracelular.10 En este proceso participan canales de Ca2+ dependientes de voltaje de tipo L23 y canales catiónicos no selectivos, tipo no L,24 en relación con el músculo liso estudiado. En concordancia con Noguera y cols.,11-13 los presentes resultados muestran que el ITR consecuente a la disminución de los reservorios intracelulares de Ca2+ por la fenilefrina es, al menos parcialmente, debido a la entrada capacitativa de Ca2+ a través de canales de Ca2+ dependientes del voltaje de tipo L, dado que el efecto fue significativamente reducido con diltiazem. Como la testosterona inhibió de manera similar este proceso, consideramos que actúa inhibiendo la conductancia de los canales de Ca2+ de tipo L. En este sentido, recientemente ha sido informado que la testosterona inhibe la entrada de Ca2+ a través de canales de Ca2+ de tipo L en la aorta de rata.8 Por otra parte, la testosterona también inhibió los efectos contráctiles causados por el ácido ciclopiazónico y la ryanodina en anillos aórticos inmersos en una solución normal de Krebs (con Ca2+). En la aorta de rata, los efectos contráctiles de los fármacos se asocian con el ingreso capacitativo de Ca2+ extracelular a través de canales de Ca2+ tipo no L, estimulados por el previo vaciamiento de los reservorios intracelulares de Ca2+.11–13 En un medio libre de Ca2+, no se observa la respuesta contráctil de estas sustancias, lo cual indica que la contracción observada en la presencia de Ca2+ extracelular es ocasionada por la entrada del catión al través del sarcoplasma. En consecuencia, la testosterona además de bloquear los canales de Ca2+ de tipo L, puede también disminuir la conductancia de los canales de Ca2+ tipo no L, o canales catiónicos no selectivos, que son responsables de los efectos contráctiles del ácido ciclopiazónico y de la ryanodina.
Los resultados mencionados, considerados en conjunto, permiten sugerir que la testosterona modifica la función de moléculas incluidas en la membrana del músculo liso vascular, como los canales de Ca2+ de tipo L y tipo no L, así como elementos de la vía de transducción de la señal de los receptores acoplados a proteínas Gq–11, posiblemente los receptores mismos, las proteínas G o la fosfolipasa C. Esto puede ser consecuencia de la habilidad de la testosterona para disolverse en la membrana plasmática, causando cambios en la fluidez de la membrana y en el contenido y balance de los lípidos que circundan las proteínas de la membrana, afectando sus funciones. El hecho de que los efectos son observados solamente con altas concentraciones de testosterona sugiere que no son consecuencia de la interacción del esteroide con un receptor específico, o de otra forma, que son mediados por un receptor insensible a la flutamida con baja afinidad por la testosterona. Cabe mencionar que desde la década del 70, ha sido reiteradamente propuesto que los esteroides pueden concentrarse en la bicapa de fosfolípidos alterando la fluidez de la membrana.26–29 Además, recientemente, fue establecido en la primera reunión internacional sobre las respuestas rápidas de las hormonas esteroides (First Internacional Meeting on Rapid Responses to Steroid Hormones,30 que los efectos no genómicos de los esteroides pueden ser ocasionados con altas concentraciones de ellos, sin la participación necesaria de receptor, por la modulación de la función proteínica que refleja cambios en las propiedades fisicoquímicas de la membrana plasmática.30
En conclusión, nosotros hemos demostrado que las concentraciones altas de la testosterona inhiben los efectos contráctiles asociados con la liberación de calcio intracelular producida por la estimulación de receptores acoplados a proteínas Gq–11 y con la entrada capacitativa de Ca2+ a través de canales de Ca2+ de tipo L y de tipo no L.
Referencias
]]>1. Costarella CE, Stallone JN, Rutecki GW, Whittier FC. Testosterone causes direct relaxation of rat thoracic aorta. J Pharmacol Exp Ther 1996; 277:34–39. [ Links ]
2. Perusquía M, Hernández R, Morales MA, Campos MG, Villalon CM. Role of endothelium in the vasodilating effect of progestins and androgens on the rat thoracic aorta. Gen Pharmac 1996; 27:181–185. [ Links ]
3. Honda H, Unemoto T, Kogo H. Different mechanisms for testosterone–induced relaxation of aorta between normotensive and spontaneously hypertensive rats. Hypertension 1999; 34:1232–1236. [ Links ]
4. Ding A, Stallone JN. Testosterone–induced relaxation of rat aorta is androgen structure specific and involves K+ channel activation. J Appl Physiol 2001; 91:2742–2750. [ Links ]
5. Yue PY, Chatterjee K, Beale C, Wilson PA, Collins P. Testosterone relaxes rabbit coronary arteries and aorta. Circulation 1995; 91:1154–1160. [ Links ]
6. Chou TM, Sudhir K, Hutchison SJ, Ko E, Amidon TM, Collins P, Chatterjee K. Testosterone induces dilation of canine coronary conductance and resistance arteries in vivo. Circulation 1996; 94:2614–2619. [ Links ]
7. Tep–areenan P, Kendall DA, Randall MD. Testosterone–induced vasorelaxation in the rat mesenteric arterial bed is mediated predominantly via potassium channels. Br J Pharmacol 2002; 135:735–40. [ Links ]
8. Tep–areenan P, Kendall DA, Randall MD. Mechanisms of vasorelaxation to testosterone in the rat aorta. Eur J Pharmacol 2003; 465:125–132. [ Links ]
9. Varma DR, Deng XF. Cardiovascular a1–adrenoceptor subtypes: functions and signaling. Can J Physiol Pharmacol 2000;78:267–292. [ Links ]
10. Putney JW. Capacitative calcium entry revisited. Cell Calcium 1990; 11:611–634. [ Links ]
11. Noguera MA, D'Ocon MP. Evidence that depletion of internal calcium stores sensitive to noradrenaline elicits a contractile response dependent on extracellular calcium in rat aorta. Br J Pharmacol 1993; 110:861–867. [ Links ]
12. Noguera MA, Ivorra MD, Chulia S, D'Ocon MP. Capacitative Ca2+ entry associated with a1–adrenoceptors in rat aorta. Naunyn–Schmiedeberg's Arch Pharmacol 1996; 355:1–7. [ Links ]
13. Noguera MA, Madrero Y, Ivorra MD, D'Ocon P. Characterization of two different Ca2+ entry pathways dependent on depletion of internal Ca2+ pools in rat aorta. Naunyn–Schmiedeberg's Arch Pharmacol 1998; 357:92–99. [ Links ]
14. Wehling M. Specific, nongenomic actions of steroid hormones. Annu Rev Physiol 1997; 59:365–393. [ Links ]
15. Falkenstein E, Tillmann H–C, Christ M, Feuring M, Wehling M. Multiple actions of steroid hormones – A focus on rapid, nongenomic effects. Pharmacol Rev 2000; 52:513–555. [ Links ]
16. McLeod DG. Antiandrogenic drugs. Cancer 1993; 71:1046–1049. [ Links ]
17. Perez N, Borja J. Aromatase inhibitors: clinical pharmacology and therapeutic implications in breast cancer. J Int Med Res 1992; 20:303–312. [ Links ]
18. Deenadayalu VP, White RE, Stallone JN, Gao X, Garcia AJ. Testosterone relaxes coronary arteries by opening the large–conductance, calcium–activated potassium channel. Am J Physiol Heart Circ Physiol 2001; 281:H1720–H1 727. [ Links ]
19. Wilson JD. Androgens. En: The Pharmacological Basis of Therapeutics. Hardman JD, Limbird LE, Molinoff PB, Ruddon RW, Goodman Gilman A (Eds.). McGraw–Hill, 9a. Edición, New York, EUA, 1996, pp 1441–1457. [ Links ]
20. Webb C, McNeill J, Hayward C, Zeigler D, Collins P. Effects of testosterone on coronary vasomotor regulation in men with coronary heart disease. Circulation 1999; 100:1690–1696. [ Links ]
21. Giuseppe M, Rosano C, Leonardo F, Pagnotta P, Pelliccia F, Panina G, et al. Acute anti–ischemic effect of testosterone in men with coronary artery disease. Circulation 1999; 99:1666–1670. [ Links ]
22. Karaki H, Ozaki H, Hori M, Mitsui–Saito M, Amano K, Harada K, et al. Calcium movements, distribution, and functions in smooth muscle. Pharmacol Rev 1997; 49:157–230. [ Links ]
23. Felder CC, Singer–Lahat D, Mathes C. Voltage–independent calcium channels. Regulation by receptors and intracellular calcium stores. Biochem Pharmacol 1995; 48:1997–2004. [ Links ]
24. Fasolato C, Innocenti B, Pozzan T. Receptor–activated Ca2+ influx: how many mechanisms for how many channels? Trends Pharmacol Sci 1994; 15:77–83. [ Links ]
25. Okoro EO. Overlap in the pharmacology of L–type Ca2+ channel blockers and 5–HT2 receptor antagonists in rat aorta. J Pharm Pharmacol 1999; 51:953–957. [ Links ]
26. Willmer EN. Steroids and cell surfaces. Biol Rev 1961;36:368–398. [ Links ]
27. Clarke R, Van den Berg HW, Murphy RF. Reduction of the membrane fluidity of human breast cancer cells by tamoxifen and 17b–estradiol. J Natl Cancer Inst 1990;82:1702–1705. [ Links ]
28. Reddy AG, Shivaji S, Gupta PD. Effect of estradiol on the membrane fluidity of the rat vagina endothelial cells. J Steroid Biochem 1989; 33:1229–1233. [ Links ]
29. Shivaji S, Jagannadham MV. Steroid–induced perturbation of membranes and its relevance to sperm acrosome reaction. Biochem Biophys Acta 1992; 1108:99–109. [ Links ]
30. Falkenstein E, Norman AW, Wehling M. Mannheim classification of nongenomically initiated (rapid) steroid action (s). J Clin Endo Met 2000; 85:2072–2075. [ Links ] ]]>