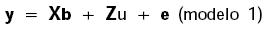
Efectos ambientales y parámetros genéticos de variables de crecimiento para un rebaño de ciervo rojo (Cervus elaphus) en cautiverio
Environmental effects and genetic parameters on growth traits for a herd of farmed red deer (Cervus elaphus)
Rodolfo Ramírez-Valverdea, Alejandra Sánchez-Cervantesb, José G. García-Muñiza, Rafael Núñez-Domíngueza, Vicente Lemus-Ramírezb
a Departamento de Zootecnia, Posgrado en Producción Animal, Universidad Autónoma Chapingo, Carretera México-Texcoco km 38.5. Chapingo, Estado de México. 56230. rodolforv@correo.chapingo.mx. Correspondencia al primer autor.
]]> b Centro de Enseñanza, Investigación y Extensión en Producción Animal en Altiplano, Facultad de Medicina Veterinaria y Zootecnia de la UNAM.
Recibido el 7 de octubre de 2010
Aceptado el 9 de febrero de 2011
RESUMEN
El objetivo fue estimar efectos ambientales y parámetros genéticos de variables de crecimiento en un rebaño de ciervo rojo en cautiverio de la zona templada del centro de México. Los animales crecieron en un sistema de pastoreo rotacional intensivo de praderas templadas mixtas de gramíneas y leguminosas. Las variables evaluadas fueron los pesos al nacimiento (PN), al destete ajustado a 100 d (PD) y al año (PA). Los análisis fueron realizados mediante el programa MTDFREML. Los modelos animales finales consideraron el efecto fijo de año-sexo, y la edad de la madre al parto como covariable para PN y PD, y los efectos genéticos aleatorios directo (PN, PD y PA) y materno (PD). Las medias y desviaciones estándar de PN, PD y PA fueron 8.7±1.2, 42.4±5.6 y 80.8±13.3 kg. Las machos fueron más pesados (P<0.05) que las hembras (4.6, 9.9 y 21.9 %, para PN, PD y PA). En todas las características se detectaron diferencias entre grupos contemporáneos (P<0.05). El efecto lineal de edad de la madre al parto fue importante (P<0.05) para PN y PD. Los valores estimados de heredabilidad directa fueron 0.00±0.08, 0.41 ±0.33 y 0.17±0.15, para PN, PD y PA; mientras que la heredabilidad materna de PD fue 0.22±0.16. Con base en la variabilidad genética aditiva estimada de PD y PA, los resultados sugieren la posibilidad de mejoramiento genético en estas características por medio de selección; sin embargo, estos resultados no son concluyentes, debido a la limitada cantidad y calidad de la información utilizada.
Palabras clave: Cervus elaphus, Modelo animal, Heredabilidad.
ABSTRACT
]]> The objective was to estimate environmental effects and genetic parameters on growth traits in a flock of farmed red deer of the central Mexican highland. Animals were raised on an intensive rotational grazing system of grass-legume mixed temperate pastures. Evaluated traits were birth (BW), weaning adjusted to 100 d (WW) and yearling (YW) weights. Analyses were performed using the MTDFREML program. Final univariate animal models considered the fixed effect of sex-year, and calving age as a covariate for BW and WW, and the random direct (BW, WW and YW) and maternal (WW) genetic effects. Means and standard deviations for BW, WW and YW were 8.7±1.2, 42.4±5.6 and 80.8±13.3 kg. Males were higher (P<0.05) than those of females (4.6, 9.9 and 21.9 %, for BW, WW and YW). There were differences between contemporary groups for all traits (P<0.05). The linear effect of hind calving age was important (P<0.05) for both BW and WW. Estimates of direct heritability were 0.00±0.08, 0.41 ±0.33 and 0.17±0.15 for BW, WW and YW. Maternal heritability of WW was 0.22±0.16. Based on the estimated genetic additive variability of WW and YW in this herd, the results suggest the possibility for genetic improvement of those traits through selection; however, these results are not definitive given the limited quantity and quality of the used information.Key words: Cervus elaphus, Animal model, Heritability.
Los ciervos evolucionaron durante el periodo medio del Mioceno de hace 16 millones de años(1). La mayoría de las especias animales de interés zootécnico han sido domesticadas por miles de años, mientras que los ciervos han sido utilizados durante los 40 años más recientes. La utilización de ciervos en confinamiento comenzó a principios de 1970 en Nueva Zelanda, el Reino Unido y Australia(2,3,4), y actualmente esta especie representa una alternativa para la producción de carne. La carne de ciervo es reconocida en algunos países de Europa como un producto de uso tradicional, que es promovido por su alta calidad y efectos benéficos para la salud humana, debido a su bajo contenido de colesterol(5).
El mejoramiento genético sistemático de razas y líneas de ciervos es probable que en el futuro aumente en importancia, a medida que se agote el suministro internacional de líneas superiores(6). No todos los animales se adaptan exitosamente al cautiverio y uso por los humanos, por lo que es posible que una variación considerable surja en generaciones subsecuentes que permita una respuesta adaptativa al cautiverio(1). La investigación basada en el análisis de registros a nivel de rebaños ha mostrado variación genética considerable para variables de crecimiento en ciervo rojo(2,3,7); sin embargo, las pocas estimaciones realizadas provienen de hatos relativamente pequeños y con poca exactitud en los parámetros genéticos estimados. Adicionalmente, la expresión de la variabilidad genética puede variar con las diferentes condiciones ambientales.
En 1994, el gobierno federal mexicano importó de Nueva Zelanda un rebaño de alrededor de 900 ciervos rojos (Cervus elaphus), para promover su cría en cautiverio como una alternativa de diversificación en las empresas de producción ganadera(8). Los animales fueron distribuidos entre varias instituciones de investigación. La producción de ciervo rojo puede ser una buena opción para algunos pequeños productores o para complementar empresas ganaderas convencionales. El número de ganaderos utilizando la producción de ciervos ha aumentado en este país, debido a que los productores reconocen el beneficio económico de su uso potencial.
En México, se ha publicado poco acerca de la caracterización genética para producción en ciervo rojo(4). Por tanto, es importante evaluar esta especie en los principales sistemas de producción e identificar efectos ambientales que afectan su productividad. El conocimiento de parámetros genéticos y fenotípicos es necesario para evaluar las posibilidades de mejoramiento genético en empresas de producción de carne de ciervo, y para proveer a ganaderos y genetistas de herramientas de selección objetivas. El objetivo de este estudio fue estimar efectos ambientales y parámetros genéticos de variables de crecimiento en un rebaño de ciervo rojo en cautiverio de la zona templada del centro de México.
El rebaño de ciervo rojo estudiado fue parte de los animales importados de Nueva Zelanda a México en 1994. Este rebaño fue establecido en el "Centro de Enseñanza, Investigación y Extensión en Producción Bovina y Caprina" de la Universidad Nacional Autónoma de México. El lugar está localizado en la región central del país (Tepotzotlán, México, 2,450 msnm, 19° 43' N y 94° 14' O), con clima templado.
Las variables evaluadas fueron los pesos al nacimiento (PN), al destete ajustado a 100 d (PD) y al año (PA). Los registros de pesos usados en los análisis fueron obtenidos de animales nacidos entre 1996 y 2003 (218 machos y 210 hembras), provenientes de 8 sementales y 100 hembras. Las edades de las madres al parto variaron de 23.2 a 126.2 meses (1 a 8 partos). Los registros de crecimiento usados en este estudio fueron 428, 383 y 270 para PN, PD y PA. El archivo de pedigrí incluyó 530 animales nacidos de 1992 a 2003; el número total de sementales y vientres considerados en este estudio fue de 22 y 102, respectivamente.
El rancho experimental de ciervos tuvo 11 divisiones de una hectárea cada uno. Los animales pastorearon todo el año en praderas mixtas de gramíneas y leguminosas, que consistieron aproximadamente de 70 % alfalfa (Medicago sativa), y 30 % rye grass (Lolium perenne) y orchard grass (Dactylisglomerata). El área de pastoreo tuvo riego y fertilización (nitrógeno y fósforo) cuatro veces durante el otoño-invierno de cada año. Los animales estuvieron la mayor parte del año en las praderas, y suplemento concentrado se ofreció sólo a hembras en lactación. Después de que parió el 80 % de las hembras reproductivas (principios de julio), se les ofreció un concentrado alimenticio que contenía 14 % de proteína cruda (0.2 a 0.4 kg por animal por día) hasta 15 días después del destete. Aunque los cervatos tuvieron acceso al alimento suplementario hasta el destete, no se observó consumo del concentrado asignado a sus madres. El amamantamiento de cervatos por hembras diferentes a sus madres se observó pocas veces y por periodos cortos.
]]> Los partos de cada año ocurrieron naturalmente en periodos de dos meses, la mayoría durante junio y julio. Durante los años estudiados, el porcentaje de nacimientos por mes fue 7.6, 67.1, 23.5 y 1.8 para mayo, junio, julio y agosto. La mortalidad promedio al nacimiento fue 8 % y los animales sobrevivientes fueron pesados dentro de las primeras 24 h de nacidos. Durante la época de empadre (octubre a diciembre), las hembras se asignaron en grupos de 30 a 40 con un semental. Al final de la época del empadre, los sementales se removieron de sus respectivos grupos reproductivos y se reunieron para el pastoreo invernal como un solo grupo. Después del destete (aproximadamente a 100 días de edad), los cervatos machos y hembras estuvieron pastoreando como un grupo en divisiones separadas hasta diciembre (aproximadamente a 200 días de edad). Desde enero del año siguiente hasta que los cervatos hembras tuvieron un año de edad pastorearon conjuntamente con el grupo de las hembras adultas, y los machos se asignaron en una división separada como otro grupo.Durante el tiempo del trabajo, el tamaño del rebaño permaneció relativamente estable, con un promedio de 97 hembras reproductivas. El rebaño tuvo la mayoría de las hembras y machos originales en reproducción, y pocos reemplazos se utilizaron cada año. Como consecuencia, el 65 % de los animales con registros de PN fueron hermanos completos. Los animales nacidos en México y usados como reemplazos fueron seleccionados principalmente con base en su temperamento, salud y apariencia. Hasta ahora, el criterio usado para asignar machos a las hembras ha sido sólo con el fin de evitar apareamientos entre animales cercanamente emparentados.
Los análisis estadísticos para estimar parámetros genéticos y efectos ambientales se realizaron mediante un modelo animal y el programa MTDFREML(9). En cada característica evaluada, un modelo animal univariado se ajustó para identificar efectos fijos y aleatorios importantes (P<0.05).
Para seleccionar efectos aleatorios en las características evaluadas, los modelos iniciales incluyeron los efectos directos, maternos y ambientales permanentes. La significancia de incluir un efecto aleatorio en el modelo fue probada mediante la prueba de proporción de verosimilitudes(10). Un efecto fue considerado importante cuando su inclusión en el modelo causó un incremento significativo (P<0.05) en el log de verosimilitud, comparado con un modelo en el cual el efecto fue omitido. Cuando -2 veces la diferencia entre el log de verosimilitud fue mayor que un valor crítico de una distribución Ji cuadrada con un grado de libertad, el factor aleatorio adicional se consideró significativo.
Para seleccionar efectos fijos en las características evaluadas, los modelos iniciales incluyeron el sexo del cervato, el año de nacimiento, el orden de nacimiento anual como una covariable o como una variable de clasificación conjuntando los registros de los animales dentro de dos grupos (nacimientos tempranos y tardíos), y edad de la madre al parto (lineal y cuadrática). Para probar la significancia de los efectos, contrastes y sus errores estándar fueron estimados con el programa MTDFREML(9). Se usaron pruebas del estadístico t con un grado de libertad para probar la significancia (P<0.05) de contrastes específicos. Para cada variable, si los contrates fueron estimados como no significativos, el efecto respectivo fue removido del modelo final.
En notación matricial, los modelos finales para PN y PA (modelo 1) y PD (modelo 2) fueron:

donde y es el vector para los registros de pesos; b es el vector de efectos fijos para año de nacimiento y sexo del cervato (grupo contemporáneo) en las tres características, más la covariable lineal de edad de la madre al parto para PN y PD; u es el vector de efectos genéticos aditivos directos; m es el vector de los efectos genéticos maternos; e es el vector de efectos residuales; y X, Z, y M son matrices de incidencia que asocian los registros de pesos con los vectores correspondientes. Las suposiciones de los modelos fueron: E[y] = Xb, E[u] = E[m] = E[e] = 0, y

Las medias y desviaciones estándar de las características de crecimiento estudiadas se muestran en el Cuadro 1, mientras que los contrastes significativos (P<0.05) y sus errores estándar de los efectos ambientales se presentan en el Cuadro 2. Todas las variables estudiadas fueron afectadas (P<0.05) por el sexo del cervato, el año de nacimiento y la interacción sexo del cervato por año de nacimiento. El efecto lineal de edad de madre al parto afectó (P<0.05) los PN y PD. El efecto del orden de nacimiento como variable de clasificación o como covariable, y el efecto cuadrático de la edad de la madre al parto no fueron significativos (P>0.05) en las características evaluadas; sin embargo, para el último efecto se detectaron tendencias a la significancia (valores de t fueron 1.4 y 1.2 para PN y PD).


Las medias de los pesos calculados en este estudio indicaron que la producción de carne de ciervo en zonas templadas de México puede ser un sistema económicamente viable. Los valores de crecimiento en este estudio fueron similares a los publicados en otros ambientes con el uso de ciervo rojo en cautiverio. Para rebaños en Escocia con ciervo rojo en pastoreo(2) se estimaron 7.9, 38.9 y 55.6 kg para PN, PD (109 días) y peso a los 11 meses de edad. En información publicada de ocho rebaños en pastoreo en el Reino Unido, con cervatos en confinamiento después del destete; las medias entre rebaños variaron de 8.3 a 9.4 kg para PN, de 41.6 a 51.2 kg para PD (98 a 108 d), y de 67.1 a 80.1 kg para el peso entre 10 y 11 meses de edad(7). En un rebaño de México con animales y ambiente similar al de este estudio se estimaron 8.9, 36.4 y 68.9 kg para PN, PD y PA(4).
Los cervatos machos tuvieron 4.6, 9.9 y 21.9 % mayores PN, PD y PA que las hembras. Este patrón en las diferencias también ha sido observado en otros estudios con ciervo rojo en cautiverio. En fallow deer, se estimaron pesos mayores en machos que en hembras(11) para PN, PD y PA (7.2, 13.0 y 25.1 %, respectivamente). En ciervo rojo, la superioridad de machos sobre hembras ha sido estimada en 8, 10 y 17 %(4), y 5.4, 11.3 y 17.9 %(7) para PN, PD y PA, respectivamente.
Las diferencias de pesos entre los años de nacimiento y las interacciones de año por sexo (grupo contemporáneo) variaron dependiendo de la característica estudiada. Por ejemplo, los contrastes respectivos en relación con las medias mínimas y máximas fueron 5.9 y 12.3 % para PN, 13.4 y 22.6 % para PD, y 22.5 y 42.7 % para PA. Estas diferencias en las características estudiadas son típicas de sistemas de producción altamente dependientes de los factores ambientales, como los sistemas en pastoreo, y es importante considerarla en la modelación estadística para evitar valores estimados sesgados de los efectos genéticos. Diferencias similares también han sido publicadas en otros estudios con animales domésticos, incluyendo ciervos rojos(2,4,7).
Los PN y PD aumentaron conforme la edad de la madre incrementó, con pendientes de 6.6 y 71.8 g/mes, respectivamente. Este patrón de aumentos es similar al comúnmente observado en otros mamíferos domésticos; sin embargo, en este estudio el efecto cuadrático sólo tendió a ser significativo (P>0.05). Dentro del rango de edad y número de parto de las hembras en este estudio, parece que no se ha obtenido el máximo de peso en sus cervatos. Las hembras de ciervo rojo en hábitats naturales pueden vivir hasta 18 a 20 años de edad, y los resultados en cautiverio sugieren que las hembras pueden permanecer reproductivas aproximadamente hasta los 16 años de edad(12). En el presente estudio, la edad de las hembras reproductivas varió entre 1.9 y 10.5 años, y con una tendencia a la significancia del efecto cuadrático de la edad de la madre, lo que sugiere que la edad de la madre para el máximo peso de sus cervatos está cercana a alcanzarse. En otros estudios con ciervo rojo también se ha estimado un patrón similar de hembras más viejas criando cervatos más pesados(2,4,7).
]]> Los cervatos que nacen primero en la estación de partos pueden mostrar mejor crecimiento que los que nacen hacia el final de la estación. La razón por este efecto estacional no es clara. Los partos tardíos en la estación puede resultar en menor disponibilidad de nutrientes para soportar la secreción láctea y por tanto podrían estar en desventaja con los nacidos a inicio de la estación; además, para estos últimos el efecto del amamantamiento a otras hembras podría provocar mayor crecimiento de estos cervatos(13). El orden de nacimiento en la estación de partos ha sido encontrado como un efecto ambiental importante en diversos estudios con ciervos(13,14,15) y generalmente es incluido en los modelos para la evaluación genética de características de crecimiento(6). En este estudio, la inclusión de este efecto no fue significativa (P>0.05), por lo que no se utilizó en los modelos finales. Esto pudo deberse a las estaciones de parto cortas en este rebaño, lo que no permitió variabilidad en condiciones ambientales suficiente para detectar diferencias, además que el amamantamiento por otras hembras diferentes a las madres no fue común. Resultados similares con ciervo rojo fueron publicados en otro estudio(16).Los valores estimados de (co)varianzas y parámetros genéticos de las características estudiadas se muestran en el Cuadro 3. Las proporciones de varianza ambiental respecto al total de la varianza para PN, PD y PA fueron 100, 66 y 83 %. El estimado de heredabilidad directa para PN fue cero, y para PD y PA los estimados fueron de magnitud moderada (0.41 ± 0.33 y 0.17 ± 0.15); el de heredabilidad materna para PD fue de magnitud moderada (0.22 ± 0.19), y la correlación genética entre efectos directo y materno fue alta y negativa (-0.98 ± 0.17).

En poblaciones en ambientes naturales, el PN es una característica asociada con la aptitud y tiene una heredabilidad baja(17). Las condiciones ambientales difíciles de poblaciones naturales están asociadas con una selección intensa para incrementar el PN, lo que provoca una baja varianza genética aditiva(18). Además, otros autores han mencionado que es difícil obtener valores estimados precisos de varianza genética aditiva en características con baja heredabilidad(19). Similar a la heredabilidad de cero obtenida en este estudio para PN, para ciervos en cautiverio se estimaron heredabilidades que oscilaron entre 0.00 y 0.17(20); sin embargo, se han publicado valores mayores para esta característica (0.11 a 0.31) en fallow deer(11) y ciervo rojo en cautiverio(2,4,7) o en condiciones naturales(21). Las razones de falta de variabilidad genética aditiva para PN en este estudio no son claras. Algunos autores han mencionado como factores que pueden reducir la exactitud de los valores estimados de heredabilidad, las fallas para contabilizar la variación por el orden de nacimiento de los cervatos(6), la heterosis(7), los efectos genéticos maternos(22,23), y la varianza de dominancia(21). Adicionalmente, la baja exactitud de los valores estimados de heredabilidad también ocurren si el análisis está basado en un número de animales relativamente pequeño(4,7,11) o cuando existen errores en la identificación de la paternidad(4,21,23).
Algunos de los posibles factores que contribuyeron para no detector variabilidad genética aditiva de PN en este estudio pudieron ser: la cantidad pequeña de datos, el uso de solo animales nacidos vivos, la falta de exactitud en los intervalos de peso registrados (0.5 kg), y la posible varianza de dominancia presente por la alta proporción de hermanos completos. Por lo anterior, para proponer conclusiones definitivas al respecto, se requiere realizar análisis posteriores con mayor cantidad y calidad de información.
Los valores estimados para PD de este estudio fueron similares a los publicados en otras poblaciones de ciervos. En fallow deer se estimó una heredabilidad de 0.29 ± 0.19(11), McManus y Hamilton(2) publicaron un valor estimado de 0.21 ± 0.06, y McManus(7) publicó valores que oscilaron entre 0.22 y 0.89. Para una población de ciervo rojo en México, se estimaron 0.15 ± 0.06 y 0.14 ± 0.11 para heredabilidades directa y materna del PD(4). La correlación entre efectos genéticos directo y materno obtenida en el presente estudio (alta y negativa) fue similar a las publicadas para otras poblaciones de ciervos. En fallow deer se estimó una correlación de -0.64(11), mientras que en ciervo rojo se publicó un valor de -0.92 ± 0.11 (4). Esto tiene fuertes implicaciones para el progreso genético del PD, ya que parte de la ganancia en el crecimiento de los cervatos podría comprometerse por pérdidas en el componente genético materno de las madres, y viceversa. Para aminorar este problema se requiere de futura investigación con mayor cantidad y calidad de información. El problema de la estimación de correlaciones genéticas altas y negativas entre efectos genéticos directo y materno del PD también ha sido observado en otras especies de mamíferos; en bovinos para carne de los EE. UU., por ejemplo, dada la falta de claridad en los mecanismos para estimar con precisión estas correlaciones, 13 de 20 asociaciones de criadores utilizan un valor de cero para esta correlación en las evaluaciones genéticas nacionales(24).
Para PA, un rango amplio de valores estimados de heredabilidad en ciervos ha sido publicado. En ciervo rojo se publicó un valor de 0.14 ± 0.06 para peso a los 11 meses de edad(2), lo que es similar al estimado para PA del presente estudio. Sin embargo, una heredabilidad mayor (0.41 ± 0.23) para PA fue estimada en fallow deer(11), y utilizando varios ranchos de ciervo rojo en cautiverio, se publicaron valores estimados que variaron entre 0.37 y 0.45 para peso a los 11 meses de edad(7). Por el contrario, usando información de un rancho de ciervo rojo en cautiverio con genotipos y manejo similares a los de este estudio, se estimó una heredabilidad para PA de cero(4). El rango amplio de valores estimados de heredabilidad para PA, pudiera en parte deberse a las diferentes metodologías usadas en los análisis y la relativamente pequeña cantidad de información en todas las poblaciones estudiadas.
En general, los valores estimados de heredabilidad de magnitud moderada en PD (directa y materna) y PA (directa) sugieren la detección de variabilidad genética aditiva suficiente para implementar un programa de selección en estas características. Sin embargo, estos resultados pudieran considerarse exploratorios, dado los altos errores estándar obtenidos y la estructura deficiente de la información.
La evaluación genética de ciervo rojo mexicano en cautiverio usando el modelo animal, es importante para identificar animales élite que generen progreso genético en la industria dedicada a esta actividad. El éxito de un programa de mejoramiento debe basarse en una visión compartida de los ganaderos que crían ciervos. Además, en esta especie también conviene implementar otras alternativas para ampliar los objetivos de mejoramiento genético al incluir características adicionales, como las relacionadas con la calidad de la carne y la fertilidad de los animales.
]]>AGRADECIMIENTOS
Los autores agradecen al Centro de Enseñanza, Investigación y Extensión en Producción Bovina y Caprina de la UNAM, por facilitar el uso de la información productiva y genealógica utilizada en este estudio; asimismo, al Consejo Nacional de Ciencia y Tecnología por el financiamiento otorgado para los estudios de maestría del segundo autor.
LITERATURA CITADA
1. Fisher MW, Bryant LD. What might be the consequences of adapting wild animals, such as wapiti, to a farm environment? Proc N Z Soc Anim Prod 1993;53:457-460. [ Links ]
2. McManus CM, Hamilton WJ. Estimation of genetic and phenotypic parameters for growth and reproductive traits for red deer on an upland farm. Anim Prod 1991;53:227-235. [ Links ]
3. Van den Berg GHJ, Garrick DJ. Inheritance of adult velvet antler and live weights in farmed red deer. Livest Prod Sci 1997;49:287-295. [ Links ]
4. Delgadillo CAC, Montaldo HH, Berruecos JM, Luna AA, Vásquez PG. Direct and maternal genetic variance components for growth traits in red deer (Cervus elaphus scoticus). Vet Mex 2008;39:237-245. [ Links ]
5. Stevenson JM, Seman DL, Littlejohn RP. Seasonal variation in venison quality of mature, farmed red deer stags in New Zealand. J Anim Sci 1992;70:1389-1396. [ Links ]
6. Amer PR, Nicoll GB, Garrick DJ. The importance of date of birth in genetic evaluation of deer. Proc N Z Soc Anim Prod 1999;59:129-130. [ Links ]
7. McManus C. Within-farm estimates of genetic and phenotypic parameters for growth and reproductive traits for red deer. Anim Prod 1993;57:153-159. [ Links ]
8. Vásquez CG, Olvera L, Siqueiros Y, Kuri ML, Navarro VL, Rovelo AE, Shimada A. Nursing and feeding behaviour of confined red deer (Cervus elaphus scoticus) in the Mexican highlands. N Z J Agr Res 2004;47:1-9. [ Links ]
9. Boldman KG, Kriese LA, Van Vleck LD, Van Tassell CP, Kachman SD. A Manual for Use of MTDFREML. A set of programs to obtain estimates of variances and covariances [DRAFT]. USDA, ARS. 1995. [ Links ]
10. Meyer K. Variance components due to direct and maternal effects for growth traits of Australian beef cattle. Livest Prod Sci 1992;31:179-204. [ Links ]
11. Morris CA, McCall JE, Baker RL, Southey BR. Genetic parameters for live weights in fallow deer (Dama dama L.). Proc N Z Soc Anim Prod 1992;52:133-135. [ Links ]
12. Fisher MW, McLeod BJ, Mockett BG, Moore GH, Drew KR. Reproductive senescence in aged red deer hinds. Proc N Z Soc Anim Prod 1996;53:457-460. [ Links ]
13. Landete-Castillejos T, García A, Gomez JA, Berruga MI, Gallego L. Effects of birth date and order in lactation performance of Iberian red deer (Cervus elaphus hispanicus). J Anim Sci 2005;83:154-158. [ Links ]
14. Fennessy PF, Thompson JM. Biological efficiency for venison production in reed deer. Proc N Z Soc Anim Prod 1989;49:5-10. [ Links ]
15. Landete-Castillejos T, García A, Gallego L. Calf growth in captive Iberian red deer (Cervus elaphus hispanicus): effects of birth date and hind milk production and composition. J Anim Sci 2001;79:1085-1092. [ Links ]
16. García A, Landete-Castillejos T, Molina A, Albiñana B, Fernández C, Garde J, Gallego L. Lactation curves in captive Iberian red deer (Cervus elaphus hispanicus). J Anim Sci 1999;77:3150-3155. [ Links ]
17. Slate J, Visscher PM, MacGregor S, Stevens D, Tate ML, Pemberton JM. A genoma scan for quantitative trait loci in a wild population of red deer (Cervus elaphus). Genetics 2002;162:1863-1873. [ Links ]
18. Wilson AJ, Pemberton JM, Pilkington DW, Coltman DW, Mifsud DV, Clutton-Brock TH, Kruuk LEB. Environmental coupling of selection heritability limits evolution. PLoS Biology 2006;4:1270-1275. [ Links ]
19. Houle D. Comparing evolvability and variability of quantitative traits. Genetics 1992;130:195-204. [ Links ]
20. Williams JD, Krueger WF, Harmel DH. Heritabilities for antler characteristics and body weight in yearling white-tailed deer. Heredity 1994;73:78-83. [ Links ]
21. Kruuk LEB, Clutton-Brock TH, Slate J, Pemberton JM, Brotherstone S. Heritability of fitness in a wild mammal population. PNAS 2000;97:698-703. [ Links ]
22. Kruuk LEB. Estimating genetic parameters in natural populations using the 'animal model'. Phil Trans R Soc Lond B 2004;359:873-890. [ Links ]
23. Wilson AJ, Coltman DW, Pemberton JM, Overall ADJ, Byrne KA, Kruuk LEB. Maternal genetic effects set the potential for evolution in a free-living vertebrate population. J Evol Biol 2005;18:405-414. [ Links ]
24. BIF (Beef Improvement Federation). Uniform Guidelines for beef Improvement Programs. Beef Improvement Federation, Animal and Dairy Science Department, The University of Georgia. Athens, GA, USA. 2002. [ Links ]
]]>