
Equivalencia ambiental en la productividad de la vegetación
Environmental equivalence in vegetation productivity
Fernando Paz–Pellat*, Magali Odi–Lara, Alejandro Cano–González, Martín A. Bolaños–González, Alfonso Zarco–Hidalgo
Campus Córdoba y Montecillo, Colegio de Postgraduados. Km. 36.5 Carretera México–Texcoco, 56230.Montecillo, Estado de México. *Autor responsable: (pellat@colpos.mx)
]]> Recibido: Diciembre, 2008.
RESUMEN
La estimación de la productividad de la vegetación por cambios ambientales o de manejo requiere de una modelación caso por caso, por lo que resulta en requerimientos altos de información. Una alternativa es usar un enfoque global que defina la relación, en forma compacta, entre la productividad y los factores ambientales o de manejo. En este trabajo se presenta el concepto de la equivalencia ambiental en la productividad de la vegetación, cultivos, usando para esto el modelo expo–lineal del crecimiento y la propiedad de un conjunto de líneas rectas que se intersectan en un punto común. Se presenta un marco teórico genérico para hacer equivalentes las productividades asociadas a diferentes niveles de estrés o manejo y a diferentes medios ambientales. Toda la variabilidad durante la etapa vegetativa puede ser compactada como un punto en un espacio supra–paramétrico. La evidencia experimental presentada soporta la propuesta teórica desarrollada.
Palabras clave: Equivalencia, espacio paramétrico y supra–paramétrico, modelo expo–lineal, productividad vegetación.
ABSTRACT
Estimation of vegetation productivity due to environmental or management changes requires case by case modeling, resulting in high requirements of information. One alternative is to use a global approach that defines, in a compact form, the relationship between productivity and environmental or management factors. This paper presents the concept of environmental equivalence in vegetation productivity, or crops, using an expo–linear growth model and the property of a set of straight lines that intersect at a common point. A theoretical framework is presented to generate productivity equivalents associated with different levels of stress or management and in different environments. All of the variability occurring during the vegetative stage can be compacted as a supra–parametric point in space. The experimental evidence presented supports the theoretical proposal developed.
Key words: Equivalence, parametric and supra–parametric space, expo–linear model, vegetation productivity.
]]>INTRODUCCIÓN
La productividad (biomasa y área foliar, principalmente) de la vegetación es dependiente del medio ambiente (temperatura y radiación solar, principalmente) en que se desarrolla, además de factores de estrés y manejo (agua, nutrientes, plagas y enfermedades, competencia, etc.). La distinción entre ambiente y factores de estrés y manejo es algo arbitraria, ya que una variable asociada al medio ambiente como la temperatura puede ser controlada por manejo en un invernadero; no obstante lo anterior, la terminología utilizada es congruente con la de los modelos biofísicos (Jones y Kniry, 1986). En lo general, el término de ambiente es usado en este trabajo para denotar una fecha de siembra, la cual tiene asociada una evolución temporal de variables agrometeorológicas que inciden en los patrones de crecimiento de la vegetación. La modelación de los factores incidentes en la productividad puede hacerse usando modelos mecanicistas semi–empíricos (Jones y Kiniry, 1986; Spitters et al., 1989; Stockle et al., 2003) o usando funciones matemáticas con bases empíricas (Hunt, 1982). En el primer caso, el número de variables es muy alto para predecir una o dos variables de salida de interés, además de la necesidad de conocer su variabilidad temporal. En el segundo caso, las funciones matemáticas no tienen un significado biofísico, por lo que resultan muy restringidas en las aplicaciones.
En la perspectiva de las aplicaciones de los sensores remotos, donde se observan los resultados del proceso biológico del crecimiento de la vegetación, es necesaria una formulación general que permita caracterizar en forma adecuada la productividad. En este trabajo se plantea un esquema integral para hacer equivalentes las condiciones ambientales, incluyendo los factores de manejo y estrés, de tal forma que se puedan interpretar las funciones del crecimiento de la vegetación en un marco unificado. Aunque los resultados presentados son para tres cultivos, los argumentos son válidos para otros tipos de vegetación, incluida la arbórea; esto es discutido en otros trabajos de los autores.
MODELO DEL CRECIMIENTO DE LA VEGETACIÓN
En el espacio del tiempo, la modelación del crecimiento de la vegetación (índice de área foliar o IAF y biomasa o B en peso (a menos que se defina lo contrario, B es usado en términos genéricos para denotar biomasa seca o fresca) por unidad de área (representadas en forma general por la variable V) en la etapa vegetativa, se compone de dos fases: una exponencial y una lineal. La solución clásica de estas dinámicas (consultar cualquier texto de ecuaciones diferenciales) está dada por:
Crecimiento exponencial:
Crecimiento lineal:
]]>
donde, R es una tasa relativa de crecimiento, C es la tasa máxima de crecimiento y tb es el tiempo cuando V(t) = 0 en la fase lineal.
Aunque es posible analizar las dos fases por separado, al hacerlas compatibles en los puntos de transición (Lieth et al., 1996), podemos generar un modelo con una transición gradual entre ambos tipos de crecimiento. Así, en el momento de la transición del crecimiento exponencial al lineal, las tasas de crecimiento deben ser iguales:

El modelo expo–lineal es simplemente la solución de los dos tipos de crecimiento representados por las ecuaciones diferenciales (1) y (2), sujeta a la condición dada por la relación (3) (Goudriaan, 1994):
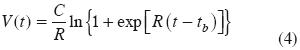
Con las siguientes características:

donde, tT es el tiempo donde la condición (3) se cumple.
En la Figura 1a se muestra el modelo expo–lineal del crecimiento de la vegetación, exhibiendo las características definidas previamente y en la 1b se muestra una transformación invariante (conservación de la métrica asociada) del modelo.
]]> El modelo expo–lineal introducido es usado para mostrar el concepto de equivalencia ambiental y no implica una etiqueta de superioridad en relación a otros modelos. Los argumentos discutidos más adelante pueden ser aplicados a otros modelos matemáticos, después de su transformación lineal.El modelo expo–lineal fue desarrollado para el IAF por Ferrandino (1989) usando la hipótesis de medio homogéneo (medio turbio en términos de transferencia radiativa) y por Goudriaan y Monteith (1990) para la B, usando una relación lineal entre el IAF y la B. La transformación invariante de la Figura 1b es discutida por Monteith (2000).
En la Figura 2 se muestra el crecimiento del IAF para un cultivo de maíz (Huete et al., 1985) donde se presenta una fase reproductiva, en la que las propiedades geométricas u ópticas varían en relación a la fase expo–lineal (etapa vegetativa). Tal como se discute más adelante, la etapa vegetativa (su pendiente) define la productividad de la vegetación.

La transición entre la fase exponencial y la reproductiva (representada por una función exponencial o de otro tipo) puede ser modelada en forma similar que el caso de la transición exponencial a la lineal (Goudriaan, 1994); aunque el modelo es un sigmoide simétrico que puede generalizarse a uno asimétrico (Yin et al., 2003), pero con una pérdida de interpretación biofísica. Los modelos expo–linea–les y sus generalizaciones discutidas utilizan la hipótesis de medios ambientes relativamente similares (principalmente la radiación fotosintética activa), por lo que es posible generalizarlos a medios ambientes variables (Monteith, 2000). No obstante esto, Yuan y Bland (2004) mostraron que los beneficios del modelo ambientalmente variable no mejoran apreciable–mente los resultados de ambiente constante, por lo que la estructura matemática del modelo expo–lineal, ecuación (4) puede ser usada en términos generales. Paz[1] analiza esta situación, desde el punto de vista teórico y experimental, tanto para medios homogéneos, heterogéneos y multi–componentes de la vegetación, concluyendo que el modelo expo–lineal es suficientemente general para ser usado en aplicaciones prácticas.
FUNDAMENTOS DE LAS RELACIONES EN ESPACIOS META–PARAMÉTRICOS
Se puede analizar sólo la fase lineal del modelo expo–lineal discutido anteriormente o una transformación aproximadamente lineal del modelo, tal como la mostrada en la Figura 1b. El uso de la fase lineal o una transformación lineal de la etapa vegetativa es un requisito para los desarrollos mostrados, pero éstos pueden ser presentados en igual forma, aunque más compleja, para curvas de crecimiento que tienen un punto de intersección.
Para definir el contexto de las propiedades de los patrones del crecimiento de la vegetación, fase lineal, podemos analizar un conjunto de líneas rectas que se intersectan en un punto cualquiera (Figura 3a). Bajo esta consideración, las intersecciones ak y las pendientes bk de las líneas rectas (V = ak + bkt) tienen una relación lineal (Figura 3b), definiendo un espacio meta–paramétrico dependiente de las constantes A y B (bk= A + Bak).
]]>
En el caso particular del modelo expo–lineal se puede establecer la hipótesis; la tasa de crecimiento relativa R es más o menos constante (los niveles de estrés y manejo, hasta un umbral crítico, tienen poco efecto), por lo que se espera que las fases lineales tengan un punto aproximado de intersección (Goudriaan y Monteith, 1990; Goudriaan, 1994), como se muestra en la Figura 3.
EQUIVALENCIA AMBIENTAL DE LA PRODUCTIVIDAD DE LA VEGETACIÓN
Para poder hacer equivalentes los medios ambientes (clima) y factores de estrés y manejo, es conveniente analizarlos por separado.
Medio ambiente fijo (misma fecha de inicio del crecimiento)
Considerando la variable de productividad Vy, donde i se refiere al nivel de estrés o manejo para un medio ambiente j (j = 1, por ejemplo, en nuestro caso), en la Figura 3a se muestra la situación donde el nivel de estrés i se manifiesta en un cambio de la pendiente bij en el espacio del tiempo, haciendo un cambio de origen en el punto donde las fases lineales aproximadamente se intersectan (Figura 4a). Se puede suponer que un cambio de pendiente implica un cambio en la productividad de la vegetación (Figura 4b). En la Figura 4b se ha conservado el origen inicial de acuerdo con la Figura 4a, pero las ai,j se hicieron positivas (multiplicadas por – 1) y la relación lineal entre ai,j y bi,j se ha modificado restando a los valores de las bi,j la intersección con ese eje. Así, con la estimación de la pendiente de las fases lineales de la etapa vegetativa del crecimiento podemos estimar la productividad de la vegetación, ya que para una fecha determinada la pendiente es equivalente a un cambio en la productividad V.
Medio ambiente variable (diferentes fechas de inicio del crecimiento)
Usando un cambio de origen similar al mostrado en la Figura 4a, para el caso de medios ambientes variables (j variable), podemos analizar el caso de un mismo nivel de estrés (i = 1, por ejemplo, en nuestro caso), Figura 5a, después de hacer que los inicios del crecimiento de la vegetación sean coincidentes (translación de orígenes a uno en común), Figura 5b.
]]>
En la Figura 5b se muestra que las intersecciones y pendientes de las curvas de crecimiento temporales para el nivel de estrés i y medios ambientes j variables puede ser aproximada por una relación lineal usando argumentos similares a los definidos en la Figura 3. Para cada punto (ai,j, bi,j), podemos trazar una línea recta desde el punto hasta el origen, en forma similar al caso de la Figura 4b y que representa las trayectorias de diferentes niveles de estrés para un ambiente j dado.
Modelo general de equivalencia ambiental
En la situación donde el ambiente y los niveles de estrés o manejo varían simultáneamente, en la Figura 6a se muestra cómo varía la pendiente de las líneas rectas de la fase lineal (o la fase expo–lineal transformada linealmente) del crecimiento de la vegetación. En la Figura 6b (línea punteada fuerte) se muestra el concepto de equivalencias ambientales en la productividad, donde para un valor de ésta, las intersecciones (ambientes y niveles de estrés) en la fase lineal del crecimiento de la vegetación pueden ser múltiples y equivalentes, dependiendo de i y j.

En la Figura 6a se muestra con una línea recta diferentes estreses asociados a un ambiente potencial donde la vegetación puede alcanzar su potencial genético productivo, Figura 6b.
La Figura 6a puede ser compactada en una ecuación que caracterice el espacio meta–paramétrico de los ambientes que definen las líneas bi,j= Ai,j+ Bi,jai,j (ver Figura 3b):
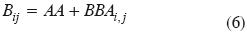
EVIDENCIA EXPERIMENTAL
En este apartado se revisan varios experimentos realizados en las instalaciones del Colegio de Post–graduados en Montecillo, Estado de México, México, bajo las siguientes consideraciones: dado que las mediciones del IAF o B son destructivas, la hipótesis de representatividad de estos datos para un tratamiento y repetición particular fueron revisadas usado diferentes espacios de representación y variables medidas; es decir, se analizaron en forma simultánea mediciones de biomasa, área foliar y altura, revisando las relaciones entre ellas y en el tiempo, para detectar patrones biofísicamente incongruentes. Sólo en aquellos casos donde existía coherencia de patrones se retuvieron los datos para análisis posteriores. Las repeticiones de cada tratamiento fueron consideradas en forma individual, para evitar la hipótesis de homogeneidad física, estacionariedad estadística, asociada a los diseños experimentales. Finalmente, el modelo expo–lineal fue ajustado a través de un proceso de minimización no lineal del error cuadrático de estimación y sólo las curvas con dos o más datos en la fase lineal fueron analizadas. En ningún caso de los mostrados más adelante se realizó un proceso de re–centrado de las curvas de crecimiento.
Experimento con tomate en invernadero
Solorio (2007)[2] realizó un experimento con tomate variedad Río Grande a través de la siembra de 500 semillas en un almacigo de charolas de plástico llenas con sustrato de turba de musgo (peat moss). La germinación ocurrió 3 d de la siembra y permaneció en el almacigo 42 d, tiempo durante el cual se le proporcionaron 14 riegos. A continuación, se realizó el transplante en macetas de plástico con forma de cono truncado, conteniendo un sustrato de tezontle graduado a 2 mm de diámetro, con un orificio para drenar excedencias de agua y un filtro que impide el deslave. Los tratamientos al cultivo consistieron en variaciones de nitrógeno y humedad, en niveles óptimos y restrictivos. De estos factores, se propusieron tres niveles de nitrógeno (100 %, 50 % y 10 % respecto a la dosis óptima requerida por la planta) combinados con dos niveles de humedad (100 % y 27 % del consumo de la humedad aprovechable a capacidad de contenedor, determinada por mediciones directas), de donde se obtuvieron seis tratamientos. Para dosificar el N se procedió conforme al equivalente a la solución de Steiner (Steiner, 1984), usando 280 kg ha–1 de N. Se realizaron tres repeticiones de cada tratamiento, obteniéndose 18 parcelas experimentales, la cuales fueron distribuidas al azar dentro del invernadero. Con respecto al control de plagas del cultivo, se realizaron 10 aplicaciones preventivas principalmente contra mosquita blanca usando PlenumMR (ingredientes activos: picloram 8 % y fluroxypyr 8 %) diluído en agua (3 g L–1) y DAP–PLUSMR (ingredientes activos: ácido fosfórico 32 % y nonil fenol etoxilado 38 %) (1 mL L–1), aplicado con mochila aspersora, a razón de 0.5 y 1 L por parcela experimental, dependiendo del crecimiento de las plantas.
Experimento de cebada en parcelas
Romero (1987)[3] realizó un experimento con cebada (Hordeum vulgare L.) donde utilizó un diseño experimental, considerando como factores la variación del régimen de humedad del suelo en tres etapas de desarrollo de la cebada: vegetativa, reproductiva y maduración. En cada factor se ensayaron cuatro valores de tensión de la humedad en el suelo: 0.5, 3.0, 5.5, y 8.0 atm, equivalentes a 80, 35, 20 y 10 % de humedad aprovechable residual en el suelo. A los tratamientos obtenidos con la matriz experimental se adicionaron otros dos: 0.36 y 15 atm, correspondientes a 85 % y 0 % de humedad aprovechable residual. Los tratamientos se distribuyeron en un diseño experimental de bloques al azar con cuatro repeticiones. Se utilizaron parcelas experimentales de 3 m×3 m las cuales se delimitaron por bordos de 30 cm de altura. Se dejó un espacio de 3.0 m entre parcelas para evitar traslapes de humedad. Las parcelas se protegieron contra la lluvia utilizando estructuras protectoras.
La siembra se realizó de forma manual y en seco (18 de diciembre de 1985), dejando una separación de 18 cm entre hileras. Se utilizó la variedad cerro prieto, con una densidad de siembra de 100 kg ha– 1 y también se sembraron las calles entre parcelas y alrededor del lote experimental.
La fertilización fue manual, con la fórmula 10060–00 kg ha–1 (N, P, K). Se utilizó urea como fuente de N y superfosfato triple de calcio como fuente de P. La mitad del N y todo el P se aplicaron en la siembra y la mitad restante de N aproximadamente al mes de la germinación, haciéndola coincidir con un riego de auxilio. Las malezas se controlaron manualmente. Para controlar plagas se realizaron dos aplicaciones de folidol al 50 %, en las dos primeras semanas de marzo y abril, para controlar pequeñas infestaciones de pulgón de la hoja (Rhopalosiphum padi). Después de la siembra se aplicó el riego de germinación; los riegos de auxilio se proporcionaron cuando la humedad en la zona radical del suelo llegó a los niveles definidos para el experimento (0.5, 3.0, 5.5, y 8.0 atm o 80, 35, 20 y 10 %) o cuando el cultivo consumió 20, 65, 80 y 90 % de la humedad aprovechable en el suelo. Los volúmenes suministrados fueron los requeridos para reponer el agua consumida por el cultivo. Los riegos se suspendieron cuando el cultivo alcanzó la madurez fisiológica. La humedad en el perfil del suelo se estimó, de forma simultánea, con el método gravimétrico y con un aspersor de neutrones. En las parcelas seleccionadas para este fin (parcelas útil), se muestreó la humedad por estratos de 20 cm (en el caso del aspersor de neutrones no se realizó muestreo en los primeros 20 cm del suelo).
]]> Durante el desarrollo del cultivo se midió el área foliar para obtener el índice de área foliar (IAF), cada 15 d. Para no destruir las plantas se utilizó el procedimiento de correlacionar el producto obtenido de multiplicar el largo por el ancho de la hoja con mediciones del área foliar obtenido por medio de un integrador de áreas. Las variables de respuesta fueron: rendimiento de grano (t ha–1), rendimiento total de materia seca (t ha–1), contenido de proteína en grano (%), contenido de extracto en el grano (%), población final de tallos (No. de tallos al final del ciclo), altura final de plantas (cm), longitud de espigas (cm) y peso de 100 granos (g).Experimento de coquia en parcelas
Hoechst (1992)[4] realizo un experimento con co–quia (Kochia scoparia L. Schrad) usando semilla proveniente de Vega, Texas, EE.UU. Antes de trazar las parcelas experimentales se realizó un barbecho, un rastreo y una nivelación del suelo. Cada parcela experimental fue de 6 mx9 m, con las calles entre parcelas de 2 m. La siembra se efectuó en forma manual a "chorrillo", con un promedio de trescientos gramos de semilla en cada parcela. En todas las parcelas se aplicaron tres dosis de N: 0 kg ha–1, 60 kg ha–1 y 120 kg ha–1 (como nitrato de amonio y urea); y 80 kg ha–1 de P (como superfosfato de calcio triple). La fertilización se realizó en bandas sobre el surco, antes de la siembra y por única vez. El fertilizante se cubrió con tierra después de aplicarlo. El control de malezas se realizó manualmente. Los riegos se aplicaron usando una lamina de 33.3 mm de agua durante diferentes tiempos de acuerdo a las fecha de siembra, para mantener una humedad homogénea. Para cada fecha de siembra se mantuvo el cultivo a base de riegos hasta que se estableció el temporal de lluvias.
Las variables biológicas fueron: peso húmedo de la planta, peso seco de la planta, altura de la planta, duración de periodos de desarrollo, fecha de floración y producción de semilla.
Se utilizó un diseño experimental de parcelas divididas, considerando los siguientes factores: fechas de siembra, dosis de fertilización y fuente de N. Para el factor de fechas de siembra se escogieron 6 niveles: 6–III–90, 15–III–90, 30–III–90, 16–IV–90, 30–IV–90 y 15–V–90. Para el factor de dosis de fertilización se consideraron tres niveles: 0, 60 y 120 kg ha–1. Para las fuentes de N se evaluaron dos fuentes: urea y nitrato de amonio.
Resultados de los experimentos
Para el experimento con tomate en invernadero, en la Figura 7a se muestra los resultados de las intersecciones y pendientes ajustadas a los diferentes niveles de estrés para el caso de la biomasa fresca aérea, y en la Figura 7b para el área foliar (AF). Los resultados son compatibles con los planteamientos realizados.
En la Figura 8 se muestran los ajustes realizados sobre el cultivo de cebada. En la Figura 8a se muestra algunas curvas de crecimiento asociadas a diferentes niveles de Humedad Aprovechable Radicular (HAR), donde las fases lineales se analizaron usando regresiones lineales por falta de datos para ajustar el modelo expo–lineal. En la Figura 8b se muestra la relación entre la productividad (rendimiento en biomasa) y la pendiente b estimada para la fase lineal del crecimiento.
Partiendo de que las relaciones entre a y b son lineales para dos variables V, se puede analizar las razones entre estas variables, que deberán tener un patrón lineal. En la Figura 9a muestra la razón entre las constantes de las líneas rectas para B seca total y el AF para el caso del tomate y la Figura 9b para B seca total y IAF para cebada.
Finalmente, en la Figura 10a se presenta la relación entre las pendientes de un cultivo de coquia para la B seca total (t ha–1). En la Figura 10b se muestra la relación entre las constantes A y B, similares a las de la Figura 3b, que definen los parámetros AA y BB del espacio supra–paramétrico.
]]>CONCLUSIONES
La evidencia experimental presentada soporta la propuesta del concepto de equivalencia ambiental de la productividad de la vegetación. La equivalencia permite una compactación extrema, dentro del espacio supra–paramétrico, de la variabilidad de los patrones del crecimiento observados en la vegetación.
El esquema de modelación introducido en este trabajo usa las propiedades asociadas a un conjunto de líneas rectas, que se puede expandir a un conjunto de curvas de cualquier complejidad, para caracterizar los patrones del crecimiento de la etapa vegetativa de la vegetación. La hipótesis subyacente a este esquema de modelación simplificado es que los factores de estrés y de manejo, dejando el medio ambiente o fecha de siembra fijo, inciden poco en los patrones iniciales del crecimiento (exponencial), ya que éste está limitado principalmente por el ambiente (radiación fotosinteticamente activa y temperatura) y la capacidad de asimilación de fotosintatos de la vegetación (propiedades ópticas y coeficiente de extinción foliar). Los factores no ambientales tienen un efecto pequeño que difícilmente es detectable en los experimentos, además de que en términos de modelación su impacto es prácticamente despreciable. Para los ambientes (fechas de siembra) variables, el recentrar las curvas de crecimiento (llevarlas a un origen común) hace equivalente los factores de estrés y de manejo con los ambientales. En ambos casos el efecto en la productividad de la vegetación se manifiesta en un cambio de pendiente en las fases lineales de la etapa vegetativa. Bajo el argumento de utilizar periodos de crecimiento más o menos similares, particularmente desde la emergencia al pico del crecimiento, la relación entre la pendiente y la productividad es simple consecuencia del teorema de Pitágoras.
La ventaja del concepto de equivalencia ambiental introducido es que es posible caracterizar un número infinito de patrones del crecimiento de un tipo de vegetación utilizando un mínimo de datos, al menos para la construcción de un sistema de indicadores relativos a un valor de referencia, asociados a combinaciones de ambientes y factores de estrés y de manejo. Por ejemplo, si se utiliza una cámara de crecimiento controlada para definir el potencial genético de crecimiento (usando el valor de 1.0 para esta condición, entonces todas las demás tendrán un valor entre 0 y 1) de un cultivo, esto establece un punto de referencia general para analizar cualquier otra situación. Si recentramos las curvas de crecimiento para tener un origen común y lo trasladamos al punto (0, 0) del espacio tiempo–variable del crecimiento, entonces el punto de referencia define totalmente la condición del ambiente optimo y todos los niveles de estrés y manejo asociados (línea recta que une el punto de origen con el determinado en el medio ambiente optimo). Asimismo, esta situación define la pendiente máxima de los patrones de productividad del cultivo (Figura 6a). Bajo este esquema de modelación, un ciclo cualquiera del crecimiento (con su variable biofísica asociada) define un punto en el espacio paramétrico de la Figura 6a y, por tanto, podemos determinar su línea recta uniendo este punto con el origen. Así, el punto particular analizado puede referenciarse a la recta del potencial genético y determinarse un indicador de su crecimiento (0–1), que al compararse con el valor absoluto del máximo potencial genético define su productividad. Evidentemente hay un número prácticamente infinito de combinaciones de ambientes y factores de estrés y manejo que dan el mismo resultado (equivalencia ambiental).
En otros trabajos de los autores se presenta evidencia experimental, de campo y con sensores remotos, de que el concepto de equivalencia ambiental es aplicable a pastizales, matorrales y bosques/selvas, generalizando el concepto a medios heterogéneos y multi–especies.
LITERATURA CITADA
Ferrandino, F. J. 1989. Spatial and temporal variation of a defoliating plant disease and reduction in yield. Agric. Forest Meteorol. 47: 273–289. [ Links ]
]]>Goudriaan, J. 1994. Using the expolinear growth equation to analize resource capture. In: Monteith, J. L., R. K. Scott, and M. H. Unsworth (eds). Resource Capture by Crops. Nottingham University Press. Nottingham. pp: 99–110. [ Links ]
Goudriaan, J., and J. L. Monteith. 1990. A mathematical function for crop growth based on light interception and leaf area expansion. Annals Bot. 66: 695–701. [ Links ]
Huete, A. R., R. D. Jackson, and D. F. Post. 1985. Spectral response of a plant canopy with different soil backgrounds. Remote Sensing of Environ. 17: 35–53. [ Links ]
Hunt, R. 1982,. Plant Growth Curves. Edward Arnold. London. 248 p. [ Links ]
Jones, C. A., and J. R. Kiniry. 1986. CERES–Maize. A Simulation Model of Maize Growth and Development. Texas A & M. University Press. College Station, Texas. [ Links ]
]]>Lieth, J. H., P. R. Fisher, and R. D. Heins. 1996. A phasic model for the analysis of sigmoid patterns of growth. Acta Horticulturae 417: 113–118. [ Links ]
Monteith, J. L. 2000. Fundamentals equations for growth in uniform stands of vegetation. Agric. Forest Meteorol. 104: 5–11. [ Links ]
Spitters, C. J. T., H. V. Keulen, and D. W. G. V. Kraalingen. 1989. A simple and universal crop growth simulator SUCROS87. In: Rabbinge, R., S. A. Ward and H. H. van Laar (eds). Simulation and Systems Management in Crop Protection. Simulation Monograph. PUDOC. Wageningen, The Netherlands. pp: 147–181. [ Links ]
Steiner, A. A. 1984. The universal nutrient solution. In: Proccedings 6th International Congress on Soilles Culture. Wageningen, The Netherlands. pp: 633–650. [ Links ]
Stockle, C. O., M. Donatelli, M., and R. Nelson. 2003. CropSyst, a cropping systems simulation model. Eur. J. Agron. 18: 289307. [ Links ]
]]>Yin, X., J. Goudriaan, E. A. Latinga, J. Vos, and H. J. Spiertz. 2003. A flexible sigmoid function of deterministic growth. Annals Bot. 91: 361–371. [ Links ]
Yuan, F. M., and W. Bland. 2004. Light and temperature modulated expolinear growth model for potato (Solanum tuberosum L.). Agric. Forest Meteorol. 121: 141–151. [ Links ]
1 Paz, F.2006. Hacia una teoría general de modelación de la dinámica de la vegetación. Reporte septiembre para AGROASEMEX. 43 p.
2 Solorio, N. 2007. Modelo semi–empírico de simulación del crecimiento del cultivo tomate (Lycopersicum esculentum Mill) en invernadero, bajo restricción hídrica y de nitrógeno, Tesis de Maestría, Hidrociencias, Colegio de Postgraduados, Montecillo, México. 62 p.
3 Romero, M. E. 1987. Respuesta de la cebada (Hordeum vulgare L.) a variaciones de la humedad en diferentes etapas de crecimiento. Tesis de Maestría. Colegio de Postgraduados. Montecillo, México. 181 p.
4 Hoechst, M. C. 1992. Efecto de las fechas de siembra y fertilización sobre el crecimiento de Kochia scoparia (L.) Schrad. Tesis de Maestría, Colegio de Postgraduados, Montecillo, México. 93 p.
]]>