 = es la media de las n variedades parentales;
= es la media de las n variedades parentales;
Heterosis in hybrids of squash type Grey Zucchini
César Sánchez-Hernández1*; Clemente Villanueva-Verduzco2; Jaime Sahagún-Castellanos2; Juan Porfirio Legaria-Solano2; Juan Martínez-Solís2; Miguel Ángel Sánchez-Hernández3; Jorge Alfredo Ortíz-Quintero4; Salomón López-Serrano4
1 Universidad de la Cañada. Teotitlán de Flores Magón, Oaxaca. México. C. P. 68540. Autor para correspondencia: cesarsh79@hotmail.com.
2 Universidad Autónoma Chapingo. Departamento de Fitotecnia. km 38.5 carretera México-Texcoco. Chapingo, Estado de México. México. C. P. 56230.
3 Universidad del Papaloapan. Av. Ferrocarril Hidalgo s/n. Ciudad Universitaria Loma Bonita, Oaxaca. México. C. P. 68400.
]]> 4 Instituto Tecnológico Superior de Cosamaloapan. Av. Tecnológico s/n. Col. Los Ángeles Cosamaloapan, Veracruz. México. C. P. 95400.
Recibido: 15 de febrero, 2011
Aceptado: 07 de marzo, 2013
Resumen
Puesto que los híbridos comerciales pueden usarse exitosamente en programas de mejoramiento genético, el objetivo del presente trabajo fue identificar híbridos comerciales con valor genético para producir híbridos y variedades de alto rendimiento. Se evaluaron ocho híbridos de calabacita (Cucurbita pepo L.) tipo Grey Zucchini y sus 56 cruzas posibles en Chapingo, México durante 2007 y 2008. Con el modelo II de Gardner y Eberhart (1966) se estimó heterosis (media, varietal, específica) y con respecto al mejor progenitor. Los resultados indicaron que hubo heterosis media positiva en todos los caracteres estudiados (REN: 1.10 t·ha−1; RPP: 13.10 g·planta−1; AFR: 0.10 cm, LFR: 0.50 cm, DFF: 3 días y DFM: 3 días). El híbrido comercial 'Terminator' manifestó efectos positivos favorables de heterosis varietal en rendimiento y sus componentes (RPH: 3.62 t·ha−1, RPP: 63.37 g·planta−1, FPP: 0.73 frutos, LFR: 0.33 cm, AFR: 0.20 cm, DFF: 1.66 días y DFM: 1.67 días). Los mejores progenitores para rendimiento serían 'Terminator' y 'Lolita', mismos que mostraron mayor heterosis varietal. Las mejores cruzas entre híbridos fueron 'Grey Zucchini M+M' x 'Terminator' (3x7) (Sij 3.78**); 'Hurakán' x 'Lolita' (5x4) (Sij 3.86**), y 'Tala' x 'Terminator' (1x7) (Sij 3.25**), las cuáles podrían utilizarse en un programa de mejoramiento genético para explotar los efectos de dominancia mediante hibridación cíclica. Las tres cruzas más precoces, con 44 días a floración femenina, fueron 'Lolita' x 'Tala' (4x1), 'WA9041' x 'Tala' (6x1), 'Dolarzini' x 'Tala' (8x1) y la más tardía fue la cruza 'Terminator' x 'Hurakán' (7X5) (54 días).
Palabras clave: Cucurbita pepo L., análisis dialélico, rendimiento de fruto.
Abstract
]]> The aim of the present study was to identify commercial hybrids with genetic value to produce high yield hybrids or varieties because commercial hybrids can be used with success in breeding programs. A total of eight commercial hybrids of squash (Cucurbita pepo L.) type Grey Zucchini and their 56 possible crosses were evaluated at Chapingo, Mexico during 2007 and 2008. Heterosis (average, varietal, specific) was estimated using the model II of Gardner and Eberhart (1966) and with respect to the best parent. Results showed positive average heterosis for all the characters (YH: 1.10 t·ha−1; YPP: 13.10 g·plant−1; FRW: 0.10 cm; FRL: 0.50 cm; DFF: 3 days and DMF: 3 days) studied. The hybrid 'Terminator' showed favorable positive effects of varietal heterosis for yield and components (RPH: 3.62 t·ha−1; YPP: 63.37 g·plant−1; FPP: 0.73 fruits, FRL: 0.33 CM, FRW: 0.20 CM, DFF: 1.66 days and DMF: 1.67 days). The best parents for yield were 'Terminator' and 'Lolita' because they showed greater varietal heterosis. The best crosses among hybrids were 'Grey Zucchini M+M' x 'Terminator' (3x7) (Sij 3.78**), 'Hurakan' x 'Lolita' (5x4) (Sij 3.86**) and 'Tala' x 'Terminator' (1x7) (Sij 3.25**), which could be used in a breeding program to exploit the dominance effects by means of cyclic hybridization. The earliest three crosses, with 44 days to feminine flowering, were: 'Lolita' x 'Tala' (4x1), 'WA9041' x 'Tala' (6x1), 'Dolarzini' x 'Tala' (8x1) and the latest was 'Terminator' x 'Hurakan' (7X5)(54 days).Keywords: Cucurbita pepo L., diallel analysis, fruit yield.
INTRODUCCIÓN
La forma de aprovechamiento más importante en México de los frutos de calabaza (Cucurbita pepo L.) es como verdura, tanto para el consumo nacional como para su exportación. En 2009 se cultivaron 1,525,378 ha en el mundo, y en México, 26,600 ha. México está entre los siete principales productores y aun cuando experimentalmente se han obtenido y reportado rendimientos de 73 t·ha−1, el rendimiento nacional es inferior a los obtenidos en España (42.9 t·ha−1), Francia (40 t·ha−1) y países bajos (55.0 t·ha1) (Anónimo, 2008). Aunado a lo anterior, no existen programas en México enfocados al mejoramiento genético de calabacita para verdura, por lo que estudiar los efectos heteróticos partiendo de híbridos comerciales es una opción para iniciar un programa de mejoramiento genético para la obtención de híbridos y variedades de alto rendimiento.
Gardner y Eberhart (1966) propusieron varios métodos de análisis, de los cuales el más utilizado es el análisis tipo II. Esta metodología puede aplicarse desde grupos parentales totalmente homocigotos (F = 1) hasta aquellos sin ningún grado de endogamia (F = 0). Típicamente requiere de la evaluación de los padres y los cruzamientos F1 directos. Este método considera sólo modelos fijos y, por lo tanto, no tiene sentido aplicarlo para la estimación de componentes de varianza genética (modelos aleatorios).
En su trabajo, Gardner y Eberhart (1966) señalan que esta metodología permite un análisis más preciso y detallado de los efectos heteróticos que se manifiestan en los cruzamientos. En general, este método se ha usado con más frecuencia en el análisis de cruzamientos dialélicos, en el que los padres son variedades de polinización abierta (Pérez et al., 1995; Machado y Miranda, 2003).
La estimación de los parámetros genéticos se obtiene por medio del análisis de los diseños dialélicos propuestos por Griffing (1956a, 1956b) y Gardner y Heberhart (1966). Estos últimos autores propusieron un análisis alternativo de los datos de cruzas dialélicas obtenidas de poblaciones o progenitores heterogéneos (variedades). El análisis del Método I de Gardner y Heberhart (GEAN I) requiere la evaluación de n progenitores, n(n−1)/2 cruzas F1 y la progenie de los progenitores y las cruzas, pero proporciona información de la acción génica aditiva, de dominancia, heterosis y depresión endogámica. El Método II (GEAN II) es útil en la evaluación de n poblaciones (variedades) y sus n(n−1)/2 cruzas F1. La heterosis se particiona en heterosis promedio, varietal y específica. Sin embargo, no se pueden estimar los efectos aditivos y de dominancia porque están confundidos con la fuente de variación "variedad". El Método III (GEAN III) estima los efectos de variedad y Aptitud Combinatoria General (ACG) de un análisis que contiene las siguientes fuentes de variación: progenitores, progenitores vs cruzas F1, y cruzas F1. Los efectos de ACG se estiman de manera similar al Método 4 Modelo 1 de Griffing (Gardner y Heberhart, 1966; Murray et al., 2003).
Puesto que los híbridos comerciales pueden ser usados exitosa y ventajosamente en programas de mejoramiento genético, el objetivo del presente trabajo fue identificar híbridos comerciales con valor genético en un programa de mejoramiento para producir híbridos y variedades de alto rendimiento.
]]> MATERIALES Y MÉTODOS
Progenitores
Se emplearon ocho híbridos de calabacita tipo 'Grey Zucchini' de diferentes casas comerciales (Cuadro 1). Con ellos se obtuvieron las 56 cruzas (28 directas y 28 recíprocas) posibles en la primavera de 2006, garantizando en cada cruza al menos la polinización de 10 plantas del progenitor femenino.
Evaluación experimental
Las cruzas y los progenitores se evaluaron en dos ciclos en el año 2007 y 2008 en el Campo Agrícola Experimental de la Universidad Autónoma Chapingo. En el año 2007 la siembra se realizó el día 15 de abril y en 2008 el día 20 de abril de forma directa, depositando dos semillas por golpe para posteriormente ralear a una planta, bajo condiciones de riego.
La unidad experimental estuvo constituida por parcelas de dos surcos de 5 m de largo separados a 0.8 m, con una separación entre plantas de 0.2 m (62,500 plantas·ha−1). Se usó un diseño experimental bloques al azar con tres repeticiones.
La fertilización total fue de 120-80-00, se aplicó 6080-00 al momento de la siembra, y el resto de la fertilización nitrogenada en el aporque a los 35 días de la siembra. Las malezas se controlaron manualmente.
Se realizaron 17 cortes o cosechas de fruto, a partir de los cuales registró: peso de fruto por corte (PFR, g); número de frutos por corte (NFR); largo y ancho de fruto (LFR, AFR, cm), de una muestra aleatoria de cinco frutos; rendimiento por planta por corte (g·planta−1), como el cociente del peso de frutos por corte, entre el número de plantas por unidad experimental, y rendimiento por hectárea (t·ha−1). Adicionalmente se registró los días a floración masculina y femenina (DFM y DFF), cuando existía 50 % de plantas con flores abiertas.
Heterosis
Se utilizó el modelo de análisis II, propuesto por Gardner y Eberhart (1966). En éste, la expresión del valor fenotípico promedio de cualquier padre o híbrido F1, está dada por la ecuación:
]]>
Donde:
Yij = valor promedio esperado para una variedad (i = j) o para un cruzamiento (i ≠ j);
µv = = es la media de las n variedades parentales;
Vi = efecto de la i-ésima variedad;
Vj = efecto de la j-ésima variedad;
θ = : Si i = j, entonces θ = 0, y si i ≠ j, entonces θ = 1;
hij= efecto de heterosis que resulta cuando la variedad i se cruza con la variedad j. Este efecto ocurre solamente en los cruzamientos F1.
El efecto de la heterosis a su vez puede dividirse así:

 = heterosis promedio de todos los cruzamientos;
= heterosis promedio de todos los cruzamientos;
hi = heterosis varietal producida por la i-ésima variedad;
hj = heterosis varietal producida por la j-ésima variedad;
Sij = heterosis específica para el cruzamiento entre las variedades i y j; es una desviación de la media esperada basada en los efectos ( + hj)
De acuerdo con Gardner (1967), el efecto de la variedad (Vi), es la diferencia entre la media de un padre per se y la media de todos los padres. La heterosis promedio (), del grupo de progenitores usados en el estudio es la diferencia entre la media de todos los cruzamientos F1 y la media de todos los progenitores. La heterosis varietal (hi), es la contribución de heterosis por la variedad i en sus cruzamientos F1, medida como una desviación del promedio de heterosis. La heterosis específica (Sij) entre las variedades i y j, y mide la desviación entre el comportamiento observado de un cruzamiento específico y su comportamiento esperado, basado en el efecto varietal (Vj) la heterosis promedio () y la heterosis varietal (hi).
La heterosis se calculó con base en modelo de Gardner y Eberhart (1966), ya descrito, en el que también se consideran los siguientes parámetros:
a) Heterosis media ( )
)

Donde:
]]>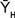 = media de todas las cruzas realizadas;
= media de todas las cruzas realizadas;  = media de todos los progenitores.
= media de todos los progenitores.
b) Heterosis varietal (hi):

Donde:
n = número de progenitores;
 = media de las cruzas en que interviene el progenitor i;
= media de las cruzas en que interviene el progenitor i;
 = media de todas las cruzas realizadas;
= media de todas las cruzas realizadas;
Yi = media del progenitor i;
 = media de todos los progenitores.
= media de todos los progenitores.
c) Heterosis respecto al mejor progenitor (Hmp),
Hmp = Yij − Ymp
Donde:
Yij = media de la cruza i x j
Ymp = media del mejor progenitor de la cruza i x j
d) Heterosis específica de cruzas entre los progenitores i j (Sij)
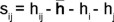
Donde:
hij = heterosis con respecto al progenitor medio de cada cruza i j
]]> = heterosis promedio de todos los cruzamientos;
= heterosis promedio de todos los cruzamientos; hi = heterosis varietal producida por la i-ésima variedad;
hj = heterosis varietal producida por la j-ésima variedad;
Análisis estadístico
El análisis estadístico se realizó con el programa SAS versión 9.0 para microcomputadora, estimando los efectos varietales, heterosis específica, heterosis media y heterosis varietal con el procedimiento MIXED de SAS considerando como factor de efectos fijos a los materiales genéticos. La heterosis con respecto al mejor progenitor Hmp% se calculó con la fórmula:

Donde:
Hmp% = heterosis en porcentaje en relación al mejor progenitor
Yij= media de la cruza i x j
Ymp = media del mejor progenitor de la cruza i x j
]]>RESULTADOS Y DISCUSIÓN
Los cuadrados medios combinados por ambientes y los niveles de significancia para los caracteres rendimiento por planta (RPP, g), frutos por planta (FPP), largo de fruto (LFR, cm), ancho de fruto (AFR, cm), días a floración masculina (DFM), días a floración femenina (DFF) y rendimiento de calabacita para verdura por hectárea (REN, t·ha−1) con el modelo II propuesto por Gardner y Eberhart (1966), se presentan en el Cuadro 2. Al examinar las fuentes de variación más importantes de este análisis, la significancia de heterosis x ambiente indica que la heterosis cambió a través de los ambientes. Dado que no se encontró significancia estadística en heterosis promedio por ambiente, se infiere que se mantiene estable a través de los ambientes. En la comparación de cruzas vs híbridos, los progenitores obtuvieron un rendimiento de verdura de 33.7 t·ha−1, y las cruzas, de 34.8 t·ha−1 (heterosis promedio por ambientes).
En heterosis varietal por ambiente se encontró significancia estadística, lo cual indica que la heterosis varietal de los híbridos no fue estable a través de ambientes, pero tampoco fue muy alta debido probablemente a que varios de los híbridos usados como progenitores están emparentados (cuatro de los ocho híbridos empleados como progenitores son de la misma compañía). Respecto a la interacción heterosis específica x ambiente que fue significativa revela que la heterosis de las cruzas fue diferente para cada localidad.
Heterosis media ( )
)
La heterosis media, dado que se calcula como la diferencia entre la media de todos los híbridos F1 (cruzas dialélicas posibles entre los progenitores estudiados) y la media de todos los progenitores como tal, es un primer indicador de la existencia o no de heterosis en el conjunto de los híbridos F1 formados (cruzamientos dialélicos), con respecto a la población de los progenitores comerciales utilizados. La existencia de heterosis media puede interpretarse como una consecuencia de la divergencia genética entre los híbridos comerciales progenitores, ya que numerosos estudios experimentales de varios autores han mostrado que a mayor divergencia genética entre los progenitores hay mayor heterosis en las cruzas.
Se encontró heterosis media positiva (Cuadro 10) en todos los caracteres estudiados (REN = 1.10 t·ha−1; RPP = 13.10 g·planta−1; AFR = 0.10 cm; LFR = 0.50 cm; DFF = 3 días y DFM = 3 días). La existencia de heterosis positiva en todos los caracteres evaluados sugiere que en la población teórica obtenida del dialélico entre ocho híbridos comerciales de calabacita son más importantes los efectos de aptitud combinatoria específica que los de los efectos de aptitud combinatoria general. Lo anterior coincide con el análisis de varianza del análisis dialélico con la metodología de Montesinos et al. (2007) en el que la ACE es más importante que la ACG en los principales componentes del rendimiento de calabacita tipo Grey Zucchini (Análisis no mostrado). La heterosis media en días a floración masculina y femenina mostró efectos positivos, situación no deseada, puesto que refleja ciclos más tardíos de las cruzas.
Heterosis varietal (hi)
El híbrido comercial 'Terminator' manifestó efectos positivos favorables de heterosis varietal en todos los caracteres evaluados (Cuadro 10), incluidos los principales componentes de rendimiento (RPH = 3.62 t·ha−1, RPP = 63.37 g·planta−1, FPP = 0.73 frutos, LFR = 0.33 cm, AFR = 0.20 cm, DFF = 1.66 días y DFM = 1.67 días). Sin embargo, los valores de heterosis varietal positivos en días a floración femenina y masculina son indeseables agronómicamente, ya que sus cruzas tienden a ser más tardías. El progenitor 5 también mostró una heterosis varietal positiva, en días a floración (DFF= 0.50 y DFM = 1.67), efecto no deseado, pues implica que produce híbridos tardíos (Cuadro 10). Los demás progenitores presentan valores negativos de heterosis varietal para días a floración femenina, situación que los hace deseables desde el punto de vista agronómico porque en sus cruzas se mantiene la precocidad de los progenitores.
El híbrido 'Tala' también presentó heterosis varietal positiva en rendimiento (REN = 4.26 t·ha−1, RPP = 73.51 g·planta−1 y FPP = 2.11 frutos), y el híbrido 'Lolita', heterosis varietal positiva en rendimiento (REN = 0.53 t·ha−1, RPP = 2.58 g·planta−1 y LFR = 0.56 cm) (Cuadro 10). Los resultados de la heterosis varietal positiva tanto en los progenitores 'Terminator' (7), 'Tala' (1) y 'Lolita' (4) explican la alta productividad mostrada por sus cruzas.
]]> El híbrido 'Dolarzini' presentó la menor heterosis varietal en rendimiento (REN = −3.15 t·ha−1, RPP = −66.0 g·planta−1, FPP = −1.63 frutos, LFR = −0.51 cm y AFR = −0.16 cm, (Cuadro 10), seguido de 'Grey Zucchini' de seminis con heterosis varietal en rendimiento (REN = −2.48 t·ha−1, RPP = −34.65 g·planta−1, FPP: −0.13 frutos, LFR = −0.32 cm y AFR = −0.05 cm) y 'Grey Zucchini M+M' (REN = −1.95 t·ha−1, RPP = −26.65 g·planta−1, FPP = −0.07 frutos, LFR = −0.15 cm y AFR = −0.12 cm).Con base en lo anterior, para un programa de selección recíproca recurrente, en donde se explota tanto los efectos aditivos como los no aditivos, los mejores progenitores para rendimiento por planta y por hectárea serían 'Terminator' y 'Lolita' que mostraron mayor heterosis varietal.
Heterosis específica (Sij)
La heterosis específica permitió identificar las cruzas de alto rendimiento con mejores resultados agronómicos que los progenitores comerciales. Tres cruzamientos presentaron efectos de heterosis específica (Sij) significativos en rendimiento por hectárea (Cuadro 9): 'Grey Zucchini M+M' x 'Terminator' (3 x 7) (Sij 3.78**), 'Hurakán' x 'Lolita' (5 x 4) (Sij 3.86**) y 'Tala' x 'Terminator' (1 x 7) (Sij 3.25**). El valor de heterosis específica positivo y significativo sugiere que su promedio fue superior al promedio esperado con base en la gi de sus progenitores y la media general.
El mayor rendimiento por planta y por hectárea (Cuadro 3 y Cuadro 9), así como el mayor número de frutos por planta y largo y ancho de fruto (Cuadro 5 y Cuadro 6), lo presentaron las cruzas con la mayor heterosis específica y además, con respecto al mejor progenitor (Cuadro 11).
En rendimiento por hectárea el mejor cruzamiento fue 'Grey Zucchini M+M' x 'Terminator' (3 x 7) con 50.0 t·ha−1 (Cuadro 9), 799.9 g·planta−1 (Cuadro 3) y 17.5 frutos por planta de calabacita (Cuadro 4) en 17 cortes acumulados, que superó en 15.3 % el rendimiento por hectárea al mejor de sus progenitores. Esto se explica en parte porque esta cruza es la que presentó la mayor heterosis tanto específica (Sij) como con respecto al mejor progenitor (Hmp), situación que sólo permite usarla como híbrido F1. La cruza 'Lolita' x 'Tala' (4 x 1) promedió un rendimiento por hectárea de 50.14 t·ha−1 y superó 18.8 % de heterosis con respecto al mejor de sus progenitores. La cruza 'Lolita' x 'Terminator' rindió igual que el mejor de sus progenitores (44.09 t·ha−1).
Dos de las mejores cruzas, 'Lolita' x Tala (4 x 1) (50.14 t·ha−1) y 'Lolita' x 'Terminator' (4 x 7) (44.09 t·ha−1), se obtuvieron con progenitores que pertenecen a la misma compañía semillera, lo que indica que presentan alta divergencia genética. Mientras que la cruza 'Tala' x 'Grey Zucchini' de Seminis (1 x 2) tuvo la heterosis específica más baja, lo que sugiere que estos materiales están relacionados genéticamente.
Heterosis respecto al mejor progenitor (Hmp%)
La heterosis de mayor significado agronómico es la que se calcula con respecto al mejor progenitor. La cruza con mayor heterosis con respecto al mejor progenitor en rendimiento por hectárea, por planta y número de frutos por planta fue 'Grey Zucchini' M+M' x 'Terminator' (3 x 7) (Cuadro 11), la cual superó en 15.3 % el rendimiento por hectárea (44.2 t·ha−1) y por planta (707.58 g·planta−1), y en 1.5 % en frutos por planta (13.6) a su mejor progenitor. Le sigue la cruza 'Lolita' x 'Hurakán' (4 x 5), que superó en 25.6 % el rendimiento por planta (675.3 g·planta−1) y por hectárea (42.2 t·ha−1) con respecto a su mejor progenitor.
La mejor cruza para largo de fruto fue 'Tala' x 'Dolarzini' (1 x 6) (Cuadro 5), con una heterosis con respecto al mejor progenitor de 11.5 % seguida de la cruza 'WA9041' x 'Lolita' (6x4) (8.1 % de Hmp ) (Cuadro 11). No se encontraron efectos significativos (p ≤ 0.05) de Hmp en ancho de fruto.
]]> En floración femenina y masculina, se encontró heterosis positiva respecto al mejor progenitor en la cruza 'Hurakán x 'Terminator' (5 x 7) (DFF, 7.7% de Hmp ) y (DFM, 1.8 % de Hmp ), tanto para días a floración femenina, como para días a floración masculina. Esto significa que las cruzas (de efectos positivos) son más tardías que el progenitor más tardío, lo cual agronómicamente no es deseable, por lo que son preferibles las cruzas con heterosis negativa, las más precoces. En este sentido, la mayoría de las cruzas donde intervinieron como progenitores los híbridos 'Tala', 'Lolita' y 'WA9041' presentaron valores negativos en su mayoría en heterosis con respecto al mejor progenitor, lo cual indica que, en general, las cruzas donde intervienen estos progenitores tienen igual calidad agronómica y precocidad que sus progenitores.Las tres cruzas más precoces, con 44 días a floración femenina, fueron 'Lolita' x 'Tala' (4 x 1), 'WA9041' x 'Tala' (6 x 1), 'Dolarzini' x 'Tala' (8 x 1) (Cuadro 7), resultado que es consistente con las respectivas heterosis negativas (4 x 1 = −2.3, 6 x 1 = −6.9, 8 x 1 = −2.3 días) (Cuadro 11) que reflejan precocidad en dichas cruzas. Mientras que las más tardías fueron las cruzas 'Terminator' x 'Hurakán' (7 x 5), 'Terminator' x 'Grey Zucchini de seminis' (7 x 2) y 'Hurakán' x 'Grey Zucchini M+M' (5 x 3) (Cuadro 8), con valores positivos de heterosis con respecto al mejor progenitor (7 x 5 = 11.5 %; 7 x 2 = 5.8 % y 5 x 3 = 5.8 %) (Cuadro 11).
Finalmente, en rendimiento por hectárea, las cruzas mejores en heterosis con respecto al mejor progenitor fueron 'Grey Zucchini M+M' x 'Terminator' (3 x 7) (17.9 % Hmp), 'Lolita' x 'Hurakán' (4 x 5) (25.6 % Hmp ) y 'Terminator' x 'Tala' (7 x 5) (10.4 % Hmp ) (Cuadro 11). Los resultados del presente estudio en calabacita tipo Grey Zucchini coinciden con los de Ruiz et al. (2004) en calabaza (Cucurbita moschata Duch), Martínez (2004) y Magaña (2006) en jitomate (Lyco-persicon esculentum), quienes encontraron cruzamientos dialélicos entre progenitores comerciales que superaron a los híbridos comerciales progenitores.
CONCLUSIONES
Se encontró heterosis significativa en las cruzas entre híbridos comerciales de calabacita tipo Grey Zucchini con el modelo II de Gardner y Eberhart (1966) para los principales componentes del rendimiento (largo de fruto, rendimiento por planta y por hectárea).
Los híbridos con los mayores efectos varietales para rendimiento por planta y por hectárea fueron 'Terminator', 'Lolita' y WA9041, y las cruzas entre híbridos que presentaron mayor heterosis específica fueron 'Grey Zucchini M+M' x 'Terminator' (3 x 7), 'Hurakán' x 'Lolita' (5 x 4) y 'Tala' x 'Terminator' (1 x 7). Estas cruzas pueden ser recomendadas por su uso como híbridos de cruza doble y en un programa de mejoramiento genético para aprovechar los efectos de dominancia mediante hibridación cíclica.
Las cruzas 'WA9041' x 'Lolita' (6 x 4) y 'WA9041' x 'Tala' (6 x 1) mostraron efectos significativos en heterosis específica en largo de fruto.
Las tres cruzas más precoces fueron 'Lolita' x 'Tala' (4 x 1), 'WA9041' x 'Tala' (6 x 1), 'Dolarzini' x 'Tala' (8 x 1), y la más tardía fue 'Terminator' x 'Hurakán' (7 x 5).
]]> LITERATURA CITADA
ANÓNIMO. 2008. Superficie, producción y rendimiento de calabazas en el mundo. Food and Agriculture Organization. http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor [ Links ]
GARDNER, C. O.; EBERHART, S. A. 1966. Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22(3): 439-452. http://www.jstor.org/stable/pdfplus/2528181.pdf [ Links ]
GARDNER, C. O. 1967. Simplified methods for estimating constants and computing sums of squares for a diallel cross analysis. Fitotecnia Latinoamericana. 4(2): 1-12. http://hdl.handle.net/2027/coo.31924066717384 [ Links ]
GRIFFING, B. 1956a. A generalized treatment of the use of diallel crosses in quantitative inheritance. Heredity 10: 31-50. doi:10.1038/hdy.1956.2 [ Links ]
GRIFFING, B. 1956b. Concept of general and specific combining ability in relation to diallel crossing system. Austr. J. Biol. Sci. 9:463-493. http://www.publish.csiro.au/?act=view_file&file_id=BI9560463.pdf [ Links ]
MAGAÑA L., N. 2006. Comportamiento productivo de poblaciones F2 y heterosis intervarietal en jitomate (Lycopersicon esculentum Mill.) cultivado a un racimo. Tesis de Maestría en Ciencias. Instituto de Horticultura. Fitotecnia. Chapingo. México. 83 p. [ Links ]
MACHADO, R.; MIRANDA, J. B. 2003. Heterosis expression in crosses between maize populations. year yield. Scientia Agricola 60(3):519-524. http://www.scielo.br/pdf/sa/v60n3/16407.pdf [ Links ]
MARTÍNEZ S., J. 2004. Heterosis intervarietal y uso de generaciones F2 de híbridos de jitomate (Lycopersicon esculen-tum Mill.). Tesis de Doctorado en Ciencias en Horticultura. Fitotecnia. Chapingo. México. 88 p. [ Links ]
MONTESINOS L., O. A.; MASTACHE L., A. A.; LUNA E., I.; HIDALGO C., J. V. 2007. Mejor predictor lineal e insesgado combinado para aptitud combinatoria general y análisis combinado de los diseños uno y tres de Griffing. Téc. Pecu. Méx. 45(2):131-146. http://www.tecnicapecuaria.org.mx/trabajos/200705295738.pdf [ Links ]
MURRAY, L. W.; RAY I. M.; DONG, H.; SEGOVIA-LERMA, A. 2003. Clarification and reevaluation of population-based diallel analyses: Gardner and Eberhart analyses II and III revisited. Crop Science. 43(2):1930-1937. doi:10.2135/ cropsci2003.1930 [ Links ]
PÉREZ, J. C.; CEBALLOS, H.; PANDEY, S.; DÍAS, C. 1995. Analysis of diallel crosses among Colombian landraces and improved populations of maize. Crop Science 35(1):572-578. doi: 10.2135/cropsci1995.0011183X003500020048x [ Links ]
RUIZ E.; SIGARROA A.; CRUZ J., A. 2004. Análisis dialélico del rendimiento y sus principales componentes en variedades de calabaza (Cucurbita moschata Duch) I Tabla dialélica de Griffing. Biología 18: 65-73. http://revistas.mes.edu.cu/greenstone/collect/repo/archives/D0864349/0181065.dir/08643490181065.pdf [ Links ] ]]>