
Incompatibilidad gametofítica en las razas mexicanas de maíz*
Gametophytic incompatibility in Mexican maize breeds
José Miguel Padilla García1§, José de Jesús Sánchez González1, Lino de la Cruz Larios1, José Ariel Ruiz Corral2, José Ron Parra1 y Moisés Martín Morales Rivera1
1 Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA). Universidad de Guadalajara. Carretera Guadalajara-Nogales, km. 15.5. A. P. 129. C. P. 45110. Las Agujas, Nextipac, Zapopan, Jalisco, México. Tel. y Fax.: (33)3682 - 0743. §Autor para correspondencia: (jmpadillagarcia@hotmail.com).
]]> 2 Campo Experimental Centro Altos de Jalisco. Centro de Investigación Regional del Pacífico Centro. Carretera Tepatitlán-Lagos de Moreno, km 8. INIFAP. A. P. Núm. 56. Tepatitlán de Morelos 47600, Jalisco. Tel. 33-36413575 Ext. 114.
* Recibido: septiembre de 2011
Aceptado: enero de 2012
Resumen
La incompatibilidad esporofítica y gametofítica evitan la autofecundación o la fecundación cruzada, y es una forma de aislamiento reproductivo. Los genes de incompatibilidad-cruzada en maíz de los loci ga1 y tcb1, son causantes del reconocimiento o rechazo entre el estigma y el polen, y de ellos depende la presencia o ausencia de grano en la mazorca del maíz. El presente estudio tuvo como objetivo detectar alelos de incompatibilidad-cruzada de ga1 y tcb1 en colecciones representativas de grupos raciales de maíz de México. El trabajo se realizó en el Centro Universitario de Ciencias Biológicas y Agropecuarias de la Universidad de Guadalajara, ubicado en Zapopan, Jalisco, México durante los ciclos agrícolas de primavera-verano 2007 a 2009. Se evaluaron, en condiciones de temporal, 184 accesiones de razas mexicanas de maíz con base en porcentaje de llenado de grano de mazorca en sus cruzas con probadores macho recesivos (sin factores de incompatibilidad) y probadores hembra dominantes (con factores de incompatibilidad). En el análisis de varianza hubo diferencias altamente significativas tanto entre razas como entre accesiones dentro de razas para el locus ga1. Con relación al locus tcb1 sólo se encontraron diferencias altamente significativas entre razas. El alelo Ga1-S, responsable de la incompatibilidad no-recíproca, se detectó 53.8% de las accesiones, mientras que el alelo neutro Ga1-m y el alelo recesivo ga1 se identificaron 55.43 y 15.76%, respectivamente. El alelo Tcb1-S fue detectado en muy bajas proporciones, concentrado principalmente en las razas de Elotes-Occidentales y Maíz-Dulce.
Palabras clave: Zea mays L., factores gametofíticos, flujo genético, incompatibilidad-cruzada.
Abstract
]]> Sporophytic and gametophytic incompatibility avoids self-fecundation or cross-fertilization, and it is a form of reproductive isolation. The cross-incompatibility genes in maize from loci ga1 and tcb1, cause recognition or reject between stigma and pollen, and from them depends presence or absence of grain in maize ear. The aim of this study was to detect alleles of cross-incompatibility of ga1 and tcb1 in collections representing maize breeds groups from Mexico. The work was made at Centro Universitario de Ciencias Biológicas y Agropecuarias from Universidad de Guadalajara, located in Zapopan, Jalisco, Mexico during agricultural spring-summer 2007 to 2009 cycles. Under rainfall conditions 184 accessions of Mexican maize breeds were assessed based in percentage of grain filling in ear in their crosses with recessive male testers (without incompatibility factors) and dominant female testers (with incompatibility factors). The analysis of variance suggested highly significant differences as well between breeds as between accessions within breeds for locus ga1. With regards locus tcb1 there were found only highly significant differences between breeds. Allele Ga1-S, responsible of non-reciprocal incompatibility, was detected in 53.8% of accessions, while neutral allele Ga1-m and recessive allele ga1 was identified in 55.43 and 15.76%, respectively. Allele Tcb1-S was detected at very low rates, mainly found in breeds Elotes-Occidentales and Maíz-Dulce.Key words: Zea mays L., gametophytic factors, genetic flow, cross-incompatibility.
Introducción
En especies alógamas una característica esencial de la autoincompatibilidad es evitar la autofecundación, como un medio para reducir los efectos deletéreos de la endogamia (Castric y Vekemans, 2004; Pickup y Young, 2008). Asimismo, la incompatibilidad-cruzada causa restricción del flujo de genes, aislamiento reproductivo y la prevención de híbridos inadaptados (Matsubara et al., 2003; Kermicle et al., 2006). Se conocen dos tipos de incompatibilidad, el tipo heteromórfico, que se caracteriza por las diferencias morfológicas entre genotipos. Por ejemplo las diferencias en la longitud o la forma de los estilos, y el tipo homomórfico, en donde los genotipos incompatibles no se distinguen morfológicamente; éste último también se clasifica en gametofítico y esporofítico. En el sistema gametofítico, el fenotipo del polen es codificado por su propio genoma haploide, mientras que en el sistema esporofítico, el fenotipo del polen está determinado por el esporofito (de los padres diploides del polen) y puede involucrar interacciones de dominancia entre los alelos (Glémin et al., 2005). En la mayoría de los sistemas de incompatibilidad están presentes factores adicionales, referidos como modificadores, porque influyen en la respuesta de dichos sistemas; la especificidad de la incompatibilidad sólo puede determinarse bioquímicamente (Cruz-García et al., 2003).
Los factores de incompatibilidad-cruzada en el género Zea causantes del reconocimiento o rechazo entre el estigma y el polen son heredados de manera mendeliana simple. Nelson (1996) identificó diez regiones cromosómicas influidas por estos factores. El locus ga1 ha sido muy estudiado, pues el alelo Ga1-S causa esterilidad no-recíproca; una planta Ga1-S/Ga1-S no es receptiva al polen ga1; sin embargo, el polen Ga1-S induce un llenado de grano completo en la mazorca de plantas homocigotas ga1 / ga1. Un tercer alelo, Ga1-m, desde el punto de vista de la compatibilidad, se ha definido como alelo de acción neutral dado que fertiliza todos los genotipos incluyendo a Ga1 -S/ Ga1 -S y las plantas homocigotas para este alelo son receptivas al polen con cualquiera de los tres alelos del locus ga1 (Kermicle y Allen 1990). Kermicle y Allen (1990) encontraron un factor genético que limita el cruzamiento del maíz con el teocintle (Zea spp.) y lo denominaron Complejo de Incompatibilidad del Teocintle (TIC); posteriormente, Evans y Kermicle (2001) y Kermicle y Evans (2005) lo denominaron como el locus teosinte crossing barrier-1 (tcb1). Los mismos autores en 2010 identificaron el alelo de incompatibilidad Ga2-S encontrado predominantemente en poblaciones de teocintle. La acción génica del alelo Ga1-S es parcialmente dominante sobre el alelo ga1, mientras que el alelo Tcb1-S es completamente dominante sobre tcb1 (Kermicle y Allen, 1990; De la Cruz et al., 2008b).
El maíz tiene su centro de origen y diversidad genética en México (Wellhausen et al., 1952; Sánchez et al., 2000), y ha sido ampliamente estudiado en aspectos evolutivos, de diversidad y flujo genético. Los estudios incluyen caracteres morfológicos, fisiológicos (Sánchez et al., 1993), citológicos, bioquímicos, análisis de izoenzimas (Sánchez et al., 2000), moleculares (Matsuoka et al., 2002; Rice et al., 2006), adaptación climática y descriptores ecológicos (Ruíz et al., 2008). Sin embargo, en México, el estudio de los sistemas de incompatibilidad-cruzada en el género Zea es escaso sobresaliendo el estudio de Cíntora (1963) realizado en algunas razas Mexicanas de maíz, y los trabajos de De la Cruz (2007) y De la Cruz et al. (2007 y 2008a) en razas de maíz, teocintle y en híbridos comerciales de maíz.
Se considera que el uso de sistemas de incompatibilidad entre especies o variedades es uno de los mecanismos más efectivos que podrían utilizar los agricultores que producen maíces para usos especiales como: maíz dulce, palomero, de alta lisina, y maíz orgánico libre de organismos genéticamente modificados (OGM); ya que previenen la hibridación indiscriminada del maíz comercial de siembras adyacentes y de plantas voluntarias (Palaudelmàs et al., 2009) y evita el flujo genético no deseado. Asimismo, en regiones de México en donde todavía se encuentran variedades nativas de maíz y poblaciones de teocintle se requiere obtener información de su constitución genética, respecto a la incompatibilidad, con el propósito de tener una buena planeación en los programas de monitoreo y de conservación in situ. El objeto de la presente investigación fue detectar la presencia y distribución geográfica de alelos de incompatibilidad-cruzada gametofítica de los loci ga1 y tcb1 en colecciones representativas de las razas de maíz de México.
Materiales y métodos
]]> Sitio de estudio y manejo de los materialesEl estudio se efectúo en los ciclos agrícolas de primavera-verano 2007 a 2009 en condiciones de temporal, en el campo experimental del Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA) de la Universidad de Guadalajara, ubicado en Nextipac, Zapopan, Jalisco, México, a 20° 44' 42. 5" latitud norte y 103° 30' 52.5" longitud oeste, con una altitud de 1 650 m, temperatura media de 23.6 °C en los meses de junio-octubre y precipitación media anual de 816 mm.
Material genético
Para el estudio del locus ga1 se sembraron 184 accesiones y para el locus tcb1 122, todas representativas de 54 razas mexicanas de maíz (Cuadro 1). Nótese en el Cuadro 1, que para ciertas razas el número de accesiones fue mayor que en otras debido a que en las razas donde se habían detectado los genes de incompatibilidad (Cíntora, 1963; De La Cruz, 2007) se amplió el muestreo dentro de ellas. Adicionalmente, se usaron los probadores hembra NC-348xW22-P4830 y NC-348xW22-P5270, para evaluar el locus ga1, ambos son homocigotes dominantes (Ga1-S/Ga1-S), y fueron identificados por De la Cruz et al. (2008b). En el caso del locus tcb1 se usó la cruza simple LUG03xW22-TIC-1.1 en estado heterocigote (Tcb1-S/-). Además se utilizaron los probadores macho G-8327A y P-32R21 sin factores de incompatibilidad (ga1/ga1, tcb1/tcb1), seleccionados a partir del estudio realizado por De la Cruz et al. (2008a).
Variables de estudio
La variable estudiada fue el llenado de grano de la mazorca, que se cuantificó con la presencia o ausencia de grano de los cruzamientos de las razas de maíz con cada tipo de probador, y en las plantas individuales de cada cruza se estimó el porcentaje del llenado de grano, obtenido con tres probadores aplicando la metodología propuesta por Kermicle y Allen (1990). La estimación del llenado de grano se hizo visualmente redondeando con aproximación a 10%. Esto es, llenado abajo 5% se designó 0%, de 5 a 15% el valor fue 10% y así sucesivamente hasta alcanzar 95% al cual se asignó 100%.
Diseño y manejo de experimentos
Las accesiones se sembraron en el campo con base en su clasificación racial en dos surcos de 5 m de longitud y 0.77 de ancho con 17 plantas por surco. El manejo agronómico fue el recomendado por el INIFAP para esta zona. Se llevó a cabo un análisis de varianza en donde se consideraron como fuentes de variación a las razas y a las accesiones dentro de razas. En los casos en que se detectaron diferencias significativas, se utilizó la prueba de medias de Dunnett al 0.05 de probabilidad. En ambos análisis se utilizó el procedimiento GLM del paquete estadístico SAS versión 8.2 (SAS Institute, 1992). Los testigos de comparación para la prueba de Dunnett fueron tomados de los mismos materiales incluidos en el estudio. El procedimiento GLM calculó para cada comparación el valor de Dunnett debido a que hubo diferencias entre razas para el número de accesiones. Para el caso del probador macho recesivo sin factores de incompatibilidad, se eligió un material con valores de 100% de llenado de grano en mazorca, mientras para el caso de probadores hembra dominantes con factores de incompatibilidad se usaron materiales con valores de 0% de llenado de grano.
Los probadores hembra dominante del locus ga1 (Ga1 -S/ Ga1-S) y del locus tcb1 (Tcb1-S/tcb1), recibieron polen de cada accesión bajo prueba, y cada accesión recibió polen del probador macho recesivo (ga1/ga1, tcb1 /tcb1). Las plantas del probador hembra con factores de incompatibilidad y las plantas de las accesiones se jilotearon antes de la emergencia de los estigmas. Las espigas de los probadores macho y de las accesiones se cubrieron el día anterior a la polinización; las polinizaciones se realizaron manualmente entre las 12:00 y 14:00 h, horario en que el polen es viable en Nextipac, Zapopan, Jalisco (Rodríguez et al., 2006). El diseño fue de bloques completos al azar con una repetición. Los dos primeros ciclos fueron evaluados 184 accesiones con el locus ga1 y en el último ciclo 122 accesiones con el locus tcb1. Los probadores hembra se sembraron en tres fechas de siembra, y los probadores macho tuvieron hasta cinco fechas de siembra para lograr la coincidencia con la floración de las accesiones en estudio.
Criterios para detectar los alelos de incompatibilidad
]]> Para detectar los alelos de los loci ga1 y tcb1 se utilizaron los promedios de llenado de grano en mazorca con ambos tipos de probadores, macho recesivo y hembra dominante. Se ejecutaron tres etapas:Etapa 1. Probador macho recesivo (sin alelos de incompatibilidad): cuando la accesión se polinizó con el probador macho recesivo y no hubo llenado de grano y además, los valores determinados con la prueba de Dunnett fueron estadísticamente iguales al control (0% de llenado de grano en la mazorca), se identificó la presencia de algún alelo de incompatibilidad (Ga1-S, Tcb1-S). En el caso de formación de grano en la mazorca y cuando la prueba de Dunnett señaló valores estadísticamente iguales al control, con 100% de llenado de grano en la mazorca, se identificó la presencia de los alelos neutros (Ga1-m, Tcb1-m) o recesivos (ga1, tcb1), y se continuó con la etapa 2.
Etapa 2. Cruzamientos al probador hembra dominante Ga1 -S/ Ga1-S: una vez excluidas las accesiones con presencia de los alelos Ga1-S o Tcb1-S, el siguiente paso fue llevar polen del material seleccionado a los probadores hembra dominantes. Cuando en el material de prueba se observó 100% de llenado de grano y fue estadísticamente igual al control se identificó la presencia del alelo Ga1-m. Si el llenado de grano fue 0% y fue estadísticamente igual al control, se interpretó como accesión sin alelos de incompatibilidad.
Etapa 3. Cruzamientos al probador hembra heterocigote dominante (Tcb1-S/-): sin considerar las accesiones con alelos Ga1-S o Tcb1-S de la Etapa 1, entonces si el llenado de grano fue 0% y fue estadísticamente igual al control, se identificó la presencia del alelo recesivo tcb1. En cambio, si el llenado de grano fue 100% y fue estadísticamente igual al control, se identificó la presencia del alelo Tcb1-m.
Resultados y discusión
En el análisis de variación de la variable llenado de grano, se obtuvieron los resultados siguientes: en las cruzas con los probadores recesivos (ga1 /ga1, tcb1 /tcb1) y los probadores hembra dominantes (Ga1 -S/Ga1 -S) hubo diferencias altamente significativas tanto entre razas como entre accesiones dentro de razas; mientras que en las cruzas con el probador hembra Tcb1-S/- las diferencias sólo fueron significativas entre razas (Cuadro 2). Los valores altos del coeficiente de variación para el caso del locus tcb1 son debidos a que los porcentajes de llenado de grano son predominantemente ceros, con pocos casos cercanos a 50% (Cuadro 3). Los coeficientes de determinación fueron superiores a 0.8, valores que son considerados como reflejo de un modelo apropiado.
El promedio general de llenado de grano en las razas de maíz con los probadores macho recesivo ga1 /ga1, tcb1/ tcb1 y hembra dominante Ga1-S/Ga1-S fueron altos, del orden de 85 y 72% respectivamente (Cuadro 3), mientras que las cruzas con el probador hembra Tcb1-S/- el promedio general fue muy bajo (6%). En las cruzas de las razas de maíz con el probador hembra Ga1-S/Ga1-S, muy pocas razas tuvieron promedios de llenado de grano en mazorca menores que 20%, entre ellas Azul, Bofo (Coreño), Cristalino de Chihuahua, Gordo, Harinoso de Ocho, Jala, Ratón, Tabloncillo, Tuxpeño Norteño y Zamorano Amarillo, mientras que en cruzas con el probador hembra Tcb1-S/-, sólo las razas Elotes Occidentales y Maíz Dulce, tuvieron promedios de llenado de grano mayores que 20%.
Con base en los promedios de llenado de grano del Cuadro 3 y las pruebas de Dunnett, se determinó la presencia de ambos sistemas de incompatibilidad (Ga1-S y Tcb1-S) en seis razas de maíz: Bolita, Dulcillo del Noroeste, Elotes Occidentales, Maíz Dulce, Negrito y Reventador (Cuadro 3), y en el resto de las razas se detectaron cualquiera de las tres variantes de los alelos del locus ga1.
La presencia proporcional del alelo Ga1-S, responsable de la incompatibilidad no-recíproca, fue 53.8% de las 184 accesiones evaluadas de las 54 razas; en el alelo neutro Ga1-m, compatible con cualquier genotipo del locus ga1; 55.43% de las accesiones, y 15.76% con el alelo ga1 (Cuadro 4). Sólo dos razas en casi la totalidad de sus accesiones, Elotes Occidentales y Maíz Dulce, contienen el alelo dominante en estado homocigoto (Ga1 -S), junto con otras cinco razas en al menos una de sus accesiones Bofo, Chapalote, Dulcillo del Noroeste, Palomero de Chihuahua y Reventador. En ocho razas, todas las accesiones fueron homocigotas recesivas (sin alelos de incompatibilidad) y fueron: Azul, Bofo (Coreño), Cristalino de Chihuahua, Gordo, Ratón, Tabloncillo, Tuxpeño Norteño y Zamorano Amarillo; además en otras tres razas fueron homocigotas recesivas en al menos una de sus accesiones Jala, Palomero de Chihuahua y Reventador (Cuadro 3). De la Cruz (2007) reportó en 71 accesiones que 48% fueron Ga1-m/Ga1-m; 18% recesivas y sólo 21% con el alelo Ga1-S, además en estado heterocigoto Ga1-m/ga1 con 13% contra 6.52% del presente estudio (Cuadro 4). Cíntora (1963) de 229 accesiones estudiadas, informó que 74% para Ga (probablemente Ga1-m/Ga1-m); 17% recesivas ga1/ga1 y sólo 3% para Ga1-S/Ga1-S. Los resultados obtenidos en esta investigación muestran que en las razas Mexicanas de maíz existe una alta presencia del alelo neutro Ga1-m, el cual rompe el aislamiento reproductivo de aquellas poblaciones de maíz homocigotas para el alelo Ga1-S.
]]>
El alelo dominante Tcb1-S del locus tcb1 estuvo presente en las razas Elotes Occidentales y Maíz Dulce; así como en algunas accesiones de las razas: Bolita, Dulcillo del Noroeste, Negrito y Reventador. El resto de las razas presentó porcentajes menores a 3% de llenado de grano en mazorca para el alelo homocigoto recesivo tcb1 (Cuadro 3).
En estudios anteriores Kermicle y Allen (1990), Kermicle (2006), Evans y Kermicle (2001) y Kermicle y Evans (2005) informaron la presencia de factores genéticos que limitaban el cruzamiento del maíz con el teocintle y lo denominaron Complejo de Incompatibilidad del Teocintle (TIC), responsable de la incompatibilidad no-recíproca. Asimismo, indicaron que TIC es único en teocintle y "podría estar jugando un papel muy importante en el aislamiento reproductivo entre el maíz y el teocintle". Con base en el muestreo amplio de accesiones de maíz de todo México y la utilización de probadores apropiados, el sistema de incompatibilidad TIC se encontró en las razas de maíz: Bolita, Dulcillo del Noroeste, Elotes Occidentales, Maíz Dulce, Negrito y Reventador.
Las bases bioquímicas y genéticas para los sistemas de incompatibilidad en el género Zea son aún muy poco conocidas; no hay claridad si son genes individuales, o grupos de genes estrechamente ligados, e incluso existe la idea de que la fecundidad del grano de polen se ve afectada por genes expresados por ambos sistemas, tanto esporofítico como gametofítico (Bedinger y Fowler, 2009). Evans y Kermicle (2001) y Kermicle y Evans (2005) reportan que los genes de incompatibilidad conocidos en maíz y teocintle actúan independientemente y que la sola existencia de un alelo incompatible es suficiente para bloquear la fertilización, lo cual concuerda con este estudio, en que se las accesiones de maíz reaccionaron al reconocimiento o rechazo del grano de polen, este reacción depende del genotipo en estudio y del los alelos del probador macho recesivo (ga1/ga1, tcb1/tcb1). Kermicle et al. (2006) señalaron que los factores gametofíticos de la incompatibilidad, además de ser agentes de aislamiento reproductivo, previenen la formación de híbridos mal adaptados y, además, mantienen la identidad de las razas, específicamente las razas Elotes Occidentales y Maíz Dulce de este estudio. Kermicle (2006) señaló que la presencia del gene Tcb1-S, con potencial de dispersión y fijación, podría añadir diversidad genética en las poblaciones de maíces locales, que concuerdan con los resultados del estudio reflejada en la variación existente entre razas como entre accesiones dentro de razas.
Los sistemas de incompatibilidad presentan un patrón definido dentro de los complejos raciales del maíz, especialmente en las razas Elotes Occidentales y Maíz Dulce (Figura 1); el origen de este sistema de incompatibilidad pudiera ser diferente al complejo de incompatibilidad del teocintle (TIC); sin embargo, se requerirán pruebas de alelismo detalladas y ubicación y secuencia de las regiones del genoma involucradas con dicho sistema a fin de considerarlo como de novo.
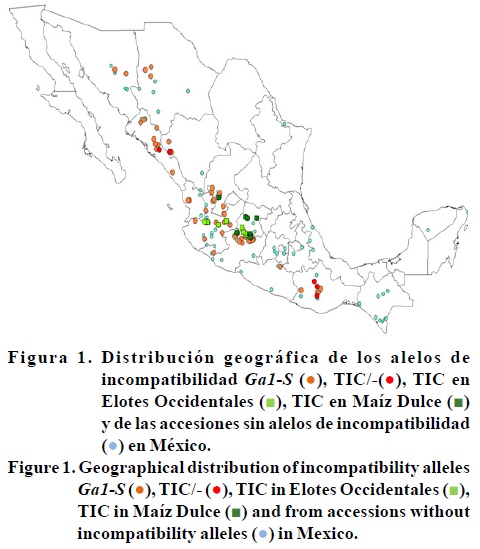
Las razas mexicanas que presentaron en la misma accesión los alelos Tcb1-S como Ga1-S fueron: Dulcillo del Noroeste, Elotes Occidentales, Maíz Dulce y Reventador. Kermicle et al. (2006) señalaron que es posible que el alelo Ga1-S, responsable de la incompatibilidad no-recíproca, sea en si un modificador para el fortalecimiento de otros sistemas de barrera de incompatibilidad como el locus tcb1 de manera independiente o de forma interactiva, faltan estudios adicionales para sustentar si el alelo Ga1-S per se, un alelo modificador del alelo Tcb1-S, ya que los probadores de ambos locus del presente estudio fueron trabajados en forma separada y se obtuvo respuestas individuales por locus probado.
]]> Conclusiones
Los genes responsables de la incompatibilidad no recíproca Ga1-S y Tcb1-S fueron encontrados en seis de las razas mexicanas de maíz evaluadas con ambos sistemas de incompatibilidad en al menos una accesión de maíz (Bolita, Dulcillo del Noroeste, Elotes Occidentales, Maíz Dulce, Negrito y Reventador), pero debido a la presencia en altas proporciones del alelo neutro Ga1-m, las barreras existentes en las razas de maíz y del teocintle basadas únicamente en el alelo Ga1-Spueden no ser suficientes para evitar el flujo genético no deseado.
Se identificaron 16 accesiones de maíz de las razas Bolita, Dulcillo del Noroeste, Elotes Occidentales, Maíz Dulce, Negrito y Reventador, con frecuencias altas del alelo Tcb1-S; las cuales se localizan especialmente en el Occidente de México; resultados que demuestran que el sistema TIC no está presente sólo en el teocintle de México.
Literatura citada
Bedinger, P. A. and Fowler, J. E. 2009. The maize male gametophyte. In: handbook of maize: its biology. Bennetzen, J. and Hake, S. (eds.). Springer. 57-75 pp. [ Links ]
Castric, V. and Vekemans, X. 2004. Plant self-incompatibility in natural populations: a critical assessment of recent theoretical and empirical advances. Mol. Ecol. 13:2873-2889. [ Links ]
Cíntora, O. C. 1963. Incompatibilidad no-recíproca en algunas colecciones mexicanas de maíz. Tesis de Maestría en Ciencias. Facultad del Colegio de Graduados de la E. N. A. Chapingo, Estado de México. 39 p. [ Links ]
Cruz-García F.; Hancock, C. N. and McClure, B. 2003. S-RNase complexes and pollen rejection. J. Exp. Bot. 54:123-130. [ Links ]
De la Cruz, L. L. 2007. Sistemas de incompatibilidad genética en maíz y teocintle (Zea spp.) en México. Tesis de Doctorado. Centro Universitario de Ciencias Biológicas y Agropecuarias. Universidad de Guadalajara. México. 81 p. [ Links ]
De la Cruz, L. L.; Sánchez, G. J. J.; Rodríguez, G. E.; Ron, P. J. y Santacruz, R. F. 2007. El factor gametofítico-1 (ga1) en Teocintle (Zea spp.) en México. Scientia- Cucba 9:129-141. [ Links ]
De la Cruz, L. L.; Sánchez, G. J. J.; Ron, P. J.; Santacruz, R. F.; Baltazar, M. B.; Ruíz, C. J. A. y Morales, R. M. M. 2008a. El factor gametofítico-1 (ga1) en híbridos comerciales de maíz en México. Rev. Fitotec. Mex. 31:57-65. [ Links ]
De la Cruz, L. L.; Sánchez, G. J. J.; Ron, P. J.; Santacruz, R. F.; Rodríguez, G. E.; Ruíz, C. J. A. y Morales, R. M. M. 2008b. Probadores de maíz para factores de incompatibilidad gametofítica. Rev. Fitotec. Mex. 31:341-349. [ Links ]
Evans, M. M. S. and Kermicle, J. L. 2001. Teosinte crossing barrier-1, a locus governing hybridization of teosinte with maize. Theor Appl. Genet. 103:259-265. [ Links ]
Glémin, S.; Gaude, T.; Guillemin, M. L.; Lourmas, M.; Olivieri, I. and Mignot, A. 2005. Balancing selection in the wild: testing population genetics theory of self-incompatibility in the rare species Brassica insularis. Genetics 171:279-289. [ Links ]
Kermicle, J. L. and Allen, J. O.1990. Cross-incompatibility between maize and teosinte. Maydica 35:399- 408. [ Links ]
Kermicle, J. L. and Evans, M. M. S. 2005. Pollen-pistil barriers to crossing in maize and teosinte result from incongruity rather than active rejection. Sex Plant Reprod 18:187-194. [ Links ]
Kermicle, J. L. 2006. A selfish gene governing pollen-pistil compatibility confers reproductive isolation between maize relatives. Genetics 172:499-506. [ Links ]
Kermicle, J. L.; Taba, S. and Evans, M. M. S. 2006. The gametophyte-1 locus and reproductive isolation among Zea mays subspecies. Maydica 51:219-226. [ Links ]
Kermicle, J. L. and Evans, M. M. S. 2010. The Zea mays sexual compatibility gene ga2: naturally occurring alleles, their distribution, and role in reproductive isolation. J. Heredity 101:1-13. [ Links ]
Matsuoka, Y.; Vigourous, Y.; Goodman, M. M.; Sánchez, G. J.; Buckler, E. S. and Doebley, J. F. 2002. A single domestication for maize shown by multilocus microsatellite genotyping. Proceedings of the National Academy of Sciences 99:6080-6084. [ Links ]
Matsubara, K.; Thidar, K. and Sano, Y. 2003. A gene block causing cross-incompatibility hidden in wild and cultivated rice. Genetics 165:343-352. [ Links ]
Nelson, O. E. 1996. The gametophyte factors of maize. In: the maize handbook. Freeling, M. and Walbot, M. (Eds.). Springer-Verlag. New York/Berlín. 496-453 pp. [ Links ]
Palaudelmàs, M.; Peñas, G.; Melé, E.; Serra, J.; Salvia, J.; Pla, M.; Nadal, A. and Messeguer, J. 2009. Effect of volunteers on maize gene flow. Transgenic Research 18:583-94. [ Links ]
Pickup, M. and Young, A. G. 2008. Population size, self-incompatibility and genetic rescue in diploid and tetraploid races of Rutidosis leptorrhynchoides (Asteraceae). Heredity 100:268-274. [ Links ]
Rice, E. B.; Smith, M. E.; Mitchell, S. E. and Kresovich, S. 2006. Conservation and change: a comparison of In situ and ex situ conservation of Jala maize germplasm. Crop Sci. 46:428-436. [ Links ]
Rodríguez, F. J. G.; Sánchez, G. J. J.; Baltazar, B. M.; De la Cruz, L. L.; Santacruz, R. F.; Ron, P. J.; Schoper, J. B. 2006. Characterization of floral morphology and synchrony among Zea species in Mexico. Maydica 51:383-398. [ Links ]
Ruiz, C. J. A.; Durán, P. N.; Sánchez, G. J. J.; Ron, P. J.; González, E. D. R.; Holland, J. B. and Medina, G. G. 2008. Climatic adaptation and ecological descriptors of 42 Mexican maize races. Crop Sci 48:1502-1512. [ Links ]
Sánchez, G. J. J.; Goodman, M. M. and Rawlings, J. O.1993. Appropriate characters for racial classification in maize. Econ. Bot. 47:44-59. [ Links ]
Sánchez, G. J. J.; Goodman, M. M. and Stuber, C. W. 2000. Isozymatic and morphological diversity in the races of maize of Mexico. Econ. Bot. 54:43-59. [ Links ]
Statistical Analysis System (SAS Institute Inc). 1992. SAS/ STAT software: changes and enhancements. SAS Institute Inc., Cary, N. C: SAS Institute Inc. [ Links ]
Wellhausen, E. J.; Roberts, L. M.; Hernández, X. E. and Mangelsdorf, P. C. 1952. Races of maize in Mexico: their origin, characteristics and distribution. The Bussey Institution of Harvard University. 223 p. [ Links ]
]]>