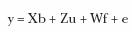
Efectos de interacción genotipo por ambiente para peso corporal a los 130 días en camarón blanco del Pacífico [Penaeus (Litopenaeus) vannamei]
Genotype by environment interaction effects for body weight at 130 days of age in the Pacific white shrimp [Penaeus (Litopenaeus) vannamei]
Gabriel Ricardo Campos–Montes* Hugo H. Montaldo* Alfonso Martínez–Ortega** Héctor Castillo–Juárez***
* Departamento de Genética y Bioestadística, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, 04510, México, D. F.
** Maricultura del Pacífico, S. A. de C. V., Pesqueira 502, Local 5, Colonia Centro,82040, Mazatlán, Sinaloa, México.
]]> *** Departamento de Producción Agrícola y Animal, Universidad Autónoma Metropolitana–Unidad Xochimilco, Calzada del Hueso 1100, Colonia Villa Quietud, 04960, México, D. F.
Autor de contacto:
Héctor Castillo–Juárez,
Tel.: +52 55 5617 4126; fax: +52 55 5483 7230,
correo electrónico: hcjuarez@correo.xoc.uam.mx
Recibido el 28 de mayo de 2009
Aceptado el 5 de agosto de 2009
]]> Abstract
Body weight of Pacific white shrimp [Penaeus (Litopenaeus) vannamei] at 130 days of age was analyzed in three environments corresponding to different management systems: semi–intensive (10 shrimp/m2) in Pozos, Sinaloa (POZOS10), intensive (30 shrimp/ m2) in Pozos, Sinaloa (POZOS30), and super–intensive (85 shrimp/m2) in Bahia de Kino, Sonora (KINO85). Data were obtained from 18 087 sibs from 113 sires and 143 dams. The aim of the study was to evaluate the presence of genotype by environment interaction effects (IGA) and the effect of the (co) variance between full–sibs family common effects on the genetic parameter estimates. Estimates of h2 with a model including independent full–sibs family common effects were between 0.26 and 0.39 across environments, while the estimates for a model with correlated full–sibs family common effects were estimated between 0.14 and 0.23. No differences were found for h2 values between environments. The genetic correlations between environments were not lower from unity with any model; therefore, it is concluded that no evidence of genotype–environment interaction exists for body weight at 130 days in Pacific white shrimp, under the environments used in this study. The inclusion of the (co)variance between full–sibs family common effects of different environments affected the parameter estimates. These results also indicate that ranking of the breeding animals will be similar in all the studied production environments.
Key words: Shrimp Breeding, Pacific White Shrimp, Genotype by Environment Interaction.
Resumen
Se analizó información de peso corporal de camarón blanco del Pacífico [Penaeus (Litopenaeus) vannamei] a los 130 días, en tres sistemas de manejo: semiintensivo (10 camarones/m2); intensivo (30 camarones/m2), ambos en Pozos, Sinaloa, y super–intensivo (85 camarones/m2), en Bahía de Kino, Sonora. Los registros corresponden a 18 087 individuos, hijos de 113 sementales y 143 hembras, con la finalidad de evaluar la existencia de interacciones genotipo por ambiente (IGA) y el efecto de la covarianza de los efectos comunes de familia de hermanos en la estimación de parámetros genéticos. Las estimaciones de h2 con un modelo que consideró los efectos comunes de familia de hermanos como independientes, fueron de 0.26 a 0.39 y de 0.14 a 0.23, para un modelo que consideró dichos efectos como correlacionados. No existió diferencia significativa entre los valores de h2 de los ambientes, y las correlaciones genéticas entre ambientes no fueron menores a uno con ninguno de ambos modelos, se concluye que no hay evidencia de IGA en el peso corporal a los 130 días de P. vannamei en los ambientes estudiados en este trabajo. La inclusión de la covarianza entre efectos de familias de hermanos en el análisis afectó los valores de los parámetros. Se concluye que el ordenamiento de los reproductores será similar en los ambientes estudiados.
Palabras clave: Mejoramiento genético, Camarón blanco del Pacífico, Interacción Genotipo–Ambiente.
Introducción
En la producción de camarón blanco del Pacífico [Penaeus (Litopenaeus) vannamei], las características de crecimiento y supervivencia determinan de manera importante la rentabilidad. Por ello, la correcta evaluación de los posibles reproductores, así como el diseño de programas de selección eficaces, requiere la estimación de componentes de varianza del peso a edades comerciales.
]]> En general, resulta preferible que las evaluaciones genéticas se realicen en condiciones ambientales similares a las que prevalezcan en el lugar en que se desempeñarán los descendientes de los animales seleccionados, debido a que las condiciones de manejo pueden hacer variar la expresión genética aditiva [por ejemplo, interacción genotipo–ambiente (IGA)], haciendo que las respuestas a la selección dependan del ambiente.1,2 Sin embargo, lo anterior rara vez es posible en la camarinocultura debido a las diferencias de manejo existentes entre las granjas núcleo y las granjas de engorda.Entre los principales factores que determinan las diferencias de manejo en las granjas camaroneras está la densidad de siembra, que es un factor determinante en el crecimiento del camarón.3 Por ello, es importante estudiar su efecto en las evaluaciones genéticas, como ocurre en otras especies acuícolas.4
Es posible clasificar a las granjas de engorda de camarón en cuatro sistemas, cuyas principales diferencias se basan en densidad de siembra, cantidad de alimento recibido, apoyo de oxigenación y proporción de recambio de agua. Las densidades de siembra en los sistemas extensivos varían entre dos y cinco cama–rones/m2, en semiintensivos entre ocho y 25 camaro–nes/m2, en intensivos entre 25 y 50 camarones/m2 y en los superintensivos las densidades de siembra son superiores a 50 camarones/m2.2,5,6 Diversos autores han estudiado en el camarón las posibles interacciones genotipo por ambiente, identifican a la densidad de siembra como uno de los factores más importantes en la definición de los ambientes.
Pérez–Rostro e Ibarra7 no observaron diferencias en el orden secuencial entre los valores genéticos para peso corporal a los 200 días de las familias sembradas en dos estanques con densidades de siembra de 2.5 y 4.3 camarones/m2. En los estudios de Gitterle et al.8 tampoco encontraron evidencia de IGA en el peso corporal a los 160 días de edad en P. vannamei, en siete granjas del Atlántico colombiano con salinidades que variaron de 0 a 35 ppt y densidades de 18 a 22 camarones/m2.
Sin embargo, otros estudios muestran posible existencia de IGA en características de crecimiento en P. vannamei. Castillo–Juárez et al.9 encontraron evidencia de IGA, expresada como diferencias de magnitud de la heredabilidad entre diferentes densidades de siembra (9 y 14 camarones/m2) para el peso corporal en P. vannamei a los 130 días, aunque las correlaciones genéticas entre estos ambientes fueron muy altas y positivas. Asimismo, estos autores describieron las correlaciones de los efectos comunes de familia, cercanas a la unidad entre ambientes; la inclusión de este efecto en los modelos causó cambios en la estimación de las heredabilidades y de sus errores estándar. Por su parte, Ibarra y Famula,6 sin considerar los efectos comunes de familia de hermanos en los modelos de análisis, calcularon la correlación genética entre el peso adulto en P. vannamei como 0.54 ± 0.12 en densidades iniciales de 5.9 camarones/m2 y de 400 camarones/m2.
El objetivo de este estudio fue evaluar la existencia de interacciones genotipo por ambiente para peso corporal a los 130 días de P. vannamei en tres sistemas de cultivo: semiintensivo (10 camarones/m2), intensivo (30 camarones/m2) localizados en Pozos, Sinaloa, y superintensivo (85 camarones/m2), localizado en Bahía de Kino, Sonora. Además, se estudió el efecto de inclusión de covarianza entre los efectos de ambiente común de familia entre ambientes, sobre la estimación de los parámetros genéticos.
Material y métodos
Localización
Se utilizó la información del peso a los 130 días de edad de organismos de P. vannamei del ciclo de producción de 2008, cosechados en septiembre de ese año, procedentes de tres estanques de engorda de una empresa* que se dedica a la producción de larva de esta especie de camarón en México. Los ambientes semi–intensivo e intensivo se localizan en el ejido de Los Pozos, municipio de Rosario, Sinaloa, México, y el ambiente superintensivo en el municipio de Bahía de Kino, Sonora, México.
]]>Formación y manejo de familias
Las familias empleadas provienen de una población domesticada de P. vannamei que fue seleccionada desde 2004 para aumentar el peso corporal a los 130 días de edad. Los progenitores fueron marcados individualmente con anillos colocados en un pedúnculo ocular, su proceso de maduración se realizó en tanques diferentes para cada sexo.
A partir de inseminación artificial, se generaron 300 familias de hermanos y medios hermanos paternos usando una proporción de dos hembras por cada macho. Se evitaron apareamientos que produjeran familias con 6.25% o más de consanguinidad.
Las hembras inseminadas fueron colocadas en tanques de 500 L, donde desovan. Luego de ocurrida la eclosión, los nauplios de cada familia se sembraron en dos tanques con 200 L a una densidad aproximada de 100 camarones/L. La alimentación de los organismos consistió en alimento comprimido comercial con contenido de proteína que varió entre 40% y 50% y de 8% a 10% de lípidos, microalgas del género Chaetoceros, espirulina comercial y Artemia. La dieta se ajustó de acuerdo con la etapa de crecimiento de los animales.
A los 28 días de la eclosión se seleccionaron las 150 familias con los valores genéticos predichos más altos para peso corporal a dicha edad y se trasladaron 2 100 animales de cada familia a jaulas de 1.5 x 0.5 x 0.85 m, que se colocaron en tanques de 24 x 3.5 m con una columna de agua de 0.35 m.
Las jaulas fueron colocadas en estanques en los que se mantuvo oxigenación constante, con un sistema de biofiltros que realiza recambio de agua del estanque de 300% cada 24 horas con temperatura constante de 30°C. Los animales recibieron alimento comercial con 35% a 40% de proteína, ajustándose el tamaño de la partícula y el porcentaje de proteína a la edad de los organismos.
Un mes después, los individuos de cada familia seleccionada a los 28 días fueron marcados con tres elastómeros de seis posibles colores** que fueron colocados en el último segmento abdominal, uno en cada lado del camarón y otro de manera dorsal. La combinación de colores y posiciones representó el código para la identificación de cada familia. Una semana después del marcaje de los individuos, éstos fueron trasladados a dos estanques de engorda, ubicados en Los Pozos, Ahí se sembraron 50 animales de cada familia en dos estanques de tierra de 0.2 ha a densidades de 10 (POZOS10) y 30 (POZOS30) camarones/ m2, representativas de los sistemas comerciales comunes en México.
Los estanques tuvieron una columna de agua de 1.4 m con temperatura de entre 30 y 34°C con salinidad en rango de 30 a 35 ppt y recambio diario de agua de entre 5% y 20%, según la etapa de crecimiento. Se proporcionó alimento comercial con 35% a 40% de proteína, la cantidad diaria correspondió a 3% de la biomasa de los estanques. Un tercer grupo de 50 individuos por familia fue sembrado en Bahía de Kino (KINO85) en un estanque de concreto de 4 m de ancho, 16 m de largo y 2 m de profundidad a densidad de 85 camarones/m2 con temperatura promedio de 30°C, salinidad de 35 ppt con aereación artificial constante y recambio diario de agua menor a 5%. La alimentación en este estanque consistió en alimento comercial con 35% a 40% de proteína, la cantidad diaria correspondió a 6% de la biomasa del estanque.
]]> Obtención y edición de datos de peso corporal a los 130 días
Todos los individuos de cada tanque fueron cosechados entre las semanas 11 y 12 posteriores a la siembra. A los animales de cada familia se les determinó el sexo y la ausencia o presencia de defectos físicos. Luego, para obtener el peso a la cosecha, se les retiró el agua residual con una toalla de tela y se colocaron de forma individual dentro de un vaso en una báscula tarada a cero gramos. Se descartaron del análisis los individuos con deformidades, enanismo, sexo indefinido y marcas de familia incompletas o con errores evidentes de lectura, que representaron 9.5% de los registros iniciales. Las familias provienen de 113 machos y 143 hembras, aquéllas fueron sembradas en los tres ambientes estudiados. Los números de observaciones por ambiente se muestran en el Cuadro 1.
Estimación de parámetros genéticos y ambientales
La estimación de los componentes de varianza se realizó a partir de la utilización de modelos animales bivariados, donde se consideró la medición en cada ambiente como variable diferente (POZOS10, POZOS30 y KINO85). Los efectos fijos que se incluyeron en los modelos fueron sexo y edad a la cosecha como covariable lineal. En el primer modelo (Modelo RG) se consideró a los efectos comunes de familia de hermanos como independientes, en tanto que el segundo modelo (Modelo RF) los consideró como correlacionados. La matriz de relaciones aditivas (A) incluyó a todos los progenitores nacidos desde 2002, además de los camarones con registro propio de peso a los 130 días. Se realizaron análisis trivariados; sin embargo, debido a problemas de convergencia, se decidió emplear análisis bivariados para cada combinación de pareja de ambientes. La representación matricial del modelo RF fue:
donde:
y = vector de observaciones del peso a los 130 días en cada pareja de ambientes analizados (POZOS10 con POZOS30, POZOS10 con KINO85 o POZOS30 con KINO85)
b = vector de efectos fijos,
u = vector de efectos aleatorios genéticos aditivos directos,
]]> f = vector de efectos aleatorios comunes de familia de hermanos (por ejemplo, ambientales y genéticos maternos y genéticos no aditivos),
f = vector de efectos aleatorios comunes de familia de hermanos (por ejemplo, ambientales y genéticos maternos y genéticos no aditivos), 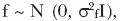 y e = vector de efectos aleatorios residuales,
y e = vector de efectos aleatorios residuales, 
X, Z, W = matrices de incidencia conocidas que relacionan los datos con los efectos fijos, animales genéticos aditivos y comunes de familia de hermanos, respectivamente.
Suponiendo que todas las covarianzas entre u, f y e son nulas, resulta lo siguiente:

donde:
gij = elementos de la matriz G; esto es, las (co) varianzas genéticas aditivas de los pesajes a 130 días de edad de dos ambientes diferentes,
A = matriz de relaciones genéticas aditivas,
fij y eij = elementos de las matrices F y R que representan las (co)varianzas de efectos comunes de familia de hermanos y de los efectos residuales, respectivamente, de los pesos a 130 días mencionados,
I = matrices de identidad de dimensiones adecuadas.
]]> Para la estimación de los componentes de (co) varianza se utilizó el programa AsReml.10 El cálculo de las varianzas fenotípicas se realizó a partir de la suma de los componentes de varianza de los efectos aleatorios de cada variable en cada caso. La heredabilidad (h2) y la proporción de efectos comunes de familia de hermanos (f 2) se estimaron a partir de la proporción de la varianza fenotípica correspondiente de cada componente en cada modelo. Las correlaciones genéticas y de los efectos comunes se estimaron a partir de las covarianzas respectivas, divididas entre los productos de las desviaciones estándar correspondientes.La significancia estadística de los parámetros estimados se determinó con base en los intervalos de confianza construidos a partir de sus errores estándar. La existencia de interacción genotipo por ambiente se determinó a partir de la existencia de diferencias entre las heredabilidades de los pesos en los diferentes ambientes y del valor de las correlaciones genéticas entre estos últimos. Se consideró que existía IGA cuando las heredabilidades fueron diferentes o cuando las correlaciones genéticas fueron estadísticamente menores a uno,1,2 en ambos casos se usó un nivel de significancia de 0.05.
Para el caso de los análisis bivariados correspondientes a los ambientes POZOS10 y POZOS30, los estimados de las correlaciones fueron obtenidos en el límite del espacio parametral, muy cerca de uno, lo que impidió obtener errores estándar válidos para los estimadores de los parámetros genéticos estimados con este modelo. En tales casos, los errores estándar utilizados en las pruebas de hipótesis se obtuvieron de los otros dos análisis bivariados para cada uno de los ambientes correspondientes.
Resultados
Los estadísticos descriptivos se muestran en el Cuadro 1, en tanto que en el Cuadro 2 se encuentran los componentes de varianza estimados con los modelos bivariados. Las heredabilidades (h2) y las proporciones de los efectos comunes de familia de hermanos (f 2) se muestran en el Cuadro 3, en tanto que en el Cuadro 4 se presentan las correlaciones genéticas y las correlaciones de los efectos comunes de familia de hermanos (FH).
La media de peso a los 130 días fue de 12.02 g con desviación estándar de 2.60 g. El promedio del peso corporal a los 130 días disminuyó conforme aumentó la densidad de siembra (Cuadro 1), siendo POZOS10 el sistema con mayor media de peso a los 130 días. Los estimadores de las varianzas genéticas aditivas del Modelo RF fueron menores a las obtenidas con el Modelo RG (Cuadro 2). Las heredabilidades calculadas en los tres sistemas no presentaron diferencia significativa (P > 0.05) cuando se empleó el mismo modelo (Cuadro 3), y en todos los casos las correlaciones genéticas (Cuadro 4) no fueron diferentes de uno (P > 0.05) al igual que en las correlaciones de FH, con excepción de POZOS10–KINO85 donde la correlación fue diferente de uno (P < 0.05).
Discusión
Las medias de peso corporal a los 130 días (Cuadro 1) fueron diferentes entre los ambientes, mayor en POZOS10 (13.54 g) y menor en KINO85 (10.84 g); el efecto de la densidad sobre el peso concuerda con lo observado por otros autores.3,11 Esta reducción del peso al incrementarse la densidad puede explicarse con la competencia por el espacio, el alimento y la modificación de factores químicos como el oxígeno disuelto, pH y nitratos resultantes del incremento de la biomasa. Lo anterior puede inducir a un estado de estrés en los organismos, modificando las respuestas metabólicas, y, por tanto, el aprovechamiento del alimento, además de aumentar la susceptibilidad a enfermedades.3,12,13
]]> Las varianzas aditivas no fueron diferentes entre ambientes (P > 0.05); sin embargo, las varianzas residuales de POZOS10 y KINO85 fueron significativamente superiores a la de POZOS30, pero no diferentes entre sí. Lo anterior derivó en disminución de la varianza fenotípica en este ambiente (Cuadro 2). La heterogeneidad de varianzas genéticas aditivas y residuales en el estudio de características medidas en diferentes ambientes, también se ha descrito en especies no acuícolas;14–16 generalmente está relacionada con factores de manejo, donde los ambientes menos controlados tienen mayor varianza residual; en especies acuícolas, los ambientes más restrictivos provocan alta mortalidad y canibalismo, contribuyendo quizá a disminución de la varianza residual.4,17En concordancia con Gitterle et al.8 y Castillo–Juárez et al.,9 las heredabilidades estimadas aquí no fueron significativamente diferentes entre ambientes (P > 0.05) con ninguno de los dos modelos (Cuadro 3). Sin embargo, los resultados del presente trabajo difieren de los obtenidos por Ibarra y Famula,6 quienes en su modelo empleado omitieron los efectos comunes de FH. Las estimaciones de h2 con el modelo RG fueron superiores a las obtenidas con el modelo RF por lo que la inclusión de la covarianza entre los efectos comunes de FH produjo una importante reducción de los valores de la heredabilidad en todos los casos.
Los valores de efectos comunes de FH del modelo RG no fueron significativamente diferentes de cero en ninguno de los ambientes (Cuadro 3), en tanto que con el modelo RF los valores de f 2 fueron significativamente distintos de cero (P < 0.05), y constantes a través de los ambientes, con cifras que variaron entre 0.07 y 0.10. Estos valores son consistentes con los considerados por Castillo–Juárez et al.,9 y el rango de los estimadores en el presente trabajo es menor que el descrito por Gitterle et al.,8 quienes calcularon valores de f 2 entre 0.00 y 0.17 en siete ambientes comerciales. Hay que destacar que la inclusión de la covarianza entre los efectos comunes de FH provocó aumento en los estimadores de f 2, contrario a lo observado en la valoración de las heredabilidades; por esto es importante considerar su inclusión en los análisis bivariados para la estimación de parámetros poblacionales en características de crecimiento en camarón.
Las correlaciones de las medias fenotípicas familiares sin ajustar entre KINO85 y POZOS10, KINO85 y POZOS30, y POZOS10 y POZOS30 fueron: 0.76, 0.73 y 0.88, respectivamente. Ello implica una alta asociación de las medias familiares entre ambientes. Las estimaciones de las correlaciones genéticas entre ambientes (Cuadro 3) con el empleo de ambos modelos no fueron diferentes de uno (P > 0.05), ello difiere de la correlación genética (0.54 ± 12) propuesta por Ibarra y Famula,6 en un estudio donde omitieron incluir los efectos comunes de FH en el modelo, lo que pudo haber generado una inadecuada estimación de los parámetros.9,18
En este contexto, Gitterle et al.8 estimaron correlaciones de valores genéticos familiares entre ambientes superiores a 0.97 para el peso corporal en diferentes granjas comerciales de Colombia, con densidades que variaron entre 18 y 22 camarones/m2, pero en su estudio no estimaron la correlación entre efectos comunes de FH; estos resultados concuerdan con las correlaciones genéticas obtenidas con el modelo RG del presente estudio. Castillo–Juárez et al.9 estimaron correlaciones genéticas que variaron entre 0.80 y 0.86 entre el peso corporal a los 130 días, medido en densidades de siembra de 9 y 14 camarones/m2 en dos localidades diferentes y correlaciones entre efectos comunes de FH que variaron entre 0.93 y 0.94.
La correlación de los efectos comunes de FH (Cuadro 3) entre KINO85 y POZOS30 no fue diferente de uno (P > 0.05), en tanto que entre POZOS10 y POZOS30 la correlación fue estimada en el límite del espacio parametral, con valor cercano a uno (0.99), estos valores son consistentes con lo encontrado por Castillo–Juárez et al.9 Sin embargo, en el presente estudio la correlación entre KINO85 y POZOS10 fue menor a uno (0.79 ± 0.09), que puede implicar que los efectos comunes de FH, que son mezcla de efectos ambientales, genéticos maternos y genéticos no aditivos,19 se manifiestan de diferente manera entre algunos ambientes de producción de camarón.
Los resultados obtenidos demuestran que no existe interacción genotipo por ambiente para el peso corporal a los 130 días en los ambientes estudiados. El ordenamiento de los valores genéticos predichos de las familias y la estimación de los parámetros genéticos son constantes a través de los ambientes estudiados, por lo que la selección de reproductores no se ve afectada.
Los resultados son importantes para el diseño de programas de selección que tienen como propósito el mejoramiento genético de P. vannamei para peso corporal, porque sugieren que no es necesario hacer programas específicos de mejora genética de acuerdo con la densidad de siembra dentro del rango estudiado en este trabajo (10–85 camarones/m2). La necesidad de hacer programas específicos para diferentes niveles ambientales podría incrementar los costos por unidad de progreso genético.
En la estimación de parámetros genéticos para peso corporal de P. vannamei en diferentes ambientes, es importante incluir la covarianza entre los efectos comunes de familia de hermanos, ya que ello afecta de manera importante la estimación de las heredabilidades y de la proporción de efectos comunes de familia de hermanos.
]]> Agradecimientos
Los autores agradecen el apoyo recibido por parte del Consejo Nacional de Ciencia y Tecnología, de México, por la beca del primer autor durante sus estudios de doctorado (registro Conacyt número 192975). Este trabajo fue financiado por Maricultura del Pacífico, S.A. de C.V., y parcialmente por el Fondo Sectorial de Investigación en Materia Agrícola, Pecuaria, Acuacultura, Agrobiotecnología y Recursos Fitogenéticos Conacyt–Sagarpa–2005 (proyecto 1210). Se agradece en especial a Marcia Castillo Mendoza por la revisón realizada en esta traducción.
Referencias
1. Falconer DS, Mackay T. Introduction to quantitative genetics. 3rd ed. New York: Longman Scientific & Technical, 1996. [ Links ]
2. Montaldo HH. Genotype by environment interactions in livestock breeding programs: A review. Interciencia 2001;26:229–235. [ Links ]
3. Cuvin–Aralar MLA, Lazartigue AG, Aralar EV. Cage culture of the Pacific white shrimp Litopenaeus vannamei (Boone, 1931) at different stocking densities in a shallow eutrophic lake. Aquaculture Res 2009; 40:181–187. [ Links ]
4. Gall GAE, Bakar Y. Stocking density and tank size in the design of breed improvement programs for body size of tilapia. Aquaculture 1999;173:197–205. [ Links ]
5. Raux P, Bailly D. Literature review on world shrimp farming. Individual Partner Report for the Project: Policy Research for Sustainable Shrimp Farming in Asia. European Commission INCODEV, Project No. IC4–2001–10042. CEMARE University of Portsmouth UK and CEDEM, Brest, France, 2002: 46. [ Links ]
6. Ibarra AM, Famula TR. Genotype by environment interaction for adult body weights of shrimp Penaeus vannamei when grown at low and high densities. Genet Sel Evol 2008;40:541–551. [ Links ]
7. Perez–Rostro C, Ibarra MA. Heritabilities and genetic correlations of size traits at harvest size in sexually dimorphic Pacific white shrimp (Litopenaeus vannamei) grown in two environments. Aquaculture Res 2003;34:1079–1085. [ Links ]
8. Gitterle T, Rye M, Saltec R, Cock J, Johansen H, Lozano C et al. Genetic (co)variation in harvest body weight and survival in (Penaeus (Litopenaeus) vannamei) under standard commercial conditions. Aquaculture 2005;243:83–92. [ Links ]
9. Castillo–Juarez H, Quintana CJC, Campos–Montes GR, Cabrera VC, Martinez OA, Montaldo HH. Heritability for body weight at harvest size in the Pacific white shrimp (Penaeus (Litopenaeus) vannamei), from a multi–environment experiment using univariate and multivariate animal models. Aquaculture 2007;273:42–49. [ Links ]
10. Gilmour AR, Gogel BJ, Cullis BR, Welham SJ, Thompson R. ASReml User Guide. Release 1.0, Hemel Hempstead: VSN International Ltd, 2002. [ Links ]
11. Chow S, Sandifer PA. Differences in growth, morphometric traits, and male sexual maturity among Pacific white shrimp, Penaeus vannamei, from different commercial hatcheries. Aquaculture 1991;92:165–178. [ Links ]
12. Palomino G, Contreras F, Sanchez A, Rosas C. Density and water exchange–dependent growth and survival of Litopenaeus setiferus post larvae. J World Aqua–culture Soc 2001;32:167–176. [ Links ]
13. Araneda M, Perez EP, Gasca–Leyva E. White shrimp Penaeus vannamei culture in freshwater at three densities: Condition state based on length and weight. Aquaculture 2008;283:13–18. [ Links ]
14. Vieira C, Pasyukova EG, Zeng Z, Hackett JB, Lyman RF, Mackay TFC. Genotype–environment interaction for quantitative trait loci affecting life span in Drosophila melanogaster. Genetics 2000;154,:213–227. [ Links ]
15. Castillo–Juarez H, Oltenacu PA, Cienfuegos–Rivas EG. Genetic and phenotypic relationships among milk production and composition traits in prim–iparous Holstein cows in two different herd environments. Livest Prod Sci 2002;78:223–231. [ Links ]
16. Fikse WF, Rekaya R, Weigel KA. Genotype x environment interaction for milk production in Guernsey cattle. J Dairy Sci 2003;86:1821–1827. [ Links ]
17. Bagley MJ, Bentely B, Gall GAE. A genetic evaluation of the influence of stocking density on the early growth of rainbow trout (Oncorhynchus mykiss). Aquaculture 1994,121:313–326. [ Links ]
18. Robison OW. The influence of maternal effects on the efficiency of selection; A review. Livest Prod Sci 1981;8:121–137. [ Links ]
19. Gjerde B, Terjesen BF, Barr Y, Lein I, Thorland I. Genetic variation for juvenile growth and survival in Atlantic cod (Gadus morhua). Aquaculture 2004;236:167–177. [ Links ]
* Maricultura del Pacífico, S.A. de C.V.
** Northwest Marine Technology®, Estados Unidos de América.
]]>