
Intervarietal heterosis in saladette tomato (Solanum lycopersicum L.)
Juan Martínez-Solís; Aureliano Peña-Lomelí; Juan Enrique Rodríguez-Pérez; Jaime Sahagún-Castellanos; Margarita Gisela Peña-Ortega*
Universidad Autónoma Chapingo, Departamento de Fitotecnia. km 38.5 Carretera México-Texcoco, Chapingo, Estado de México, MÉXICO. C. P. 56230. Tel. y Fax 01 5959521642. Correo-e: mgise@excite.com (*Autor para correspondencia).
Recibido el 7 de febrero, 2012.
Aceptado el 5 de febrero, 2013.
Resumen
Con el objetivo de evaluar la utilidad de híbridos de jitomate como progenitores potenciales para un programa de mejoramiento, dos diseños dialélicos 5 x 5 (Griffing II) fueron desarrollados usando dos tipos de jitomate saladette de hábito determinado e indeterminado. Para la variable peso de fruto (PF), el híbrido DG05 de hábito determinado mostró buena aptitud combinatoria específica (ACE) y alta heterosis intervarietal (HI), mientras que el híbrido DG03 tuvo alta aptitud combinatoria general (ACG); por lo que ambos híbridos tienen buen potencial para ser utilizados como materiales base para programas de mejoramiento por hibridación y selección, respectivamente. Para los materiales de hábito indeterminado, IG01 y IG04 mostraron los valores más altos de ACG así como buena ACE entre ellos para PF; por lo que ambos podrían ser utilizados en programas de mejoramiento por selección e hibridación.
Palabras clave: Diseños dialélicos, aptitud combinatoria, mejoramiento vegetal, cruzas, heterosis específica.
Abstract
In order to evaluate the usefulness of tomato hybrids as potential progenitors for a breeding program, two 5 x 5 diallel designs (Griffing II) were developed using two types of saladette tomato: determinate and indeterminate growth habits. For fruit weight per plant (FW), the determinate growth habit hybrid DG05 showed good specific combining ability (SCA) and high intervarietal heterosis (IH), while DG03 hybrid had high general combining ability (GCA); therefore, both hybrids have good potential for use as foundation parents in hybridization and selection breeding programs, respectively. For the saladette indeterminate growth habit type, IG01 and IG04 hybrids showed the highest GCA values in addition to good SCA between them for FW; therefore, they could be used for both selection and hybridization breeding programs.
Keywords: Diallel designs, combining ability, plant breeding, crosses, specific heterosis.
INTRODUCCIÓN
]]> En los últimos años el consumo mundial per capita de jitomate (Solanum lycopersicum L.) en fresco o procesado se ha incrementado significativamente, haciendo de este cultivo el segundo vegetal más importante a nivel mundial. En consecuencia, en varios países también se ha incrementado la producción agrícola, en algunos de ellos sobre todo en condiciones de invernadero (DeGiglio, 2003).De la inversión total necesaria para cultivar jitomate, una parte importante (aproximadamente el 20 %) (Martínez et al., 2005) se utiliza en la compra de semilla importada de variedades comerciales, producida principalmente por compañías transnacionales que la venden a altos precios. Por lo que algunos programas de investigación institucionales financiados públicamente han iniciado programas de mejoramiento genético de jitomate con el objetivo de desarrollar variedades e híbridos que puedan ser usados por los productores locales a bajo costo.
Los tipos de progenitores que se pueden usar para un programa de mejoramiento genético incluyen poblaciones de amplia base genética, tales como las variedades locales o las colecciones de germoplasma, de las cuales a través de la selección recurrente se pueden obtener variedades mejoradas. Además, las variedades comerciales tales como los híbridos simples y las variedades sintéticas podrían ser utilizadas para diversificar la reserva genética inicial y como población base (Kannenberg y Falk, 1995).
Para cultivos de grano como maíz, la utilización de semillas provenientes de cruzas entre híbridos comerciales ha sido recomendada como una alternativa más económica para productores maiceros debido al menor costo de la semilla y buenos niveles de producción de grano ocasionados por la heterosis residual (Villanueva et al., 1994).
Con objeto de poder identificar materiales con buen potencial para ser usados como progenitores base para un programa de mejoramiento genético, los métodos de análisis estadístico y genético permiten la estimación de parámetros genéticos que a su vez definen el tipo de acción génica más importante controlando el carácter bajo consideración (Vega, 1988). Los genes con efectos aditivos y de dominancia son esenciales en el control genético de caracteres de importancia agronómica, pero para propósitos de mejoramiento, la estimación de la contribución relativa de ambos tipos de acción génica es crucial para decidir la estrategia a usar, selección recurrente o hibridación, para alcanzar los mejores resultados en el desarrollo de productos de mejoramiento (Lynch y Walsh, 1998).
Los diseños de apareamiento han sido ampliamente utilizados para obtener información genética valiosa para los fitomejoradores (Gardner y Eberhart, 1966). Griffing (1956) propuso cuatro distintos diseños de apareamiento, algunos consideran la existencia de efectos maternos y por tanto incluyen las cruzas directas y recíprocas, mientras que los diseños II y IV incorporan la evaluación de poblaciones autofecundadas (Lynch y Walsh, 1998).
Aun cuando a la fecha las bases teóricas de la heterosis han permanecido inciertas, la falta de un conocimiento preciso acerca de este fenómeno no ha evitado la explotación del vigor híbrido en la producción de alimentos (Reeves et al., 1999; Birchler et al., 2006). El término heterosis fue acuñado por Shull (1948) como "... la interpretación del mayor vigor, tamaño, fructificación, velocidad de desarrollo, resistencia a plagas y enfermedades o a factores climáticos adversos de cualquier tipo, expresado por los organismos producto de cruzamiento en comparación con sus progenitores endogámicos, como resultado de la disimilitud genética de los gametos parentales", conocida como la teoría de la sobredominancia. Otra explicación la ternativa de la heterosis establece que el grado de heterosis obtenido está en función directa del número de loci dominantes; esta teoría explica tanto la heterosis como el fenómeno contrario de la depresión endogámica causada por consanguinidad. Esta teoría es conocida como la teoría de la dominancia (Lamkey y Edwards, 1999).
Jones (1917) también propuso una teoría para explicar la heterosis, es decir, que un gran número de genes son responsables de la diferenciación de la mayoría de los caracteres cuantitativos y que los genes favorables para la mayor expresión de dichos caracteres son al menos parcialmente dominantes (Lynch y Walsh, 1998).
El análisis de resultados que incluyen un amplio rango de materiales, cultivos y ambientes ha demostrado marcadas diferencias en el comportamiento agronómico, y en consecuencia, sobre los niveles de heterosis, haciendo necesaria la utilización de alguna de las dos teorías para la explicación de la heterosis práctica obtenida en los programas de mejoramiento (Falconer y Mackay, 2006).
La hibridación en cultivos autógamos no siempre tiene como objetivo incrementar los rendimientos sino conjuntar caracteres complementarios u obtener uniformidad y reproducibilidad en caracteres productivos importantes (Lamkey y Edwards, 1999). Sin embargo, en el caso del jitomate se han encontrado evidencias de sobredominancia causando heterosis en rendimiento (Kurian et al., 2001; Semes et al., 2006).
]]> Los objetivos de este estudio fueron evaluar los niveles de heterosis intervarietal para cruzas entre híbridos de jitomate de hábito determinado e indeterminado tipo saladette, así como estimar el principal tipo de acción génica controlando el peso de fruto en los materiales evaluados para identificar aquéllos con buen potencial para ser utilizados como progenitores originales dentro de programas de mejoramiento por selección e hibridación.
MATERIALES Y MÉTODOS
Diez híbridos comerciales de jitomate tipo saladette fueron divididos en dos grupos de acuerdo a su hábito de crecimiento: determinado e indeterminado (Cuadro 1). De acuerdo con el diseño II de Griffing se realizaron todas las cruzas simples posibles dentro de cada grupo.
Con el objetivo de realizar las cruzas dialélicas necesarias dentro de cada uno de los grupos por hábito de crecimiento, seis plantas de cada progenitor fueron trasplantadas para ser manejadas bajo condiciones hidropónicas en invernadero en Chapingo, México, durante el otoño de 2002. Las bolsas de plástico fueron colocadas a una distancia de 50 cm alineadas en hileras de 1.4 m de ancho y se realizaron todas las posibles cruzas controladas entre ellas. Las cruzas artificiales fueron realizadas usando el método de Pérez et al. (1998).
Los dos dialélicos incluyendo los cinco progenitores y sus quince cruzas, uno por cada hábito de crecimiento, fueron evaluados bajo condiciones de hidroponía en invernadero durante la primavera de 2003, bajo un diseño de bloques completos al azar con diez repeticiones. La unidad experimental estuvo constituida por una bolsa de plástico negro de diecisiete litros de capacidad llenada con tezontle fino como sustrato. En ambos experimentos las bolsas de plástico fueron colocadas a 50 cm entre plantas y 75 cm entre hileras. Las plantas fueron trasplantadas treinta y tres días después de la siembra; las plantas de hábito indeterminado fueron mantenidas a un tallo y no más de seis racimos, mientras que las plantas de hábito determinado fueron manejadas a dos tallos con tres racimos cada uno. Todas las plantas fueron tutoreadas con rafia a partir de que alcanzaron 50 cm de altura.
El riego fue automatizado y proporcionó dos riegos por día (9:00 y 11:00 h) de aproximadamente quince minutos cada uno. Durante las primeras dos semanas después del trasplante, la solución nutritiva, preparada de acuerdo a lo propuesto por Pérez y Castro (1999), fue suministrada diluida al 50 %; posteriormente, la solución se aplicó al 100 % manteniendo esta condición hasta el término del cultivo. La cosecha se realizó cuando el fruto alcanzó su madurez comercial en cortes con espaciamientos de 7 a 10 días entre cada uno, en los que se evaluó: Peso total de fruto por planta en gramos (PT); Número total de frutos por planta (FT); Peso promedio de fruto en gramos (PPF), obtenido como el cociente del peso total de fruto por planta entre el número total de frutos por planta.
Para cada dialélico, de acuerdo con el modelo de efectos fijos del diseño dialélico II de Griffing (1956), se realizaron los análisis de varianza para estimar los efectos de aptitud combinatoria general (ACG) y aptitud combinatoria específica (ACE) para cada carácter, usando el programa para SAS propuesto por Zhang y Kang (1997). Los datos también fueron sometidos a una prueba de comparación de medias de Tukey (P ≤ 0.05) y se realizaron pruebas de contrastes ortogonales entre las cruzas y el mejor progenitor así como con respecto al promedio de progenitores.
]]> La heterosis se estimó usando el modelo II de Gardner y Eberhart (1966) y Gardner (1967) correspondiente al diseño dialélico II de Griffing con cruzas entre poblaciones, de la siguiente forma.
Donde:
Yij= media del progenitor cuando i = j y media de la cruza cuando i ≠ j
μv = = promedio de progenitores
= promedio de progenitores
Vi= efecto varietal de la j ésima variedad
Vj= efecto de la variedad en la j ésima variedad
θ = cero cuando i = j; θ = 1 cuando i ≠ j
hij = efecto de heterosis en la cruza entre variedades i y j

 = heterosis media
= heterosis media
hi = heterosis varietal para la variedad i
hj = heterosis varietal para la variedad j
Sij = heterosis específica para la cruza entre las ¡Hvariedades i y j

Donde:
 = media de todas las cruzas;
= media de todas las cruzas;
= media de todos los progenitores

n = número de progenitores
 = media de las cruzas del progenitor i;
= media de las cruzas del progenitor i;
= media de todas las cruzas;
Yi = media del progenitor i;
YV= media de todos los progenitores.
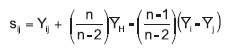
Donde:
Yij = media de cada cruza ij;
n = número de progenitores = media de todas las cruzas;
]]> = media de todas las cruzas; = media de las cruzas del progenitor i;
 = media de las cruzas del progenitor j;
= media de las cruzas del progenitor j;
RESULTADOS Y DISCUSIÓN
Los resultados del análisis de varianza (Cuadro 2) muestran que las cruzas presentaron un efecto significativo en todos los caracteres evaluados con excepción del número de frutos totales en los genotipos tipo saladette de hábito determinado. La descomposición de esos efectos mostró que la variación atribuida a la ACG fue diferente de cero para peso total de fruto (PT) en ambos grupos y para peso promedio de fruto (PF) sólo para los materiales tipo saladette determinado (SD). Los efectos de la ACE sólo fueron significativos para PPF en los tipos indeterminados.
Los coeficientes de variación fueron altos para FT y PPF, superiores al 26 %, lo cual refleja la alta variabilidad en tamaño de fruto presente dentro del mismo progenitor; sin embargo, la variación observada para rendimiento por planta (PT) tuvo un coeficiente muy pequeño.
Saladette Determinado
Los progenitores y sus cruzas se presentan en el Cuadro 3. Para la producción total por planta PT, aunque la cruza 12 (3x5) rindió estadísticamente lo mismo que sus padres, superó a las cruzas 4 (1x4) y 12 (4x5), así como al híbrido GD04.
]]> Para el resto de los caracteres evaluados, frutos por planta (FT) y peso promedio de fruto (PPF), desde el punto de vista estadístico no se presentaron diferencias que representen una tendencia obvia debido a que sólo dos cruzas, cruza 9 (2x5) y cruza 4 (1x4), se comportaron de manera distinta al resto (a = 0.05). Estos resultados indican que este grupo de progenitores no es útil para producir cruzas que superen el comportamiento de sus progenitores.El progenitor GD04 presentó el peor comportamiento para PT y PPF, lo cual como consecuencia afectó negativamente el rendimiento observado en las cruzas en que participó este progenitor.
Cuando los progenitores se analizaron por separado (Cuadro 4), la mayor estimación de ACG en PT y FT fue en GD03, mientras que para PPF el progenitor GD02 tuvo el mejor comportamiento. De acuerdo a la tendencia observada para las medias obtenidas, GD04 tuvo los menores valores de ACG en PT y FT, mientras que GD02 presentó el menor valor en PPF. A partir de estos resultados, GD03 y GD01 podrían ser considerados como progenitores potenciales para un programa de mejoramiento por selección, debido a su alta ACG y la alta heterosis observada para PT. La falta de significancia obtenida en la heterosis media pudo deberse a que su estimación involucró las cruzas con el progenitor GD04, las cuales exhibieron el peor comportamiento, por lo tanto resultando en una baja heterosis varietal para este progenitor.
Las cruzas con altos valores de ACE para PT y PPF fueron cruza 12 (3x5), cruza 9 (2x5) y cruza 5 (1x5); las últimas dos cruzas mostraron valores más altos de ACE que de ACG con respecto a sus respectivos progenitores (Cuadro 5).
El progenitor GD05 participó en todas las tres cruzas previamente mencionadas, las cuales estuvieron dentro de las mejores cruzas para PT y PPF (Cuadro 3). Estos resultados sugieren que este progenitor podría tener buen potencial para ser utilizado en un programa de mejoramiento por hibridación.
Para FT, la cruza 11 (3x4) obtuvo el más alto valor de ACE, superior a los valores observados de ACG de sus progenitores (Cuadro 5); sin embargo, este parámetro genético no estuvo asociado a una alta producción de frutos por planta probablemente debido a que los valores de ACG y ACE para PPF fueron los menores.
La heterosis con respecto al progenitor medio (Cuadro 6) fue positiva en seis de las cruzas para PT, donde las cruzas 5 (1x5), 9 (2x5) y 12 (3x5) mostraron los más altos valores: 19.4, 24.3 y 18.2 por ciento, respectivamente; no obstante, de acuerdo con los contrastes ortogonales entre las cruzas y el progenitor medio sólo la cruza 9 (2x5) fue significativamente diferente.
En el caso de la variable frutos totales por planta, con excepción de la cruza 3 (1x3), todas las cruzas presentaron estimaciones positivas de heterosis y en el caso de la cruza 11 (3x4) el contraste ortogonal entre la cruza y el progenitor medio fue significativamente diferente de cero. En relación al PPF, cuatro cruzas mostraron estimaciones positivas de heterosis absoluta ( ), donde la cruza 12 (3x5) presentó 16.5 % de heterosis. Por el contrario, la cruza 4 (1x4) fue inferior en comparación a las medias de sus progenitores (Cuadro 6).
), donde la cruza 12 (3x5) presentó 16.5 % de heterosis. Por el contrario, la cruza 4 (1x4) fue inferior en comparación a las medias de sus progenitores (Cuadro 6).
La heterosis absoluta (HAmp) respecto al mejor progenitor fue positiva en tres cruzas para PT y PPF (Cuadro 7); los contrastes ortogonales entre las cruzas y el mejor progenitor fueron negativos y significativos sólo para las cruzas 4 (1x4) y 11 (3x4).
En el número total de frutos por planta (FT) no hubo expresión de heterosis significativa aun cuando se obtuvieron valores positivos en cinco de las cruzas. En resumen, al comparar ambas estimaciones de heterosis, respecto al progenitor medio y al mejor progenitor, todas las cruzas donde participó GD05 mostraron los mayores valores positivos. Estos resultados sugieren que este progenitor tiene buen potencial para ser integrado dentro de la base genética de un programa de mejoramiento por hibridación, tanto directamente como progenitor de híbridos como fuente para derivar líneas puras.
]]> No hubo efectos de heterosis específica para ninguno de los caracteres evaluados (Cuadro 8). Estos resultados podrían deberse a la alta frecuencia de loci en condición heterocigótica y/o a la falta de diversidad genética entre los progenitores evaluados (Villand et al., 1998).Cuando los estimadores de heterosis respecto al progenitor medio () y respecto al mejor progenitor (HAmp), así como los valores obtenidos de aptitud combinatoria específica son considerados simultáneamente se puede concluir que las mejores cruzas para PT fueron cruza 5 (1x5), cruza 9 (2x5) y la cruza 12 (3x5). A partir de estos resultados puede concluirse que el progenitor GD05 está contribuyendo su superioridad genética a estas cruzas, por lo que consecuentemente tuvo también un alto valor de ACG (7.65) (Cuadro 4). No obstante, si se consideran simultáneamente la aptitud combinatoria general (ACG) y la heterosis intervarietal (HV), los progenitores GD01 y GD03 también son valiosos.
Para producción total de frutos por planta (PT) y peso promedio de fruto (PPF), GD05 también presentó buenos valores de ACE, HAmp, y HE en las cruzas 5 (1x5) y 9 (2x5). Por lo que el progenitor GD05 tiene buen potencial para ser utilizado como progenitor base para un programa de mejoramiento genético por selección.
Por otro lado, GD03 tuvo los valores más altos de ACG y HV para PT, resultados que aparentemente lo sugerirían como un buen progenitor de híbridos, sin embargo las estimaciones de ACE, HAmp, y HE permiten concluir que este progenitor sólo combinó bien con otros dos progenitores, por lo que tiene un mayor potencial para ser utilizado en un programa de mejoramiento por selección en lugar de uno por hibridación.
Finalmente GD01 tuvo valores aceptables de ACG y HV y con excepción de su cruza con GD05, sus efectos de ACG fueron más importantes que los de ACE, por lo que estos resultados sugieren la conveniencia de utilizarlo en un programa de selección recurrente con objeto de explotar la acción génica de aditividad presente.
En relación a FT, sólo GD03 mostró un valor significativo de ACG así como una buena estimación de HV. Sin embargo, las ACE para las cruzas 3 (1x3) y 7 (2x3) fueron negativas, por tanto causando bajas estimaciones de HAmp y HE. Por lo que GD03 podría ser una buena fuente para seleccionar genes favorables con efectos aditivos para FT, ya que produjo más frutos por planta tanto por sí mismo como en algunas de sus cruzas [11 (3x4) y 12 (3x5)].
Para la variable peso promedio de fruto (PPF), el valor más alto de ACG fue para GD02, sin embargo la HV negativa influenció las otras estimaciones de heterosis, por lo que sólo presentó valores positivos en combinación con GD05. En el Cuadro 5 puede observarse que los efectos de ACE de este progenitor fueron menores a los de ACG para la mayoría de sus cruzas, sugiriendo por tanto que en este progenitor la acción génica aditiva en el control genético del PPF es más importante que la acción génica de dominancia, por tanto sugiriendo que este material podría tener un mejor uso dentro de un programa de mejoramiento por selección.
En contraste, la estimación de ACG en GD04 para tamaño de fruto fue significativa pero negativa, reflejando con ello un pobre comportamiento en casi todas sus cruzas, en consecuencia con bajas estimaciones de ACE, HAmp, y HE, lo cual conduce a evaluar este progenitor como el de peor comportamiento entre los materiales evaluados en este estudio.
Variedades tipo saladette de hábito indeterminado
]]> Las medias de las quince cruzas entre los 5 progenitores considerados dentro del dialélico II de Griffing se presentan en el Cuadro 9. Para la producción total por planta (PT) en su mayoría no hubo diferencias significativas entre los progenitores y sus cruzas, con excepción de la cruza 9 (2x5) que mostró la producción más baja. Para número de frutos totales por planta (FT) se observó un patrón similar, donde sólo el progenitor GI03 tuvo significativamente menos frutos por planta que el resto de los progenitores y sus cruzas, mientras que para peso promedio de fruto (PPF) no hubo diferencias significativas.El comportamiento de los cinco progenitores evaluados se presenta en el Cuadro 10. No se encontraron diferencias significativas para peso promedio de fruto entre ellos y aun cuando el progenitor GI04 tuvo la más alta producción total por planta (PT) no fue estadísticamente diferente del resto, con excepción de GI03, el cual tuvo la menor producción debido a un reducido número de frutos (FT).
La aptitud combinatoria general de GI04 fue significativa para la producción total por planta sugiriendo la existencia de genes favorables con efectos aditivos en este material. Sin embargo, para frutos totales por planta fueron los progenitores GI01 y GI05 los que presentaron efectos significativos de ACG, donde fueron mayores en GI01. Por lo que estos resultados sugieren que los progenitores GI01 y GI04 podrían ser buenos candidatos para ser usados como progenitores base dentro de un programa de selección recurrente para aprovechar sus efectos aditivos para rendimiento. En relación al peso promedio de fruto, no resultó sorprendente que GI03 fue el único progenitor que tuvo efectos significativos de ACG, ya que produjo el menor número de frutos por planta y frutos más grandes que los que se obtuvieron del resto de los híbridos.
Las más altas y positivas estimaciones de heterosis varietal obtenidas para PT y PPF correspondieron a GI05, mientras que GI04 mostró el mayor valor para FT. Las estimaciones de heterosis media no fueron significativas para ninguno de los caracteres evaluados. Estos resultados reflejan la tendencia discutida previamente acerca de que en general los progenitores se comportaron mejor que sus cruzas, probablemente debido a que tuvieron una constitución genética similar lo que resultó en la no detección de efectos de vigor cuando se cruzaron entre sí.
Este patrón puede apreciarse claramente en el Cuadro 11, donde sólo las cruzas 4 (1x4), 7 (2x3) y 12 (3x5) mostraron efectos positivos de ACE para PT, sin embargo sólo las cruzas 7 (2x3) y 11 (3x4) presentaron valores superiores a los de las ACG de los progenitores. Para PPF las cruzas 4 (1x4) y 12 (3x5) tuvieron efectos de ACE superiores a los de ACG.
La heterosis con respecto al progenitor medio (Cuadro 12) para PT fue positiva sólo positiva para las cruzas 4 (1x4) y 7 (2x3) representando valores de heterosis porcentual de 2.8 y 5.2, respectivamente. No obstante, aun cuando las cruzas 9 (2x5) y 14 (4x5) presentaron estimaciones negativas de heterosis, fueron las únicas que mostraron heterosis significativa al compararlas con la producción promedio de sus progenitores mediante contrastes ortogonales. Estos resultados podrían explicarse debido a la alta frecuencia de loci heterocigóticos controlando este carácter en ambos progenitores, los cuales segregaron como producto del cruzamiento.
Para el carácter FT las estimaciones de heterosis respecto al progenitor medio fueron positivas para las cruzas 2 (1x2), 7 (2x3) y 11 (3x4), pero sólo las cruzas 7 y 11 mostraron valores de heterosis porcentual mayores de 10. Nuevamente la cruza 4 (1x4) que tuvo una estimación de heterosis negativa fue la única que mostró heterosis significativa cuando las cruzas fueron comparadas con el peso promedio de los frutos de sus progenitores mediante pruebas de contrastes ortogonales. La cruza 4 también presentó heterosis positiva respecto al progenitor medio para PPF, lo cual representó 47.8 % de heterosis. La cruza 12 (3x5) también mostró heterosis (12.5 %) para este carácter.
Cuando se utilizó una estimación más estricta de heterosis, al comparar la cruza contra el mejor progenitor, se observó una tendencia similar (Cuadro 13). Aun cuando las estimaciones de heterosis para la mayoría de las cruzas fueron negativas, las pruebas de contrastes ortogonales mostraron que para producción de frutos por planta (FT) las cruzas 3 (1x3), 8 (2x4), 9 (2x5), 11 (3x4) y 14 (4x5) tuvieron efectos significativos de heterosis. No obstante para frutos totales (FT) la cruza 3 (1x3), 4 (1x4) y 12 (3x5) mostraron efectos de heterosis significativos.
Para el carácter PPF las cruzas 12 (3x5) y 4 (1x4) tuvieron valores positivos de heterosis, pero sólo para la cruza 4 este valor (64.2 %) fue significativo de acuerdo al contraste ortogonal. Cuando se toman en cuenta ambas estimaciones de heterosis, y HAmp, los progenitores GI01 y GI04 podrían ser considerados con buen potencial para ser incluidos como material base dentro de un programa de mejoramiento de hibridación cuyo objetivo sea incrementar el tamaño del fruto, ya que parecen ser una buena fuente de genes favorables con efectos de dominancia.
La determinación de los efectos de heterosis específica mostró que para la producción total por planta las cruzas 4 (1x4), 7 (2x3) y 12 (3x5) tuvieron valores positivos (Cuadro 14); mientras que cuando se considera el número total de frutos por planta sólo la cruza 11 (3x4) presentó una estimación positiva aceptable, mientras que para tamaño de fruto (PPF) las cruzas 12 (3x4) y 4 (1x4) mostraron altos valores de heterosis.
]]> Los resultados obtenidos a partir de este estudio indican que la cruza 4 (1x4) y sus progenitores fueron los más relevantes, ya que mostraron los más altos valores para la mayoría de los parámetros evaluados. Esta cruza en particular superó en producción total al resto y excedió en 9.2 % a la producción de GI01. Adicionalmente, tuvo la más alta AC.01.0E4.011, HE y el segundo valor de. Los progenitores involucrados. GI01 y GI04 también mostraron altos efectos de ACG. No obstante, cuando se consideraron las variables número de frutos producidos por planta y peso promedio de fruto, estos progenitores también tuvieron altas estimaciones de ACG, pero no presentaron efectos significativos de heterosis, lo que sugiere que el tipo de acción génica controlando estos caracteres fue de tipo aditivo. En resumen puede concluirse que los progenitores GI01 y GI04 tienen buen potencial para ser usados como material base de un programa de selección recurrente para explotar el tipo de acción génica presente en estos materiales. Sin embargo es importante considerar la aceptable heterosis observada para PT cuando se cruzaron entre sí, por lo que en este caso se podría proponer el uso de un esquema de mejoramiento que contemple simultáneamente la selección recurrente en ambos materiales de manera separada con el objetivo de obtener poblaciones mejoradas que posteriormente podrían ser utilizadas para derivar líneas que podrían cruzarse entre sí conformando un grupo heterótico en concordancia con el conocimiento previo que se tiene de estos materiales de su alto potencial para producir buenas combinaciones de genes complementarios. Por lo que se podría usar un esquema de selección recíproca recurrente (Kannenberg y Falk, 1995).
CONCLUSIONES
Las estimaciones de aptitud combinatoria general y específica, junto con distintas mediciones de heterosis obtenidas en el presente estudio permitieron la identificación de progenitores prometedores para un programa de mejoramiento genético de jitomate.
Los híbridos de hábito de crecimiento determinado GD03 y GD05 tienen buen potencial para formar parte de las poblaciones base para mejorar la producción total de frutos por planta en programas de mejoramiento tanto por selección como por hibridación, mientras que el progenitor GD01 sería más útil dentro de un programa de selección debido a sus efectos genéticos aditivos.
Entre los híbridos de hábito indeterminado evaluados en este trabajo, GI01 y GI04 son buenas fuentes de genes favorables con ambos tipos de acción génica, aditividad y dominancia, por lo que tienen buen potencial para ser usados tanto de programas de mejoramiento de selección como de hibridación.
LITERATURA CITADA
BIRCHLER, J. A.; YAO, H.; CHUDALAYANDI, S. 2006. Unraveling the genetic basis of hybrid vigor. PNAS 103: 1295712958. DOI:10.1073/pnas.0605627103. [ Links ]
DEGIGLIO, M. A. 2003. Growth of the fresh greenhouse tomato market in the USA. Acta Horticulturae 611: 91-92. http://www.actahort.org/members/showpdf?booknrarnr=611_15 [ Links ]
FALCONER, D. S.; MACKAY, T. F. C. 2006. Introduction to Quantitative Genetics. W. H. Freeman and Company. New York. 469 p. [ Links ]
GARDNER, C. O.; EBERHART, S. A. 1966. Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22:439-452. http://www.jstor.org/discover/10.2307/2528181?uid=3738664&uid=2&uid=4&sid=21101635786561 [ Links ]
GARDNER, C. O. 1967. Simplified methods for estimating constants and computing sums of squares for a diallel cross analysis. Fitotecnia Latinoamericana 4(2): 1-12. [ Links ]
GRIFFING, B. 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal of Biology Sciences. 9: 463-493. http://www.publish.csiro.au/?act=view_file&file_id=BI9560463.pdf [ Links ]
KANNENBERG, L.W.; FALK, D.E. 1995. Models for activation of plant genetic resources for crop breeding program. Canadian Journal of Plant Science 75: 45-53. DOI: 10.4141/ cjps 95-008. [ Links ]
]]>KURIAN, A.; METER, K. V.; RAJAN, S. 2001. Heterosis for yield components and fruit characters in tomato. Journal of Tropical Agriculture 39: 5-8. http://www.jtropag.in/index.php/ojs/article/view/11/7 [ Links ]
LAMKEY, K. R., EDWARDS, J. W. 1999. Quantitative genetics in heterosis. In: The Genetics and Exploitation of Heterosis in Crops. COORS J. G.; PANDEY, S. (eds.). American Society of Agronomy, Inc. & Crop Science Society of America Inc. Madison, Wisconsin, U. S. pp: 31-48. DOI:10.2134/1999. geneticsandexploitation.c4. [ Links ]
LYNCH, M.; WALSH, B. 1998. Genetics and Analysis of Quantitative Traits. Sinauer Associates, Sunderland, Massachusetts. 359 p. [ Links ]
MARTÍNEZ S., J.; PEÑA L., A.; RODRÍGUEZ P., J.E.; VILLANUE-VA V., C.; SAHAGÚN C., J.; PEÑA O., M.G. 2005. Comportamiento productivo en híbridos de jitomate y sus respectivas poblaciones F2. Revista Chapingo Serie Horticultura 11(2): 299-307. www.chapingo.mx/revistas/revistas/articulos/doc/bb05a7468b2edf244395ff9f71f43f02.pdf [ Links ]
PÉREZ G., M.; MÁRQUEZ S., F.; PEÑA L., A. 1998. Mejoramiento Genético de Hortalizas. Ed. Mundi-Prensa. México, D.F. pp 159-161. http://biblioteca.universia.net/html_bura/ficha/params/id/37937722.html [ Links ]
REEVES, T.; PINSTRUP-ANDERSEN, P.; PANDYA-LORCH, R. 1999. Food security and the role of agricultural research. In: The Genetics and Exploitation of Heterosis in Crops. COORS, J. G.; PANDEY, S. (eds.). American Society of Agronomy, Inc. & Crop Science Society of America Inc. Madison, Wisconsin, U. S. pp: 1-5. DOI:10.2134/1999.ge-neticsandexploitation.c1. [ Links ]
SEMEL, Y.; NISSENBAUM, J.; MENDA, N.; ZINDER, M.; KRIEG-ER, U.; ISSMAN, N.; PLEBAN, T.; LIPPMAN, Z.; GUR, A.; ZAMIR, D. 2006. Overdominant quantitative trait loci for yield and fitness in tomato. PNAS 103: 12981-12986.DOI:10.1073/pnas.0604635103. [ Links ]
SHULL, G. H. 1948. What is "heterosis"? Genetics 33: 439-446. http://www.genetics.org/content/33/5/439.full.pdf [ Links ]
VEGA O., P. C. 1988. Introducción a la Teoría de la Genética Cuantitativa: con Especial Referencia al Mejoramiento de Plantas. Ed. Ediciones de la Biblioteca Central de Venezuela. pp 140-156. [ Links ]
VILLAND, J.; SKROCH, P. W.; LAI, T.; HANSON, P.; KUO, C. G.; NIENHUIS, J. 1998. Genetic variation among tomato accessions from primary and secondary centers of diversity. Crop Science 38: 1339-13347. DOI:10.2135/cropsci1998.0 011183X003800050032x. [ Links ]
VILLANUEVA V., C.; CASTILLO G., F.; MOLINA G., J.D. 1994. Aprovechamiento de cruzamientos dialélicos entre híbridos comerciales de maíz: análisis de progenitores y cruzas. Revista Fitotecnia Mexicana 17: 175-185. [ Links ]
ZHANG, Y.; KANG, M. S. 1997. Diallel-SAS: A SAS program for Griffing's diallel analyses. Agronomy Journal 89(2): 176182. DOI:10.2134/agronj1997.00021962008900020005. [ Links ]
]]>