Vías de señalización asociadas a la esteroidogénesis
Signaling pathways associated to steroidogenesis
Erika Gómez-Chang1, Fernando Larrea2 y Federico Martínez-Montes1
1Depto. de Bioquímica, Facultad de Medicina, UNAM. Ciudad Universitaria, C.P. 04510, Delegación Coyoacán, México, D.F. Correo: fedem@bq.unam.mx
]]> 2Depto. de Biología de la Reproducción, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán. México, D.F.
Artículo recibido el 08 de febrero de 2012
Aceptado el 12 de junio de 2012
Resumen
La biosíntesis de las hormonas esteroides es necesaria para mantener la función reproductiva y la homeostasis en general. Este proceso que se lleva a cabo en tejidos esteroidogénicos regulados en forma aguda o crónica, activa vías de señalización mediadas particularmente por cinasas de proteínas dependientes del adenosín monofosfato cíclico/AMPc (PKA). Datos obtenidos en placenta humana por nuestro grupo de trabajo y en otros tejidos informados en la literatura, muestran que la fosforilación dinámica de proteínas catalizada por PKA, está asociada a la esteroidogénesis, incluyendo su regulación hormonal. sin embargo, se conoce la participación de otras cascadas de transducción de señales independientes de PKA y que son activadas por hormonas tróficas, citocinas o factores de crecimiento, así como por iones de cloruro y calcio e inclusive por metabolitos del ácido araquidónico. El estudio y comprensión de los factores estimulantes, así como de los mecanismos que regulan la síntesis de las hormonas esteroides, permitirá identificar los sitios potenciales de intervención y de esta manera coadyuvar al mantenimiento de la homeostasis y de las diferentes funciones dependientes de los mismos.
Palabras Clave: Cinasa de proteínas dependiente de AMPc, esteroidogénesis, fosforilación de proteínas, regulación aguda y crónica, transducción de señales.
Abstract
]]> The biosynthesis of steroid hormones is necessary to maintain reproductive functions and body homeostasis. This process, which is carried out in acutely and chronically regulated steroidogenic tissues, depends on the activation of signaling pathways particularly mediated by the cyclic adenosine monophosphate dependent protein kinase (PKA). Data obtained by our and other groups in human placenta and other tissues, have clearly showed that dynamic phosphorylation catalyzed by PKA is related to steroidogenesis, including its hormonal regulation. it is known, however, that this process also involves the participation of other PKA independent signaling cascades, which are activated by trophic hormones, cytokines or growth factors and others such as calcium, chloride ions and arachidonic acid metabolites. The understanding of the stimulating factors, as well as the mechanisms that regulate the synthesis of steroid hormones, will allow us the identification of potential intervention sites that may contribute to maintain body homeostasis and different functions depending on them.Key Words: Cyclic adenosine monophosphate dependent protein kinase, steroidogenesis, protein phosphorylation, acute and chronic regulation, signal transduction.
Introducción
Las hormonas esteroides se clasifican en esteroides sexuales (andrógenos, estrógenos y progesterona), glucocorticoides (cortisol) y mineralocorticoides (aldosterona) y se sintetizan principalmente en las glándulas suprarrenales, las gónadas, la placenta y el sistema nervioso central. Su principal efecto es sobre la homeostasis corporal al modular diversas funciones como son la reproductiva, incluyendo el desarrollo y mantenimiento de los caracteres sexuales secundarios, entre otros. La esteroidogénesis puede estar sujeta a una regulación a corto o largo plazo dependiendo del tejido. En las gónadas y en las glándulas suprarrenales la regulación es aguda, en tanto que en el cerebro y en la placenta es crónica. Los dos tipos de regulación pueden estar bajo el control de diversos factores u hormonas, que en el caso de la respuesta aguda inicia en el orden de minutos e implica la movilización del colesterol hacia la mitocondria[1] y su translocación de la membrana mitocondrial externa hacia la interna, donde ocurre la síntesis de pregnenolona por acción de la enzima citocromo P450scc (CYP11A1; por sus siglas en inglés Cholesterol side ccleavage enzyme).
La cascada de señalización más estudiada en la esteroidogénesis es la mediada por la cinasa de proteínas dependiente del adenosín monofosfato cíclico/AMPc (PKA). El efecto agudo de las hormonas tróficas incrementa el contenido intracelular de AMPc activando a la PKA, la cual fosforila proteínas involucradas en el transporte de colesterol hacia la mitocondria, como es el caso de la proteína StAR (Steroidogenic Acute Regulatory protein) de 30 kDa, cuya expresión aumenta por el efecto hormonal que promueve la esteroidogénesis en gónadas y corteza suprarrenal[2]. La respuesta crónica por el estímulo de hormonas tróficas se inicia en el transcurso de horas e involucra también la participación de PKA a través de la fosforilación de factores de transcripción relacionados con la activación de genes que codifican para enzimas esteroidogénicas, además de la proteína StAR[3]. La vía mediada por PKA/AMPc es la principal cascada estimulada en los diferentes tejidos esteroidogénicos; sin embargo, hay evidencias de la participación de otras vías de señalización activadas tanto por hormonas como por factores derivados de macrófagos y factores de crecimiento que actúan a través de PKA o bien a través de otras vías independientes de AMPc y que pueden estar mediadas por la proteína cinasa dependiente de calmodulina (PKC), iones de cloruro y calcio, metabolitos derivados del ácido araquidónico o proteínas cinasas activadas por mitógeno (MAPK). Cabe mencionar que las respuestas generadas independientemente de las concentraciones de AMPc, suelen ser poco potentes comparadas con las que son mediadas por PKA/AMPc, por lo que su efecto se relaciona con la modulación o potenciación de la esteroidogénesis actuando en forma sinérgica con PKA. Cualquiera que sea la cascada de señalización activada, destaca la importancia de los eventos de fosforilación o desfosforilación de sustratos específicos por proteínas cinasas y fosfatasas, respectivamente, con efectos pleiotrópicos sobre diferentes funciones celulares como son el metabolismo, crecimiento, diferenciación y la esteroidogénesis[4-6].
El objetivo de esta revisión es mostrar un panorama general sobre el proceso esteroidogénico en los tejidos que están sujetos a regulación aguda y crónica, así como las principales vías de señalización que están involucradas.
Con ello se pretende hacer énfasis en las diferencias entre y que caracterizan a ambos tipos de regulación. Esta revisión pretende también ofrecer una descripción del proceso de biosíntesis de hormonas esteroides en los tejidos que responden de manera aguda y crónica, así como de las vías de señalización que se activan con el objeto de integrar la información existente con hincapié en la comunicación entre las varias vías o cascadas de transducción y su asociación con los eventos de fosforilación y desfosforilación que ocurren a nivel intracelular.
Regulación de la esteroidogénesis por receptores acoplados a proteínas G
]]> Tanto en forma aguda como crónica, la esteroidogénesis se inicia por el estímulo de mensajeros primarios que pueden ser de naturaleza hormonal. Estos efectos se traducen en eventos de fosforilación o desfosforilación de proteínas e incluso de factores de la transcripción que participan en la regulación diferencial de la esteroidogénesis. Para que ocurra la transducción de la señal, el primer mensajero requiere de su interacción con receptores de siete dominios transmembranales que están acoplados a proteínas Gs. Éstas inducen la activación de la adenilato ciclasa, la cual cataliza la formación de AMPc a partir de ATP. El AMPc, como segundo mensajero, activa a la PKA. Cabe mencionar que también existen otras proteínas como los canales iónicos, fosfodiesterasas, proteínas de intercambio de nucleótidos (EPAC) y proteínas cinasas dependientes de GMPc (PKG), que son activadas por AMPc[7-9]. La cascada de señalización mediada por PKA es universal pero muestra variaciones en cuanto a los factores que la estimulan y los efectos finales obtenidos. Estructuralmente, la PKA es un tetrámero conformado por dos subunidades catalíticas (C) y dos reguladoras (R)[10]. Existen 4 tipos de subunidades reguladoras (RIa, RIIa, RIP y RIIP), todas codificadas por genes separados, así como 3 tipos de subunidades catalíticas (Ca, CP y Cy). El tipo de subunidad reguladora define a la isoforma tipo I de PKA (a y P) y la tipo II (a y P). Las subunidades RIa y RIIa son ubicuas, en tanto que las subunidades RIP y RIIP se expresan predominantemente en cerebro y tejido adiposo entre otros órganos endocrinos. Las isoformas difieren en cuanto a su distribución tisular y celular y en la afinidad por AMPc, sugiriendo que la holoenzima permite decodificar señales dependiendo de la tasa de activación, duración y magnitud del estímulo inicial[11-12]. La importancia de las 2 isoformas de PKA radica en su expresión y localización diferencial que no sólo es tejido dependiente, sino también específica de algunos organelos celulares. En este sentido, destaca la participación de las proteínas AKAP (por sus siglas en inglés Akinase anchoring proteins), una familia de proteínas funcionalmente relacionadas y conformadas por más de 50 miembros, resultado del procesamiento y empalme alternativo del RNAm. Estas familias de proteínas fijan a las subunidades reguladoras de la PKA anclándola hacia una localización subcelular específica. La afinidad con la que PKA interactúa con las AKAP's depende del tipo de la subunidad reguladora. La asociación específica de ciertas AKAP's con las isoformas de PKA constituye un mecanismo de regulación de esta cinasa, asegurando que ésta se ubique cerca del sitio donde se genera el AMPc que la activa y de los sustratos que serán fosforilados.En el caso de la mitocondria, la proteína AKAP-121 fija a la isoforma II de PKA a la membrana mitocondrial externa formando complejos, pero se ha visto que las AKAP's también pueden fijar ARNm e incluso a una fosfatasa modulada por PKA denominada PTPD1 (por sus siglas en inglés protein tyrosine phosphatase)[13,14,15]. El anclaje de cinasas y fosfatasas por las AKAP's sugiere que los eventos de fosforilación y desfosforilación asociados a diversos procesos celulares podrían ser más eficientes al compartamentalizar a las vías de transducción de señales (Figura 1)[13,16].
La principal respuesta de las células esteroidogénicas a los factores hormonales es la activación de receptores acoplados a proteínas G, con la consecuente estimulación de la adenilato ciclasa y el incremento en las concentraciones intracelulares de AMPc. En las gónadas y las glándulas suprarrenales, la respuesta aguda mediada por la vía AMPc-PKA involucra la fosforilación de enzimas como la hidrolasa de ésteres del colesterol o de proteínas que participan en el transporte de colesterol hacia la mitocondria como es el caso de la StAR (Figura 2). Numerosas evidencias han demostrado la importancia de la participación de PKA en el mantenimiento de la tasa de expresión estacionaria de StAR, así como de su actividad a través de la fosforilación de los residuos de Ser194/195 y Ser 56/57 en el humano y el murino, respectivamente[17].
Los fenómenos acoplados de fosforilación/desfosforilación mediados por PKA se han documentado como punto clave de la regulación de procesos como el crecimiento, la migración celular e incluso de cambios morfológicos celulares18,19. Por otro lado, en tejidos de regulación crónica, como la placenta y el sistema nervioso central, los procesos de fosforilación/ desfosforilación ocurren a nivel de los factores de transcripción como el esteroidogénico 1(SF-1), GATA-4 y la proteína fijadora de los elementos de respuesta a AMPc (CREB) e incluso en aquellos que codifican para proteínas asociadas al transporte de colesterol y de enzimas esteroidogénicas (Figura 3)[3].
Vías de señalización independientes de AMPc Participación de PKC
La familia de PKC consiste en múltiples isoenzimas con diversos patrones de distribución en diferentes tejidos. La unión de ligandos a receptores acoplados a proteínas Gq o con actividad de cinasa de tirosinas inducen la activación de la fosfolipasa C con la consecuente producción de IP3 (inositol 1,4,5-trifosfato) y DAG (diacilglicerol). El IP3 provoca la apertura de los canales de Ca2+ del retículo endoplásmico, con lo que se incrementan las concentraciones citosólicas de este ion. El calcio ejerce su acción como segundo mensajero activando a otras cinasas por lo consiguiente se amplifica la señal iniciando así la cascada de fosforilaciones dependientes de PKC. Las isoenzimas de PKC no sólo difieren en su localización, sino también en su capacidad de respuesta inducida por IP3, DAG o calcio. Por otro lado, la actividad de PKC también es regulada por su asociación a proteínas de anclaje, como es el caso de PRKCA-binding protein, que ancla a PKCa a la mitocondria[20].
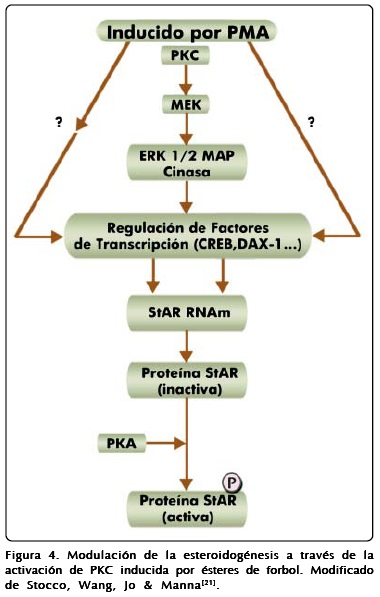
Algunos primeros mensajeros hormonales inducen la vía mediada por PKC, pero los resultados con respecto a la participación de esta cinasa en la esteroidogénesis han sido controversiales, ya que en algunos casos, los eventos de fosforilación se asocian al incremento en la síntesis hormonal, en tanto que en otros, se produce inhibición o simplemente no se observan los efectos. La activación de PKC se ha estudiado empleando PMA, un éster de forbol, que funciona como análogo del DAG; sin embargo, su efecto parece ser principalmente a nivel transcripcional sin alterar directamente la tasa de fosforilación. Por otra parte, la activación de PKC con PMA puede activar cinasas efectoras de MAPK, como ERK1/2, que a su vez pueden fosforilar a proteínas como CREB[21]. Estos datos sugieren que PKC y sus activadores funcionan modulando la esteroidogénesis a través de efectos transcripcionales y postraduccionales de proteínas asociadas a la síntesis de hormonas esteroides (Figura 4)[21].
Participación de las MAPK
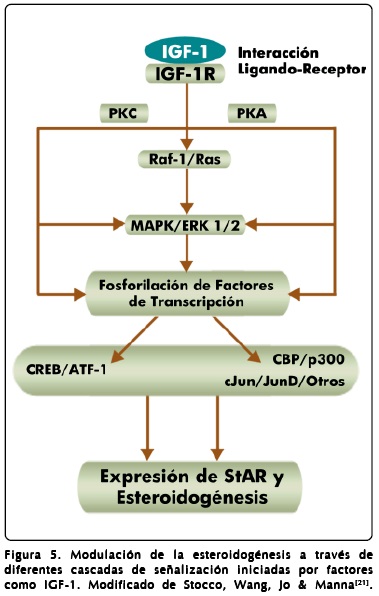
La vía mediada por las MAPK (cinasas de proteínas activadas por mitógeno) participa en la transducción de señales iniciadas por citocinas como IL-1(interleucina 1) y de factores como EGF (factor de crecimiento epidérmico) e IGF-1 (factor de crecimiento semejante a insulina), cuyos receptores tienen actividad intrínseca de tirosina cinasa, lo que desencadena la activación secuencial de otras cinasas efectoras como MEK-ERK1/2. La respuesta esteroidogénica mediada por vías como la de MAPK es moderada (menos del 1%) comparada con la que se induce a través de la vía AMPc/PKA. La función de MAPK implica la fosforilación de diversos factores de transcripción asociados a la biosíntesis de esteroides como CREB, ATF-1 (factor activador de la transcripción 1) y cJun/cJunD, lo que incluso puede resultar en la regulación de la expresión de la proteína StAR. En el caso de las células de Leydig del ratón, la respuesta provocada por la unión de IGF-1 a su receptor, activa a factores de la transcripción mediante la participación de varias cinasas como la PKA, PKC y MAPK (Figura 5)[21].
Participación del Ácido Araquidónico
En las células esteroidogénicas la respuesta aguda inducida por hormonas como la LH (hormona luteinizante) o la hCG (hormona gonadotropina coriónica humana) provoca el aumento en las concentraciones intracelulares de AMPc, así como la mayor liberación de ácido araquidónico por acción de la fosfolipasa A2. La importancia del ácido araquidónico radica en que sus metabolitos participan en la regulación transcripcional y expresión de la StAR. Además de esta vía, en las células suprarrenales, se han descrito otras alternativas para la liberación intracelular del ácido araquidónico a través de la inducción de la acil-CoA tioesterasa mitocondrial, también denominada ARTISt (de sus siglas en inglés AA-rthioesterase involved in steroidogenesis) y la ACS4 (AA-preferring acyl-CoA synthase), ambas enzimas importantes para proveer el ácido araquidónico que se requiere para la expresión de StAR[22,23]. Hasta este momento se ha visto que la principal vía de señalización involucrada en la esteroidogénesis es la que está mediada por PKA/AMPc, aunque dependiendo del tejido, pueden participar otras cascadas de transducción de señales que en general actúan en conjunto con la PKA modulando el proceso biosintético. En la siguiente sección se hará una integración de la forma en la que actúan las vías de señalización mencionadas para inducir la esteroidogénesis en tejidos de regulación aguda y crónica.
Biosíntesis de hormonas esteroides
]]> El proceso de síntesis de mineralocorticoides, glucocorticoides, andrógenos, progestinas y estrógenos a partir del colesterol, se lleva a cabo por diversas enzimas entre las cuales se encuentran diferentes tipos de citocromo P450, deshidrogenasas de esteroides y reductasas. La síntesis de las hormonas esteroides inicia con la conversión del colesterol a pregnenolona por la enzima citocromo P450scc/Cyp11A1 localizada en la membrana interna mitocondrial. La pregnenolona es biotransformada a progesterona por acción del complejo enzimático de la 3P-hidroxiesteroide deshidrogenasa A4-5 isomerasa (3P-HSD). Existen dos isoformas de la 3P-HSD, una de localización mitocondrial y otra en el retículo endoplásmico[24].
Vías de señalización que participan en los tejidos esteroidogénicos Placenta Humana
Una de las funciones de la placenta humana es la síntesis de progesterona, la cual es indispensable para el mantenimiento del embarazo. A diferencia de otros tejidos como la corteza suprarrenal o las gónadas, la placenta no cuenta con regulación aguda de la esteroidogénesis. Los esteroides sintetizados bajo la forma de regulación aguda muestran cambios sustanciales en sus concentraciones plasmáticas en sólo minutos después de la estimulación hormonal. La placenta humana no expresa la proteína StAR, y es a través de otras como la proteína MLN64 (Metastatic Lymph Node 64) que realiza sus funciones biológicas. MLN64, comparte el dominio START (por sus siglas en inglés StAR-related Transfer domain) con la StAR y por lo tanto podría estar involucrada en el transporte de colesterol hacia la mitocondria[25]. En la regulación crónica ocurre el incremento en la transcripción y traducción de genes que codifican para las enzimas esteroidogénicas e incluso la fosforilación de factores de la transcripción. Este tipo de regulación favorece el incremento en la síntesis de progesterona sin que se presenten variaciones sustanciales en sus concentraciones circulantes a lo largo del embarazo. De manera similar a lo que ocurre en otros tejidos, la biotransformación del colesterol a pregnenolona es catalizada por el complejo enzimático citocromo P450scc. Las células del sinciciotrofoblasto, a diferencia de las células suprarrenocorticales y las gónadas, expresan a la enzima 1,3P-HSD responsable de la síntesis mitocondrial de la progesterona. La placenta humana carece de la17 hidroxilasa y 17,20 desmolasa, por lo que no puede sintetizar derivados del androstano y del estrano a partir de precursores como la progesterona[26].
Se desconocen todavía los mecanismos que modulan la biosíntesis de hormonas esteroides en la placenta humana; sin embargo, existe suficiente evidencia observada en cultivos primarios de células del trofoblasto del papel del AMPc en la producción crónica de progesterona. A este respecto, los análogos de AMPc estimulan a largo plazo la expresión de genes que codifican para algunos componentes de la maquinaria esteroidogénica[27] como P450scc y adrenodoxina. Por otra parte, en cultivos de células inmortalizadas de coriocarcinoma (BeWo), la presencia de AMPc induce la acumulación del ARNm de enzimas esteroidogénicas como la P450scc[28].
Aún cuando no se conocen las principales vías de señalización para la síntesis de progesterona en la placenta, uno de los mecanismos de transducción de señales mejor estudiado es el mediado por PKA. En mitocondrias aisladas de la placenta humana se han identificado proteínas fosforiladas en serina y treonina y en menor grado en tirosina[15]. Esto sugiere la actividad potencial de cinasas. La importancia de la vía AMPc-PKA ha sido demostrada en células BeWo y en mitocondrias aisladas de la placenta humana, donde la utilización de H89 (inhibidor de la actividad de PKA) produce disminución de la síntesis de progesterona, sugiriendo de manera importante la participación de esta cinasa en la regulación de la esteroidogénesis placentaria[15,29]. Existe evidencia de que en cultivos primarios de células derivadas del trofoblasto o en células inmortalizadas del coriocarcinoma, la estimulación con hormonas como hCG, eleva el contenido intracelular de AMPc que induce la activación de PKA, lo que a su vez se traduce en el incremento de la síntesis de progesterona.
En estudios realizados por nuestro grupo utilizando mitocondrias aisladas de la placenta humana, se demostró la interacción directa entre PKA, AKAP-121 y PTPD[13,15]. Por otra parte, también se observó que existe actividad de PKA que no se modifica por AMPc, sin embargo, en presencia de H89 disminuye la síntesis de progesterona pero en forma paradójica incrementa la incorporación de 32P a 3 proteínas mitocondriales. En conjunto, todos estos hallazgos sugieren la presencia de un proceso de activación constitutiva de PKA[15] que en el contexto de la esteroidogénesis placentaria resulta imprescindible para el mantenimiento de las concentraciones plasmáticas de la progesterona necesarias a lo largo de la gestación1301. Lo anterior sugiere que la vía mediada por PKA juega un papel importante en la regulación de la esteroidogénesis placentaria, aunque no se descarta la posibilidad de la participación de otras cascadas de transducción de señales ya sea incrementando o modulando la biosíntesis hormonal.
En la placenta humana, resulta de gran relevancia elucidar las vías de señalización asociadas y la posibilidad de que las cinasas participantes formen complejos con proteínas de anclaje cerca de su sitio de acción, lo cual permitiría la integración de señales mediadas por AMPc con efectores de otras cascadas para regular procesos específicos inducidos por hormonas. Así mismo, tampoco se descarta la presencia de un sistema completo de señalización dentro de la mitocondria que module la síntesis de la progesterona mediante la fosforilación reversible de proteínas, mecanismo que ha sido descrito en mitocondrias de mamífero, donde la fosforilación de complejos de la cadena respiratoria permite una regulación a corto plazo de la fosforilación oxidativa[31].
Células de Leydig[11]
]]> La testosterona y la 5α-dihidrotestosterona son dos hormonas importantes para el desarrollo del fenotipo masculino, de los caracteres sexuales secundarios y la espermatogénesis,incluyendo la conducta sexual. En el testículo, las células de Leydig son las responsables de la síntesis de andrógenos. La síntesis de testosterona ocurre a través de dos vías conocidas como Δ4 y Δ5 [1,24,32] (Figura 6). En el humano, a diferencia del roedor, la vía predominante para la síntesis de testosterona es la vía de los Δ5. La síntesis de testosterona en las células de Leydig está regulada en forma aguda principalmente por el efecto de la LH, la cual, al unirse y activar a receptores acoplados a proteínas G estimula la síntesis de AMPc. La acumulación intracelular de AMPc conduce a la activación de PKA y a la fosforilación de la proteína StAR. Esta cascada de transducción de señal no es la única en activarse, ya que el testículo genera una gran variedad de moléculas reguladoras que inducen vías activadoras independientes de AMPc, como es el caso de los factores de crecimiento como EGF e IGF-I, los que estimulan la expresión de StAR y la síntesis de hormonas esteroides a través de vías que involucran a MAPK y PKC[21,33,34]. Algunas citocinas, derivadas de los macrófagos y del ácido araquidónico producen la activación de PKC y en ciertos casos éstos funcionan en sinergia con LH para promover la esteroidogénesis testicular[21,35,36]. Estas observaciones sugieren la presencia en el testículo de tres vías de señalización que al activarse y comunicarse entre ellas, modifican la tasa de síntesis hormonal. La participación de otras vías de señalización en la esteroidogénesis testicular no sólo involucra a las diferentes cinasas, sino también al tipo de segundo mensajero que induce su activación. Se conoce el efecto predominante de AMPc/PKA en la esteroidogénesis; sin embargo, en células de Leydig obtenidas de ratas adultas, también se han identificado la presencia de transcritos de la guanilato ciclasa (encargada de la producción del segundo mensajero GMPc), de la cinasa PKG I y II y de la fosfodiesterasa 5 (PDE5-específica para la degradación de GMPc). En estos estudios, se observó que la estimulación de la guanilato ciclasa asociada a la inhibición de la PDE5, produjo un incremento de la esteroidogénesis y de la fosforilación de StAR[37]. Aún no se conoce la modulación precisa de esta vía y la forma en la que podría estar interactuando con las cascadas dependientes de AMPc. Como ocurre con otras células esteroidogénicas, las células de Leydig cuentan con sistemas de fosforilación/ desfosforilación dependientes de la acción coordinada de cinasas y fosfatasas. En las células MA-10, la LH activa la cascada de PKA y provoca tanto la fosforilación de proteínas directamente relacionadas con la esteroidogénesis, como la desfosforilación mediada por fosfatasas de tirosinas, siendo ambos procesos necesarios para la biosíntesis hormonal[5,38,39].
Corteza Suprarrenal
Las hormonas que se sintetizan en la corteza suprarrenal ejercen importantes funciones como la regulación del equilibrio hidroelectrolítico, la respuesta inmunitaria al estrés, el metabolismo de carbohidratos y el mantenimiento de los caracteres sexuales secundarios, entre otros. La corteza suprarrenal está dividida estructural y funcionalmente en 3 capas, la glomerular, la fasciculada y la reticular donde se producen los mineralocorticoides, glucocorticoides y los esteroides sexuales, respectivamente. El principal estímulo para la síntesis de glucocorticoides y esteroides sexuales es la hormona adrenocorticotrófica (ACTH), cuyo efecto es a través de la generación de AMPc y activación de la PKA. Por otro lado, la angiotensina II y el potasio constituyen los principales estímulos para la producción de mineralocorticoides a través de la vía IP3/ DAG y la activación PKC[24].
La primera reacción comprometida en la vía biosintética de esteroides es la biotransformación del colesterol a pregnenolona por P450scc[40]. En el retículo endoplásmico de la corteza suprarrenal, la enzima 3P-HSD transforma la pregnenolona a progesterona. Posteriormente, la enzima P450c21 (CYP21A2), la cual se expresa en las 3 capas de la corteza suprarrenal, cataliza la conversión de la progesterona y de la17a-hidroxiprogesterona en precursores de las hormonas biológicamente activas, aldosterona y cortisol (Figura 7). Los andrógenos se forman por la acción de la CYP17, una enzima única que también se encuentra en los ovarios y testículos y que cuenta con actividades de 17a-hidroxilasa y 17,20 desmolasa. La actividad de este complejo enzimático convierte la 17a-pregnenolona y 17a-progesterona en dehidroepiandrosterona y androstendiona, respectivamente[19,24] (Figura 7). La producción de mineralocorticoides, glucocorticoides y de esteroides sexuales por la corteza suprarrenal se regula a través de varios estímulos externos que actúan en forma específica mediante la activación de distintas cascadas de señalización. La ACTH incrementa principalmente la producción de glucocorticoides y esteroides sexuales mediante la vía AMPc-PKA, a través de la fosforilación de StAR y de SF-1. Las hormonas con actividad mineralocorticoide dependen de factores como la angiotensina II y el potasio a través de la vía del IP3/DAG-PKC[24].
La respuesta aguda a la acción de la ACTH implica la unión al receptor 2 de melanocortina, la activación de proteínas Gs, la generación de AMPc y activación de PKA. Esta vía induce la fosforilación de StAR y el acceso de colesterol a la mitocondria. La respuesta crónica favorece la fosforilación de factores de transcripción de enzimas esteroidogénicas como el CREM (modulador de los elementos de respuesta a AMPc)[24]. En ambas respuestas, se requieren de eventos de fosforilación y acoplamiento de la desfosforilación catalizados por fosfatasas dependientes de AMPc para la expresión, por ejemplo, de CYP17[41]. En modelos in vivo e in vitro, la estimulación con ACTH resultó en la activación de fosfatasas de tirosinas. Estos resultados sugieren la dependencia de AMPc de dichas fosfatasas, cuyo efecto es mediado por PKA. En la esteroidogénesis suprarrenal pueden coexistir varias vías de señalización como se observa en ciertos tipos celulares suprarrenocorticales, en los cuales a pesar de ser deficientes de PKA, la ACTH, estimula la fosforilación de las MAPK, cuya activación temporal parece ser necesaria para la esteroidogénesis; efecto similar al observado en las células MA-10 de Leydig[42]. Estos datos sugieren que en algunos tejidos con capacidad esteroidogénica, existe comunicación entre efectores de dos cascadas de transducción diferentes[43].
Existen otros factores que actúan en la corteza suprarrenal regulando la síntesis de hormonas esteroides como son el EGF y el factor de crecimiento de fibroblastos básico (FGFb), que modulan la expresión de CYP17 mediante la cascada de MAPK. Los ésteres de forbol también modulan la expresión de CYP17 activando la vía de PKC[19].
En las células glomerulares la Ang II, a través de las MAPK, aumenta la expresión de StAR y de la 3 P-HSD, incrementando así la síntesis de aldosterona[44]. Aún hace falta establecer tanto las vías de señalización que se inducen en respuesta a los diferentes estímulos que actúan sobre las tres capas suprarrenocorticales, así como la potencial participación de otras moléculas que estarían funcionando como anclaje de los efectores de las cascadas de transducción de señales.
]]> Cuerpo Lúteo
El ovario se divide en tres regiones: la corteza externa conformada por el epitelio germinal y los folículos, la médula de localización central constituida por tejido estromal y la región del hilio[45]. Las células esteroidogénicas del ovario son las células de la granulosa, las cuales forman parte del compartimento vascular que rodea a los ovocitos y las células de la teca que están ubicadas en el estroma ovárico. Ambos compartimentos están separados por la lámina basal[46]. Las funciones principales del ovario son la ovogénesis y la síntesis de hormonas esteroides. La unidad funcional del ovario es el folículo, el cual está constituido por células de la granulosa, de la teca e intersticiales que rodean al ovocito en crecimiento, funcionando como soporte nutricional además de llevar a cabo la síntesis de hormonas esteroides[47]. El ovario secreta esteroides como la pregnenolona, progesterona, 17a-hidroxiprogesterona, 17a-hidroxipregnenolona, dehidroepiandrosterona, androstendiona, testosterona, estrona y 17P-estradiol[48-49]. Bajo el estímulo de hormonas, factores de crecimiento y citocinas, la biosíntesis de esteroides cambia cualitativa y cuantitativamente a lo largo del ciclo ovárico, presentando una intensa actividad esteroidogénica durante la fase lútea y pasando de ser un órgano principalmente productor de estrógenos a uno predominantemente secretor de progesterona. En el humano, la esteroidogénesis ovárica ocurre de acuerdo con la teoría de dos células/dos gonadotropinas[50].
La producción de estrógenos por las células de la teca, requiere de la síntesis de precursores androgénicos (androstendiona) a partir del colesterol, bajo la acción de la LH. Los andrógenos difunden hacia el compartimento avascular de las células de la granulosa. A este nivel, y por efecto de la FSH, los andrógenos son biotransformados a estrógenos por acción de la aromatasa (CYP19)[51]. En la fase lútea, el compartimento ocupado por las células de la granulosa se vasculariza, permitiendo así el aporte de colesterol, aunado a un incremento en la expresión de enzimas esteroidogénicas que resulta en el aumento de la producción de progesterona y estrógenos derivados de los andrógenos de la teca (Figura 8). En el ovario, las células de la granulosa, por acción de la FSH, constituyen la principal fuente de estrógenos requeridos para el crecimiento, la ovulación y el desarrollo de las características sexuales secundarias, entre otras. Durante la fase lútea o progestacional del ciclo ovárico, las células de la granulosa luteinizadas se encargan de la producción de progesterona, hormona responsable de los cambios durante la fase lútea del ciclo menstrual y esencial para el mantenimiento del embarazo[1]. Una vez ocurrida la ovulación, las células del folículo ovárico van a constituir el cuerpo lúteo, el cual sintetiza predominantemente progesterona en respuesta a la LH, a través de la vía AMPc/PKA. Como en otras células esteroidogénicas, existen otras vías de señalización que se activan en forma dependiente de AMPc pero independiente de PKA, como en las células de la granulosa, donde a través de factores intercambiadores de nucleótidos de guanina (dependientes de AMPc) se activan también las vías de MAPK, PI3-K y PKB. La participación MAPK ha sido implicada en la regulación de la esteroidogénesis utilizando LH y hCG, las que además de elevar las concentraciones de AMPc, activan a ERK1/2 (cinasas efectoras de MAPK) en células de la granulosa luteinizadas. También se ha observado que durante la luteinización de las células de la granulosa, la secreción de progesterona inducida por hCG coincide con un descenso en la expresión de la subunidad catalítica de PKA, lo que sugiere que durante la luteinización de las células de la granulosa la síntesis de progesterona es menos dependiente de PKA[52].
En el cuerpo lúteo de la oveja se han descrito dos poblaciones celulares, unas de aproximadamente 12-20 (om de diámetro que responden a LH con incrementos de 5 a 15 veces las concentraciones circulantes de progesterona y otras de mayor tamaño que de manera basal secretan suficientes cantidades de progesterona. Este último tipo celular se caracteriza por no responder a la LH a pesar de expresar los receptores para esta hormona[53].
No se conocen los mecanismos responsables de la modulación en las células lúteas de lo que parece ser un tipo de regulación aguda y otra probablemente constitutiva; la que asegura la adecuada producción de progesterona, durante el primer trimestre de gestación con el objeto de establecer y mantener el embarazo.
Conclusiones
La esteroidogénesis es importante para el mantenimiento de la homeostasis hidroelectrolítica, así como de las funciones reproductivas y metabólicas del cuerpo. De particular importancia es el caso de la placenta, en donde la síntesis de progesterona es indispensable para el mantenimiento del embarazo y la perpetuación de la especie. De ahí surge el interés de conocer los mecanismos que regulan o modulan la biosíntesis hormonal. Se sabe que existen tejidos esteroidogénicos que tienen la capacidad de responder en forma aguda o crónica dependiendo de las necesidades del organismo, las que son modificadas durante las distintas etapas del crecimiento y desarrollo, incluyendo las condiciones de estrés o de inflamación o de un proceso fisiológico como el embarazo; sin embargo, se desconocen los factores endocrinos, paracrinos o incluso autocrinos que dan origen a la secuencia de señales a nivel intracelular y que determinan que un tejido secrete una hormona específica en el orden de minutos o bien en el transcurso de horas a días. Para esto, las células utilizan vías de señalización donde los principales efectores son cinasas que fosforilan proteínas mediadoras que a su vez activan o inhiben la expresión y/o actividad de enzimas, incluyendo factores de transcripción o proteínas que son responsables de la respuesta esteroidogénica a corto o largo plazo. Para realizar estas funciones, las células utilizan mecanismos de transducción de señales que son comunes a varios tipos celulares, la diferencia radica en el estímulo que inicia la cascada de señalización y de comunicación entre los componentes de cada una de las vías que al final se traducen en ciclos de fosforilación y que son claves, en este caso, para la esteroidogénesis, así como para los procesos generales de crecimiento y diferenciación celular. En esta revisión se mencionaron algunas de las principales vías de señalización que utilizan los tejidos esteroidogénicos sujetos a la regulación aguda o crónica. Cabe mencionar que de las vías más estudiadas se encuentra la mediada por PKA; sin embargo, se conocen otras cinasas que actuando de manera independiente también participan en estos procesos. En este sentido, hay evidencia de que las células utilizan proteínas de anclaje que fijan a componentes específicos de las vías en localizaciones subcelulares específicas, formando así complejos de señalización que hacen más eficiente la actividad de las cinasas e incluso de las fosfatasas. La participación de estas proteínas de anclaje sugiere que los eventos de fosforilación y desfosforilación de proteínas están altamente regulados no sólo por las diferentes vías activadas en respuesta a hormonas tróficas o a otros factores, sino por la localización específica dentro de la célula y en la proximidad de un organelo en particular. El conocimiento de la secuencia que ocurre en la activación de los efectores de una cascada, en particular en los tejidos esteroidogénicos de respuesta aguda o crónica, permitirá identificar puntos clave de regulación que podrían ser objetivos terapéuticos a través del diseño de fármacos que modulen positiva o negativamente la producción de algún producto hormonal en los casos donde la deficiencia pudiera manifestarse como un espectro de alteraciones en la reproducción, el metabolismo y el balance hídrico o bien en casos de neoplasias hormono dependientes o para el tratamiento de mujeres gestantes con riesgo de aborto o de parto pre-término asociado a una disminución en los niveles circulantes de la progesterona.
Agradecimientos
]]> Los autores agradecen el apoyo parcial de los proyectos DGAPA IN217609 y IN211912, así como del proyecto de CONACyT 168025. Erika Gómez Chang es becaria del CONACyT (215670).
Referencias
1. Hu, J., Zhang, Z., Shen, W.J. & Azhar, S. Cellular cholesterol delivery, intracellular processing and utilization for biosynthesis of steroid hormones. Nutr. Metab. (Lond.) 7, 47 (2010). [ Links ]
2. Granot, Z. et al. Proteolysis of normal and mutated steroidogenic acute regulatory proteins in the mitochondria: the fate of unwanted proteins. Mol. Endocrinol. 17, 2461-2476 (2003). [ Links ]
3. Manna, P.R., Dyson, M.T. & Stocco, D.M. Regulation of the steroidogenic acute regulatory protein gene expression: present and future perspectives. Mol. Hum. Reprod. 15, 321-333 (2009). [ Links ]
4. Corso, M. & Thomson, M. Protein phosphorylation in Mitochondria from human placenta. Placenta 22, 432-439 (2001). [ Links ]
5. Thomson, M. Evidence of undiscovered cell regulatory mechanisms: phosphoproteins and protein kinases in mitochondria. Cell Mol. Life Sci. 59, 213-219 (2002). [ Links ]
6. Gorostizaga, A. et al. Tyrosine phosphatases in steroidogenic cells: Regulation and function. Mol. Cell Endocrinol. 265-66, 131-137 (2007). [ Links ]
7. Dremier, S., Kopperud, R., Doskeland, S.O., Dumon, J.E. & Maenhaut, C. Search for new cyclic AMP binding proteins. FEBS Lett. 546, 103-107 (2003). [ Links ]
8. Dodge-Kafka, K.L. &Kapiloff, M.S. The mAKAP signaling complex: Integration of AMPc, calcium, and MAP kinase signaling pathways. Eur. J. Cell Biol. 85, 593-602 (2006). [ Links ]
9. Aye, T.T. et al. Selectivity in enrichment of AMPc-dependent protein kinase regulatory subunits type I and type II and their interactors using modified AMPc affinity resins. Mol. Cell Proteomics 8, 1016-1028 (2009). [ Links ]
10. Taylor, S.S., Knighton, D.R., Zheng, J., Ten Eyck, L.F. & Sowadski, J.M. cAMP-dependent protein kinase and the protein kinase family. Faraday Discuss. 93, 143-152 (1992). [ Links ]
11. McKnight, G.S. et al. Cyclic AMP, PKA, and the physiological regulation of adiposity. Recent Prog. Horm. Res. 53, 139-159 (1998). [ Links ]
12. Taylor, S.S. et al. Signaling through AMPc and AMPc-dependent protein kinase diverse strategies for drug design. Biochim. Biophys. Acta 1748, 16-26 (2008). [ Links ]
13. Feliciello, A., Gottesman, M.E. & Avvedimento, E.V. cAMP-PKA signaling to the mitochondria: protein scaffolds, mRNA and phosphatases. Cell Signal 17, 279-287 (2005). [ Links ]
14. Livigni, A. et al. Mitochondrial AKAP121 links AMPc and src signaling to oxidative metabolism.Mol. Biol. Cell. 17, 263-271 (2006). [ Links ]
15. Gómez-Concha, C., Flores-Herrera, O., Olvera-Sánchez, S., Espinosa-García, M.T. & Martínez, F. Progesterone synthesis by human placental mitocondria is sensitive to PKA inhibition by H89. Int. J. Biochem. Cell Biol. 43, 1402-1411 (2011). [ Links ]
16. Pidoux, G. & Taskén, K. Specificity and spatial dynamics ofprotein kinase A signaling organized by A-kinase-anchoring proteins. J. Mol. Endocrinol. 44, 271-284 (2010). [ Links ]
17. Fleury, A., Mathieu, A.P., Ducharme, L., Hales, D.B. & LeHoux, J.G. Phosphorylation and function of the hamster adrenal steroidogenic acute regulatory protein (StAR). J. Steroid. Biochem. Mol. Biol. 91, 259-271 (2004). [ Links ]
18. Pollheimer, J. & Knõfler, M. Signalling pathways regulating the invasive differentiation of human trophoblasts: a review. Placenta 26, S21-30 (2005). [ Links ]
19. Sewer, M.B., Li, D., Dammer, E.B., Jagarlapudi, S. & Lucki, N. Multiple Signaling Pathways Coordinate CYP17 Gene Expression in the Human Adrenal Cortex. Acta Chim. Slov. 55, 53-57 (2008). [ Links ]
20. Horbinski, C. & Chu, C.T. Kinase signaling cascades in the mitochondrion: a matter oflife or death. Free Radic. Biol. Med. 38, 2-11 (2005). [ Links ]
21. Stocco, D.M., Wang, X., Jo, Y. & Manna, P.R. Multiple signaling pathways regulating steroidogenesis and steroidogenic acute regulatory protein expression: more complicated than we thought. Mol. Endocrinol. 19, 2647-2659 (2005). [ Links ]
22. Maloberti, P. et al. Silencing the expression of mitocondrial acyl-CoA thioesterase I and acyl-CoA synthetase 4 inhibits hormone-induced steroidogenesis. FEBS J. 272, 1804-1814 (2005). [ Links ]
23. Wang, X., Walsh, L.P., Reinhart, A.J. & Stocco, D.M. The role of arachidonic acid in steroidogenesis and steroidogenic acute regulatory (StAR) gene and protein expression. J. Biol. Chem. 275, 20204-20209 (2000). [ Links ]
24. Sanderson, T. The Steroid Biosynthesis pathway as a target for endocrine-disrupting chemicals. Toxicol. Sci. 94, 3-21 (2006). [ Links ]
25. Moog-Lutz, C. et al. MLN64 exhibits homology with the steroidogenic acute regulatory protein (STAR) and is over-expressed in human breast carcinomas. Int. J. Cancer 71, 183-191 (1997). [ Links ]
26. Tuckey, R.C. Progesterone synthesis by the human placenta. Placenta 26, 273-281 (2005). [ Links ]
27. Nulsen, J.C. et al. Control of the steroidogenic machinery of the human trophoblast by cyclic AMP. J. Reprod. Fertil. Suppl. 37, 147-153 (1989). [ Links ]
28. Ringler, G.E. & Strauss, J.F. 3rd. In vitro systems for the study of human placental endocrine function. Endocr. Rev. 11, 105-123 (1990). [ Links ]
29. Maldonado-Mercado, M.G., Espinosa-García, M.T., Gómez-Concha, C., Monreal-Flores, J. & Martínez, F. Steroidogenesis in BeWo cells: role of protein kinase A and benzodiazepines. Int. J. Biochem. Cell Biol. 40, 901-908 (2008). [ Links ]
30. Duan, L., Yan, D., Zeng, W., Yang, X. & Wei, Q. Effect of progesterone treatment due to threatened abortion in early pregnancy for obstetric and perinatal outcomes. Early Hum. Dev. 86, 41-43 (2010). [ Links ]
31. Acín-Pérez, R. et al. Modulation of mitochondrial protein phosphorylation by soluble adenylyl cyclase amelioreates cytochrome oxidase defects. EMBO Mol. Med. 1, 392-406 (2009). [ Links ]
32. Wilson, J.D. Dihydrotestosterone formation in cultured human fibroblasts. J. Biol. Chem. 250, 3498-3504 (1975). [ Links ]
33. Lin, T., Wang, D., Hu, J. & Stocco, D.M. Upregulation of human chorionic gonadotrophin-induced steroidogenic acute regulatory protein by insulin-like growth factor in rat Leydig cells. Endocrine 8, 73-78 (1998). [ Links ]
34. Manna, P.R., Huhtaniemi, I.T., Wang, X.J., Eubank, D.W. & Stocco, D.M. Mechanisms of epidermal growth factor signaling: regulation of steroid biosynthesis and the steroidogenic acute regulatory protein in mouse Leydig tumor cells. Biol. Reprod. 67, 1393-1404 (2002). [ Links ]
35. Wang, X.J. et al. Interaction between arachidonic acid and AMPc signaling pathways enhances steroidogenesis and StAR gene expression in MA-10 Leydig tumor cells.Mol. Cell Endocrinol. 188, 55-63 (2002). [ Links ]
36. Bornstein, S.R., Rutkowski, H. & Vrezas, I. Cytokines and steroidogenesis. Mol. Cell Endocrinol. 215, 135-141 (2004). [ Links ]
37. Andric, S.A., Janjic, M.M., Stojkov, N.J. & Kostic, T.S. Protein kinase G-mediated stimulation of basal Leydig cell steroidogenesis. Am. J. Physiol. Endocrinol. Metab. 293, 1399-1408 (2007). [ Links ]
38. Cornejo Maciel, F., Poderoso, C., Gorostizaga, A., Paz, C. & Podestá, E.J. LH/chorionic gonadotropin signaling pathway involves protein tyrosine phosphatase activity downstream of protein kinase A activation: evidence of an obligatory step in steroid production by Leydig cells. J.Endocrinol. 170, 403-411 (2001). [ Links ]
39. Paz, C. et al. Protein tyrosine phosphatases are involved in LH/ chorionic gonadotropin and 8Br-AMPc regulation of steroidogenesis and StAR protein levels in MA-10 Leydig cells. J. Endocrinol. 175, 793-801 (2002). [ Links ]
40. García-Pérez, C. & Martínez, F. Los mecanismos que controlan la síntesis de progesterona en la placenta humana. REB 21, 181-189 (2002). [ Links ]
41. Sewer, M.B. & Waterman, M.R. AMPc-dependent transcription of steroidogenic genes in the human adrenal cortex requires a dual-specificity phosphatase in addition to protein kinase A. J. Mol. Endocrinol. 29, 163-174 (2002). [ Links ]
42. Hirakawa, T. & Ascoli, M. The lutropin/choriogonadotropin receptor-induced phosphorylation of the extracellular signal-regulated kinases in Leydig cells is mediated by a protein kinase a-dependent activation of ras. Mol. Endocrinol. 17, 2189-2200 (2003). [ Links ]
43. Poderoso, C. et al. A mitochondrial kinase complex is essential to mediate an ERK1/2-dependent phosphorylation of a key regulatory protein in steroid biosynthesis. PLoS One 3, e1443 (2008). [ Links ]
44. Otis, M. & Gallo-Payet, N. Role of MAPKS in angiotensin II induced steroidogenesis in rat glomerulosa cells. Mol. Cell Endocrinol. 265-266, 126-130 (2007). [ Links ]
45. Carr, B.R. in William's Textbook of Endocrinology (Saunders, W.B.) 751-817 (Hartcourt & Brace, Philadelphia, PA, 1998). [ Links ]
46. Weakly, B.S. Electron microscopy ofthe oocyte and granulosa cells in the developing ovarian follicles ofthe golden hamster. J. Anat. 100, 503-534 (1966). [ Links ]
47. Gore-Langton, R.E. & Armstrong, D.T. en The Physiology of Reproduction. (Knobil, E & Neill, J.D.) 571-627 (Raven Press, New York, 1994). [ Links ]
48. Baird, D.T. & Fraser, I.J. Concentration of estrone and estradiol-17p in follicular fluid and ovarian venous blood ofwomen. Clin. Endocrinol. 4, 171-175 (1969). [ Links ]
49. Baird, D.T., Burger, P., Heavon-Jones, G.D. & Scaramuzzi, R.J. The site of secretion of androstenedione in non-pregnant women. J. Endocrinol. 63, 201-212 (1974). [ Links ]
50. Ryan, K.J. & Petro, Z. Steroid biosinthesis by human ovarian granulosa and theca cells. J. Clin. Endocrinol. Metab. 26, 56-52 (1966). [ Links ]
51. Havelock, J.C., Rainey, W.E. & Carr, B.R. Ovarian granulosa cell lines. Mol. Cell. Endocrinol. 228, 67-78 (2004). [ Links ]
52. Chin, E.C. & Abayasekara, D.R. Progesterone secretion by luteinizing human granulose cells: a possible AMPc-dependent but PKA-independent mechanism involved in its regulation. J.Endocrinol. 183, 51-60 (2004). [ Links ]
53. Niswender, G.D. Molecular control of luteal secretion of progesterone. Reproduction 123, 333-339 (2002). [ Links ]
]]> Información sobre los autores
Erika Gómez-Chang
Erika Gómez-Chang es Médica Cirujana egresada de la Facultad de Medicina de la Universidad Nacional Autónoma de México. Actualmente es Profesora de Asignatura "A" dentro del Programa de Alta Exigencia Académica (PAEA) y candidata al grado de Doctor en Ciencias en el Programa de Doctorado en Ciencias Biomédicas de la UNAM. Es Socio Estudiante de la Sociedad Mexicana de Bioquímica desde 2011. Ha desempeñado el cargo de jurado en la fase práctica ante paciente real en exámenes profesionales de la carrera de Médico Cirujano y ha sido Profesora del área de Bioquímica en el Curso Universitario de Preparación para el Examen Nacional de Residencias Médicas. Ha participado en Congresos Nacionales e Internacionales y es coautora en un capítulo del libro "Recent Advances in Research on the Human Placenta".
Fernando Larrea
Fernando Larrea es Médico Cirujano, egresado de la Facultad de Medicina (FM) de la Universidad Nacional Autónoma de México (UNAM) con estudios de posgrado en Biología de la Reproducción Humana en el Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán (INNSZ) y en la División Biomédica del Consejo de Población en la Ciudad de Nueva York, NY bajo los auspicios de la Fundación Fogarty de los Estados Unidos de Norte América. Desde 1981 es investigador titular del INNSZ, adscrito al Departamento de Biología de la Reproducción y jefe de este departamento desde 1993. Es miembro del Sistema Nacional de Investigadores Nivel III, de la Academia Nacional de Medicina y es Certificado por el Consejo Mexicano de Endocrinología. Es Profesor Titular del Curso de Subespecialización en Biología de la Reproducción Humana de la División de Estudios de Posgrado de la Facultad de Medicina de la UNAM. Cuenta con 148 publicaciones y ha participado en 320 congresos nacionales e internacionales.
Federico Martínez-Montes
Federico Martínez-Montes es Profesor Titular «C» de Tiempo Completo en la Facultad de Medicina de la UNAM y pertenece al Sistema Nacional de Investigadores nivel II; recibió el Reconocimiento Catedrático de la UNAM y actualmente es PRIDE "D". Obtuvo la Cátedra Extraordinaria "Salvador Zubirán Anchondo" de la Facultad de Medicina. Recibió el Premio GlaxoSmithKline en investigación básica, tercer lugar en el 2006. Pertenece a varias sociedades científicas y ha desempeñado funciones de revisor de proyectos a nivel nacional e internacional. Ocupó puestos directivos en la Sociedad Mexicana de Bioquímica, A.C. de la que fue su presidente en el bienio 2007-2009 y Consejero de Ciencias Básicas de la Academia de Investigación en Biología de la Reproducción, A.C. en varias ocasiones. Ha participado como revisor de la revista Placenta. Fue becario de la Ford Foundation y de la Rockefeller Foundation para estudios de colaboración en el área de bioquímica reproductiva. Su área de investigación está relacionada con el mecanismo de control de la síntesis de la hormona progesterona en la placenta humana. Ha publicado más de 50 artículos tanto a nivel internacional como a nivel nacional y cuenta con varios capítulos en libros. Ha desarrollado varios programas interactivos de apoyo a la docencia y es editor del libro de Bioquímica de Laguna. Ha graduado alumnos tanto a nivel licenciatura como a nivel posgrado.
Ha sido Consejero Técnico de laFacultad de Medicina; Miembro del Comité Académico del Programa de Doctorado en Ciencias Biomédicas como representante de los tutores y Miembro del Comité Académico de la carrera de Investigación Biomédica Básica de la Facultad de Medicina.
Actualmente es Miembro del Comité Académico de la Unidad de Biomedicina de la FES-Iztacala, de la Comisión de Investigación de la Facultad de Medicina, de la Comisión PRIDE de la Facultad de Veterinaria y Zootecnia; del Comité de Evaluación de Proyectos PAPIIT y del Grupo de Evaluación del Posgrado en el Foro Consultivo Científico y Tecnológico. Actualmente es Coordinador del Doctorado en Ciencias Biomédicas, UNAM.
]]>