,) uptake and carotenoids/chlorophyll a ratio we determined in Amphidinium carteri Hulburt, cultured under conditions of continuous light (50, 150, 300 and 750 μmol quanta m-2 s-1) and three nutrient concentrations NaNO3/NaH2PO4 at 441.5/18.1 μM (low) 883/36.3 μM (medium) and 1766/72.6 μM (high). Both nutrient and irradiance had a significant effect (p < 0.05) on cellular abundance during the period of culture, except for the fourth and seventh day for nutrients (p > 0.05). In cultures under low nutrient condition, NO- and PO4- were almost depleted by the fifth day and in cultures with medium nutrient this condition occurred in the sixth day; whereas, at high nutrient condition the nutrients were not depleted. We concluded that A. carteri had higher growth rates and nutrient consumption at 300 μmol quanta m-2 s-1 during the first five days and in general the effect of nutrients on the pigment ratios was not significant (p > 0.05). However, the average peridinin/Chla ratio decreased up to 72% from the lowest and the highest irradiance. The opposite was observed for the average of diadinoxanthin/Chla ratio that increased almost two-fold, and the averages dinoxanthin and diatoxanthin to Chla ratios that increased from low to high irradiances. The average diatoxanthin/Chla ratio at 750 μmol quanta m-2 s-1 increased up to 2.7-fold from the exponential to the stationary phase only in the low and medium nutrient concentration. Likewise, the average peridinin, dinoxanthin and diatoxanthin to Chla ratios were not significantly different at 50 and 150 μmol quanta m-2 s-1. These results indicate important changes in average carotenoids/Chla ratios in A. carteri cultured under different irradiances.]]>
,) y proporción de carotenoides/clorofila a en Amphidinium carteri Hulburt, cultivado durante 7 días, bajo condiciones de luz continua: 50, 150, 300 y 750 μmol quanta m-2 s-1 y tres concentraciones de nutrientes NaNO3/NaH2PO4 (441.5/18.1 μM (bajo); 883/36.3 μM (medio) y 1766/72.6 μM (alto). La abundancia celular fue significativamente afectada (p < 0.05) tanto por el nivel de nutrientes como por la irradianza durante el periodo de cultivo, excepto para el nivel de nutriente en el cuarto y séptimo día. En los cultivos con bajo nutriente, el se agotaron casi totalmente al quinto día, y en cultivos con medio nutriente ocurrió al sexto día; mientras que en alta concentración de nutrientes, éstos no se agotaron. Se concluye que A. carteri tuvo la mayor tasa de crecimiento y consumo de nutrientes en 300 μmol quanta m-2 s-1 durante los primeros cinco días y en general el efecto de nutrientes sobre las proporciones de pigmentos no fue significativo (p > 0.05). Sin embargo, la proporción promedio de peridinina/Chla disminuyó hasta un 72% de bajas a altas irradianzas. Lo contrario ocurrió para el promedio de las proporciones de diadinoxantina/Chla, que se incrementó casi 2 veces, y los promedios de las proporciones de dinoxantina/Chla y diatoxantina/Chla se incrementaron de altas a bajas irradianzas. Los promedios de las proporciones de diatoxantina/Chla en 750 μmol quanta m-2 s-1 aumentó casi 2.7 veces de la fase exponencial a la estacionaria sólo en la condición baja y media de nutrientes. Así mismo, no hubo diferencias significativas en los promedios de las proporciones de peridinina, dinoxantina y diatoxantina respecto a la clorofila a en las irradianzas de 50 y 150 μmol quanta m-2 s-1. Estos resultados indican cambios importantes en los promedios de las proporciones de carotenoides/Chla en A. carteri cultivada en diferentes irradianzas.]]>
Artículos
Macronutrient uptake and carotenoid/chlorophyll a ratio in the dinoflagellate Amphidinium carteri Hulburt, cultured under different nutrient and light conditions
Consumo de macronutrientes y proporción de carotenoides/clorofila a en el dinoflagelado Amphidinium carteri Hulburt, cultivado bajo diferentes condiciones de luz y nutrientes
Enrique Valenzuela–Espinoza,1 Roberto Millán–Núñez,2 Eduardo Santamaría–del–Ángel2 and Charles C. Trees3
1 Instituto de Investigaciones Oceanológicas. Universidad Autónoma de Baja California. Apdo. postal 453, Ensenada, Baja California, México.
2 Facultad de Ciencias Marinas. Universidad Autónoma de Baja California. Apdo. Postal 453, Ensenada, Baja California, México. E–mail:rmillan@uabc.edu.mx
]]>
3 NATO Undersea Research Centre La Spezia, Italy.
Recibido: 5 de febrero de 2010. Aceptado: 31 de marzo de 2011.
ABSTRACT
Population growth, macronutrient (,) uptake and carotenoids/chlorophyll a ratio we determined in Amphidinium carteri Hulburt, cultured under conditions of continuous light (50, 150, 300 and 750 μmol quanta m–2 s–1) and three nutrient concentrations NaNO3/NaH2PO4 at 441.5/18.1 μM (low) 883/36.3 μM (medium) and 1766/72.6 μM (high). Both nutrient and irradiance had a significant effect (p < 0.05) on cellular abundance during the period of culture, except for the fourth and seventh day for nutrients (p > 0.05). In cultures under low nutrient condition, NO– and PO4– were almost depleted by the fifth day and in cultures with medium nutrient this condition occurred in the sixth day; whereas, at high nutrient condition the nutrients were not depleted. We concluded that A. carteri had higher growth rates and nutrient consumption at 300 μmol quanta m–2 s–1 during the first five days and in general the effect of nutrients on the pigment ratios was not significant (p > 0.05). However, the average peridinin/Chla ratio decreased up to 72% from the lowest and the highest irradiance. The opposite was observed for the average of diadinoxanthin/Chla ratio that increased almost two–fold, and the averages dinoxanthin and diatoxanthin to Chla ratios that increased from low to high irradiances. The average diatoxanthin/Chla ratio at 750 μmol quanta m–2 s–1 increased up to 2.7–fold from the exponential to the stationary phase only in the low and medium nutrient concentration. Likewise, the average peridinin, dinoxanthin and diatoxanthin to Chla ratios were not significantly different at 50 and 150 μmol quanta m–2 s–1. These results indicate important changes in average carotenoids/Chla ratios in A. carteri cultured under different irradiances.
]]>
Se evaluaron el crecimiento poblacional, consumo de macronutrientes (,) y proporción de carotenoides/clorofila a en Amphidinium carteri Hulburt, cultivado durante 7 días, bajo condiciones de luz continua: 50, 150, 300 y 750 μmol quanta m–2 s–1 y tres concentraciones de nutrientes NaNO3/NaH2PO4 (441.5/18.1 μM (bajo); 883/36.3 μM (medio) y 1766/72.6 μM (alto). La abundancia celular fue significativamente afectada (p< 0.05) tanto por el nivel de nutrientes como por la irradianza durante el periodo de cultivo, excepto para el nivel de nutriente en el cuarto y séptimo día. En los cultivos con bajo nutriente, el se agotaron casi totalmente al quinto día, y en cultivos con medio nutriente ocurrió al sexto día; mientras que en alta concentración de nutrientes, éstos no se agotaron. Se concluye que A. carteri tuvo la mayor tasa de crecimiento y consumo de nutrientes en 300 μmol quanta m–2 s–1 durante los primeros cinco días y en general el efecto de nutrientes sobre las proporciones de pigmentos no fue significativo (p > 0.05). Sin embargo, la proporción promedio de peridinina/Chla disminuyó hasta un 72% de bajas a altas irradianzas. Lo contrario ocurrió para el promedio de las proporciones de diadinoxantina/Chla, que se incrementó casi 2 veces, y los promedios de las proporciones de dinoxantina/Chla y diatoxantina/Chla se incrementaron de altas a bajas irradianzas. Los promedios de las proporciones de diatoxantina/Chla en 750 μmol quanta m–2 s–1 aumentó casi 2.7 veces de la fase exponencial a la estacionaria sólo en la condición baja y media de nutrientes. Así mismo, no hubo diferencias significativas en los promedios de las proporciones de peridinina, dinoxantina y diatoxantina respecto a la clorofila a en las irradianzas de 50 y 150 μmol quanta m–2 s–1. Estos resultados indican cambios importantes en los promedios de las proporciones de carotenoides/Chla en A. carteri cultivada en diferentes irradianzas.
Palabras clave: Consumo de macronutrientes, proporción carotenoides/clorofila a, irradianza, Amphidinium carteri.
INTRODUCTION
Amphidinium carteri Hulburt, is a marine dinoflagellate that is frequently present in coastal waters under bloom conditions. This dinoflagellate possess chlorophyll a (Chla) and the accessory photosynthetic pigment peridinin (Jeffrey & Vesk, 1997). Some authors have noted that the pigment ratios vary as a function of light, nutrient status and specific growth rate (Goericke & Montoya, 1998; Henriksen et al., 2002). Field studies have demonstrated changes in pigment composition in relation to the structure and nutrient content of the phytoplankton community (Stón & Kosakowska, 2000; Stón & Kosakowska, 2002). Under laboratory conditions, changes in pigment ratios are induced by irradiance variability and nutrient limitation during exponential and stationary growth phases in batch cultures of marine phytoplankton (Henriksen et al., 2002). However, different species have distinct growth rates and pigment composition in response to light and nitrogen stress (Sciandra et al., 2000).
Evaluation of the phytoplankton pigment composition using high performance liquid chromatography (HPLC), Chemical Taxonomy (CHEMTAX) and supplementary microscopy analysis, have been conducted to identify phytoplankton species and functional groups by relating specific marker pigments to total Chla contents (Mackey et al., 1996; Fietz & Nicklisch, 2004; Llewellyn et al., 2005). Previous investigations utilizing A. carteri have focused on the light–harvesting system and energy transfer in the peridinin–chlorophyll–protein complex (Sharples et al., 1996; Lohuis & Miller, 1998; Damjanoviet al., 2000; Polívka et al., 2005). However, few studies have been conducted in relation to nutrient uptake and the individual pigment composition in this species under different growth conditions in laboratory cultures. In this respect, the objective of the present work was to evaluate daily changes in macronutrient uptake and carotenoid/chlorophyll a ratios of A. carteri cultured under different nutrient and light conditions.
MATERIALS AND METHODS
Amphidinium carteri cultures. The marine dinoflagellate A. carteri was obtained from the monoalgal culture collection of the Instituto de Investigaciones Oceanológicas of the Universidad Autónoma de Baja California. All nutrients in the f/2 medium (Guillard, 1975) except nitrogen and phosphorus were added to one liter of filtered seawater to obtain the following concentrations: 11.7 μM Na2EDTA–2H2O; 11.7 μM FeCl36H2O; 0.04 μM CuSO45H2O; 0.08 μM ZnSO47H2O; 0.05 μM CoCl26H2O; 0.9 μM MnCl24H2O; 0.03 μM Na2MoO4–2H2O; 0.1 mg thiamine HCl; 0.5 μg biotin; 0.5 μg vitamin B12. Silicate was omitted because it should only be used for the growth of diatoms. Cultures were started in 250 mL Erlenmeyer flasks containing 200 mL of medium. The flasks were autoclaved at 120 °C with 1.05 kg cm–2 of pressure for 15 minutes, and then inoculated with 10 mL of A. carteri culture to obtain an average initial population of 1.17 x 105 cells mL–1. The cultures were maintained during seven days at 150 μmol quanta m–2 s–1 of irradiance, 33 PSU of salinity and a temperature of 25 ± 1 °C.
Experimental setting. A factorial experiment was designed using four levels of continuous light (50, 150, 300 and 750 μmol quanta m–2 s–1) and three nutrient concentrations. Light was provided by fluorescent lamps (daylight 75 W); the irradiance was measured with a scalar photosynthetic active radiation (PAR) irradiance meter (4π sensor, Biospherical Instruments model QSL–100). NaNO3 and NaH2PO4 were added at concentrations of 441.5/18.1 μM (low), 883/36.3 μM (medium) and 1766/72.6 μM (high), keeping a nitrogen/phosphorus ratio of ~24. All experiments were carried out in 3 L Fernbach flasks (2.9 L of media, n = 2 for each treatment). These flasks were sterilized as described above. A volume of 200 mL of culture were obtained at the end of the exponential phase from the previous level Erlenmeyer and was added to each experimental unit under controlled conditions. The initial average cell densities at the experimental units was 1.22 ± 0.2 × 105, 1.15 ± 0.2 × 105, 1.13 ± 0.2 × 105 cells mL–1 for the low, medium and high nutrient conditions, respectively. The culturing temperature was 25 ± 1 °C, and the salinity was 33 PSU. The pH was measured daily using a pH meter (Altex) and adjusted between 7.3 and 8.5 by the addition of CO2 at a flow rate of 270 mL min–1. Each flask was hand shaken every 12 hours. The cell density was determined daily with a Coulter (Beckman Coulter) particle counter Multisizer 3 in duplicate. The specific growth rate (μ) day–1 was calculated as follows:
]]>
μ = [ln(N2) – ln(N1)]/(t2 – t1),
where t1 and t2 are the initial and final time, and N1 and N2 are the initial and final cell densities, respectively.
and uptake. Daily samples of 10 mL were taken to quantify nitrate and phosphate removed from the culture during the experiment. These samples were filtered through 25–mm fiber glass filters (GF/F) of 0.7 μM and stored immediately at –20 °C for later analysis. Prior to analysis, calibration curves were performed for each nutrient. The filtered water was used to determine nitrate and phosphate concentrations according to Parsons et al. (1985). The nitrate and phosphate uptake were determined by measuring the disappearance of the nutrient from the media each day. The uptake was calculated as follows:
V = Ci– Cf /t
Where V is the daily uptake rate (μM), Ciand Cfare the initial and final nutrient concentration respectively and t is the time period of uptake (d–1).
Pigment analysis. For Chla and carotenoids analysis, 10 mL samples were filtered through 25 mm GF/F glass filters during the first three days of the culture, and thereafter, only 5 mL were filtered up to the seventh day. The filters were frozen, stored in liquid nitrogen and analyzed within the next three months. Prior to the analysis, the filters were placed in 4 mL of 93% acetone, sonicated for 10 seconds and stored in the freezer for 24 hours. Subsequently, the samples were centrifuged at 2,000 × g for 5 minutes and then filtered through 0.2 μM filters. From the acetone extract, 100 uL were injected into a HPLC system according to the method described by Bidigare et al. (2003). The pigments measured were Chla, the photosynthetic carotenoid (PSC) peridinin, and the pho–toprotective carotenoids (PPC) diadinoxanthin, dinoxanthin and diatoxanthin.
Statistical analysis. Two–way analyses of variance were used to determine the individual and interactive effects of nutrients and irradiance on dependent variables (cellular density, nitrate and phosphate uptake and carotenoids/Chla ratios). For variables that were significantly affected by nutrients and/or irradiance a multiple comparison procedure of Tukey was performed (a = 0.05).
RESULTS
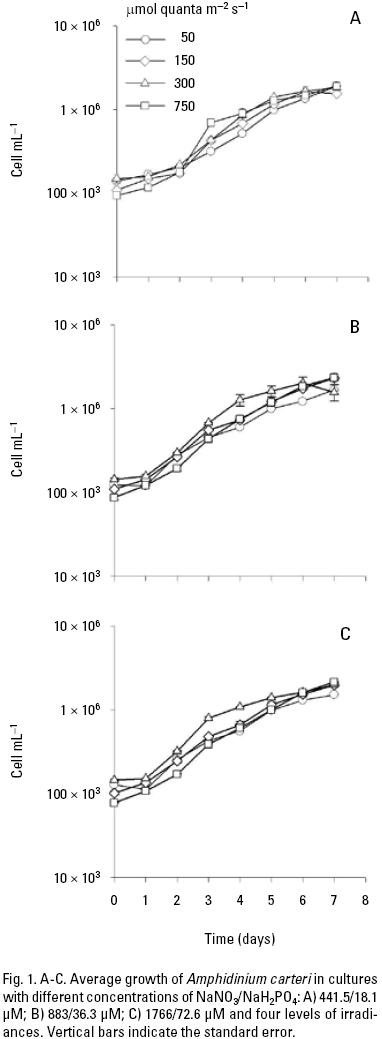
Microalgae cultures. The mean growth of A. carteri cultured under different nutrient and light conditions is shown in Fig. 1. The specific growth rate (μ) day–1 during the exponential phase was typically higher at high irradiance (0.49 to 0.66) than at low irradiances (0.42 to 0.63) per day. Subsequently, slow growth phase was observed with a reduction in specific growth rate during the latter part of the culture period in all treatments. These results indicated that the cultures had initiated the stationary growth phase (Fig. 1). The maximum cell densities at the low, medium and high nutrient concentrations were 19.1 ± 0.24 × 105, 23.3 ± 0.09 × 105 and 21.6 ± 0.008 × 105 cells mL–1, respectively and were observed at 750 μmol quanta m–2 s–1 (Fig. 1). However, during the first five days of the culture, the highest mean cell density was observed at 300 μmol quanta m–2 s–1, with a significant interaction between nutrient and irradiance in the majority of the days (Table 1A).
]]>
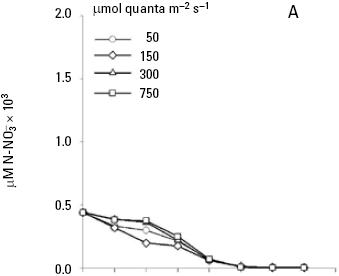
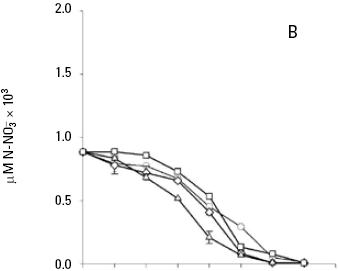
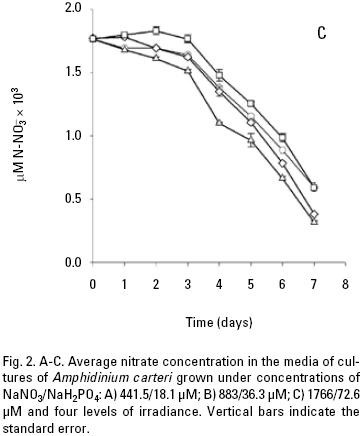
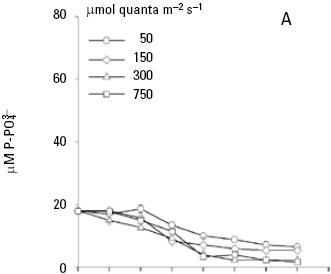
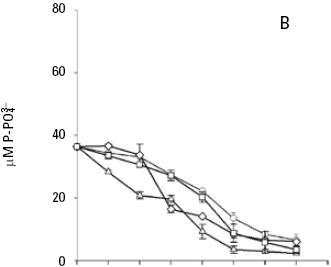
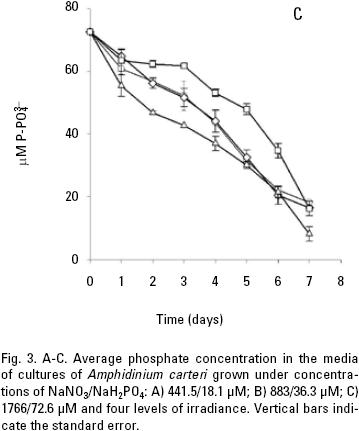
and uptake. Both nitrate and phosphate uptake was significantly affected by the levels of nutrients and light during the majority of the days of the culture, with a significant interaction between the factors only for nitrate uptake (Table 1B, C). The average nitrate and phosphate uptake during the first five days for low nutrient condition was 429.7 μM and 13.2 uM, which represents 98% and 93% of the total concentration of nitrogen and phosphate added to the culture respectively (Figs. 2A, 3A; Table 2). When cells were growing in the medium nutrient condition, in all light conditions, the average uptake was 744.4 μM of nitrate on the fifth day, representing 84% from the initial nutrient in the culture (Figs. 2B; 3B; Table 2); likewise the phosphate uptake was 27.6 μM representing the 76% from the initial nutrient in the culture. Cells growing at medium nutrient and 300 μmol quanta m–2 s–1 almost depleted the nutrients during the first five days, the nutrient uptake during this period of time was 817.5 μM of nitrate and 32.5 of phosphate representing the 92% and 90% of the initial content in the media, these results were higher than those observed at irradiances of 50, 150 and 750 μmol quanta m–2 s–1, for the same period of time. Under high nutrient concentrations and irradiances of 50 and 150 μmol quanta m–2 s–1, similar nutrient uptake were taken up from the media during the first five days (613.5–676.6 μM and 41.1–39.8 μM ). However, these values were lower than those observed in cultures that received 300 μmol quanta m–2 s–1 but higher than those in cultures exposed to an irradiance of 750 μmol quanta m–2 s–1, where only 575.6 and 24.4 μM of and were taken up from the media, these values representing the 32 and 33.6% of the initial content in the media (Figs. 2C; 3C; Table 2). A multiple comparison procedure showed that nitrate uptake by nutrient factor were significantly different (p < 0.05) from fourth to seventh day of culture, and the phosphate uptake between low and medium versus high, also were significantly different during the last two days of culture (p < 0.001).
]]>
After the fifth day in all light conditions, the nitrate was almost exhausted at low nutrient concentrations (Fig. 2A). However, at medium nutrient concentrations, depletion occurred after the sixth day (Fig. 2B). At high nutrient concentration neither nitrate nor phosphate was depleted (Figs. 2C; 3C). Comparison for light factor showed that the fourth day of culture the nitrate uptake at 300 μmol quanta m–2 s–1 was higher and significant with respect to the other irradiances (p < 0.05).
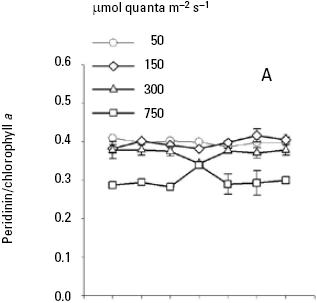
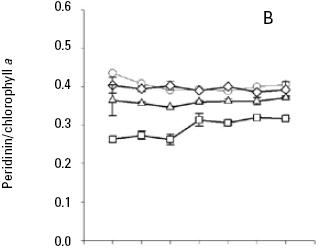
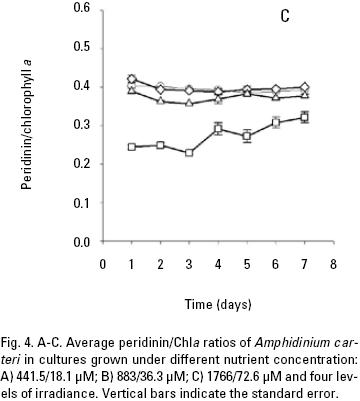
Pigment analysis. Both nutrient and light intensity had an effect on carotenoids/Chla ratios. Nutrient had not significant effect for diadinoxanthin/Chla ratio (p > 0.05; Table 3B) and light intensity had a significance effect for all carotenoids/Chla ratios (p < 0.001; Table 3, 4) at the end of the seven days experiment of A. carteri. For peridinin and dinoxanthin/Chla ratios a significant interaction was found between nutrient and light for the days 2–4 and 1–2 respectively (p < 0.05; Table 3A, 4A). Also a multiple comparison procedure (Tukey test) for factor nutrient showed that the average peridinin/Chla ratio on the second and third day was significantly different (p < 0.05) in high versus low nutrient. However, comparison for factor light among the different levels of nutrient indicated significant differences for all light conditions except for 50 and 150 μmol quanta m–2 s–1 (p > 0.05; Table 5).
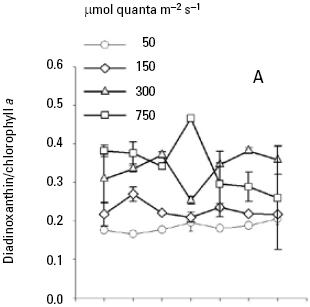
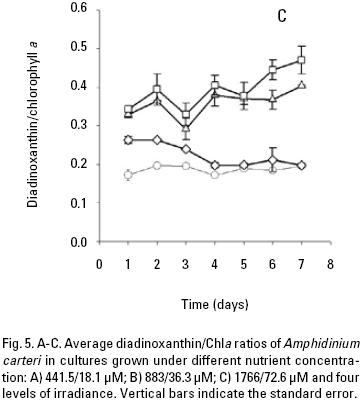
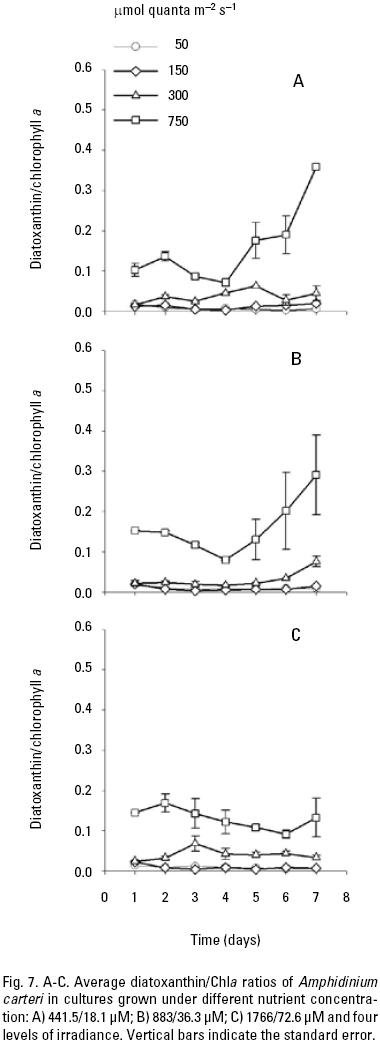
At low nutrient conditions and irradiance of 50, 150 and 300 μmol quanta m–2 s–1, the mean peridinin/Chla ratios were 0.39, 0.39, and 0.37 respectively, which were higher than at irradiance of 750 (Fig. 4A). Likewise, the average peridinin/Chla ratio in the different growth phase showed little variation among the different irradiancies. On the other hand, the averages diadinoxanthin, dinoxanthin, and diatoxanthin/Chla ratios were always greater in high irradiance than low irradiances (Fig. 5A, 6A, 7A). For diadinoxanthin/Chla ratios, a multiple range test suggested that for majority of these ratios was not significant different between 50 versus 150 and 300 versus 750 μmol quanta m–2 s–1 (p > 0.05; Table 5). However, the most conspicuous increase in the average of diadinoxanthin/Chla ratio was detected at an irradiance of 750 μmol quanta m–2 s–1, increasing up to 0.46 on the fourth day of culture (Fig. 5A), while averages dinoxanthin and diatoxanthin/ Chla ratios were higher at 750 μmol quanta m–2 s–1 (Fig. 6, 7) and almost all values were significantly different than the treatments that received a light intensity of 50; 150 and 300 μmol quanta m–2 s–1 (p < 0.05; Table 6).
]]>
]]>
When A. carteri was cultured under medium nutrient concentrations there was not significant different (p> 0.05) in the mean peridinin/Chla ratios (0.40 and 0.39) at 50 and 150 μmol quanta m–2 s–1 (Fig. 4B, Table 5). At an irradiance of 300, the average peridinin/Chla ratio was 0.36 and showed small variations during the time culture (Fig. 4B), and the ratio obtained at an irradiance of 750 μmol quanta m–2 s–1, was 0.26 during the first three days and then increased to 0.30 (Fig. 4B). A multiple range test indicated significant difference (p < 0.05; Table 5) between these treatments.
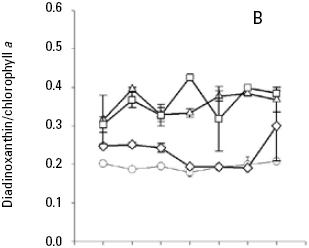
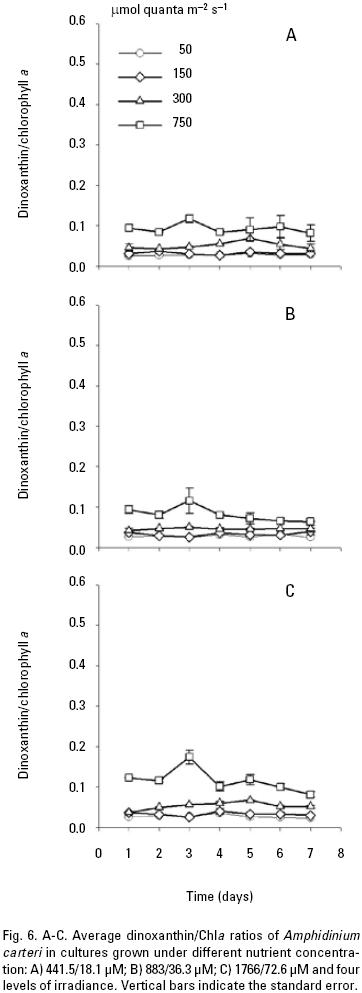
The average ratio of photoprotective pigments/Chla increased with increasing irradiance (Fig. 5, 6, 7). A multiple range test indicated that at irradiances of 300 and 750 μmol quanta m–2 s–1 the diadinoxanthin/Chla ratio was not significantly different except in the fourth day (p > 0.05; Table 5), but was higher than those obtained at low irradiances (Fig. 5B). The average values of the dinoxanthin/Chla and diatoxanthin/Chla ratios were smaller than the diadinoxanthin/Chla ratio (Fig. 6, 7) whereas averages di–noxanthin and diatoxanthin/Chla ratios at 750 μmol quanta m–2 s–1 were higher and significantly different than the treatments that received a light intensity of 50, 150 and 300 μmol quanta m–2 s–1 (p < 0.05; Table 6).
]]>
In high nutrient cultures (Fig. 4C), the average peridinin/Chla ratios (0.39) of A. carteri was not significantly affected by the irradiance at 50 and 150 μmol quanta m–2 s–1 (p > 0.05; Table 5). However, at 300 and 750 μmol quanta m–2 s–1, the average values of the peridinin/Chla ratios were 0.37 and 0.27 respectively (Fig. 4C) and a significant difference was observed (p < 0.05; Table 5). In addition, at 750 μmol quanta m–2 s–1 this ratio was 0.24 during the lag phase, which showed differences with respect to the ratios obtained during the exponential and stationary phases (0.26, 0.31) respectively (Fig. 4C). The average diadinoxanthin/Chla ratios at irradiances of 300 and 750 μmol quanta m–2 s–1 changed from the lag to the stationary phase from 0.34 to 0.38 and from 0.36 to 0.45 respectively, without significant differences (p > 0.05; Table 5) between irradiances. From the multiple range test results, we found that averages dinoxanthin and diatoxanthin/Chla ratios at 50 and 150 μmol quanta m–2 s–1 there were not a significant difference (p > 0.05) and these ratios showed little variation during the seven days of culture (Fig. 6, 7; Table 6).
DISCUSSION
Microalgae cultures. The cellular density and the growth rate of A. carteri may vary in response to interactions between irradi–ance and nutrient concentrations. Comparing all treatments, the ANOVA results indicated that the cellular density was affected both by the nutrient as irradiance, but mainly by the irradiance (Fig. 1; Table 1A). However, cells growing under different nutrient and light conditions showed low densities at the beginning of the culture period. This result indicates that A. carteri cells undergo a period of conditioning during the first two days of culture (Fogg & Thake, 1987). Likewise, the specific growth rate during the exponential phase was similar in all treatments with values from 0.40 to 0.66, these results agree with those reported by Sosik & Chisholm (1989). García and Purdie (1992) point out a maximum growth rate of 0.37 doubling day–1 for Gyrodinium cf. aureolum Hulburt grown under a light intensity of 235–380 μmol m–2 s–1 and at ~500 μmol m–2 s–1 the growth rate decreased. In contrast, Dae–IL et al. (2004) have found that the optimum irradiance for the growth of Cochlo–dinium polykrikoides Margalef is higher than 90 μmol m–2 s–1. In this study, the best response in the growth of A. carteri was at 300 μmol quanta m–2 s–1at the fifth day of culture.
and uptake. The uptake of nitrate and phosphate by A. carteri was affected for the different nutrient conditions. In low nutrient condition, during the first five days of culture the nitrate concentration was depleted, this result probably indicate a limitation of nitrate at the end of the culture period (Fig. 2A), which can lead to a decrease in protein synthesis (Bucciarelli & Sunda, 2003). Phosphate uptake showed variation with respect to different light levels, this has been observed by other authors who suggest that altering light status will change the uptake of phosphate (Lemasson et al., 1980; Nalewajko & Lee, 1983). But others, Lean & Pick (1981) point out that the phosphate uptake was not stimulated by light. However, the phytoplankton photosynthesis requires phosphate to synthesize ATP and NADPH, which are used in the the reduction and assimilation of nitrate (Clark & Flynn, 2002).
The results of the present work are also consistent with other studies (Clark & Flynn, 2002; Clark, 2001) that quantified the utilization of dissolved inorganic nitrogen and nitrate assimilation and found that the nitrate was exhausted between the fifth and sixth day. In addition, Needoba & Harrison (2004) have reported that NO3– uptake is modified by light and that the greatest concentrations of internal pools are detected during the dark period.
Pigment ratios. The average peridinin/Chla ratio was higher at irradiances of 50 and 150 than at 300 and 750 μmol quanta m–2 s–1. The converse was observed for the diadinoxantin, dinoxanthin, and diatoxanthin/Chla ratios, which were higher at 300 and 750 μmol quanta m–2 s–1. Considering the photoprotective functions of these pigments, their concentrations might be expected to co–vary with the amount of chlorophyll a and the irradiance, as was demonstrated in this work. Other studies have noted that in dinoflagel–lates the pigment diadinoxanthin is transformed into dinoxanthin under high light exposure and under limited light conditions, the peridinin concentration in A. carteri co–varies with Chla to absorb light (Jeffrey & Vesk, 1997) and transfer the absorbed energy to the reaction centers to carry out the photosynthetic process (Kirk, 1994). These observations are in agreement with the results reported by Schlüter et al. (2000) who cultured two species of dinoflagellates and found high concentration of peridinin in response to low light. Likewise, the peridinin/Chla ratio decreased with increasing irradiance due to a reduction in the cellular content of accessory pigments and Chla. Field studies have also confirmed that high concentrations of PSC are a result of chromatic adaptation to low light intensity and when phytoplankton are exposed to high irradiances, the cells produce pigments that are capable of protecting the reaction center against photo–oxidation and damage (Trees et al., 2000). On the other hand, Goericke & Montoya (1998) have pointed out that the pigment concentrations change as a function of the irradiance or the growth rate. The results of the present work indicated that the irradiance modified the pigment ratios, and a major effect was observed at 750 μmol quanta m–2 s–1, with changes detected between the exponential and stationary phases. However, changes in the growth rate at high irradiances, specifically at 300 μmol quanta m–2 s–1, improved the cell density in the cultures. This result might explain the changes observed in the growth rate with increasing light intensity, which were not observed when the nutrient concentration varied. Even though, it was not observed a limitation of the maximum population density achieved (Fig. 1), however the nutrient concentration observed at the end of the culture period, might explain that there is limited potential to increase cell yield, but the cell division continue even when the limiting nutrient is present at or near detection limit. This occurred in our experiments at the beginning of the stationary phase at low and medium nutrients (Figs. 2A, B; 3A, B). In addition, intracellular store of nitrate or phosphate might be sufficient for several hours or days of growth, whereas intracellular pools can be depleted (Leonardos & Geider, 2004). Likewise, the average carotenoid/Chla ratios were similar among the different nutrient conditions used to culture A. carteri. From our results we concluded that A. carteri had higher growth rates and nutrient consumption at 300 μmol quanta m–2 s–1 during the first five days of the experiment. The pigment ratios were affected both by nutrients and irradiance, but mainly by the irradiance. However, the average peridinin/Chla ratio decreased up to 72% from the lowest to the highest irradiance. The opposite was observed for the average of diadinoxanthin/Chla ratio that increased almost two–fold. Whereas, the average dinoxanthin and diatoxanthin to Chla ratios increased from low to high irra–diances, and these ratios were lower than the average diadino–xanthin/Chla ratio, instead the average diatoxanthin/Chla ratios at 750 μmol quanta m–2 s–1 increased almost 2.7–fold from the exponential to the stationary phase only in the low and medium nutrient concentrations. On the other hand, the average peridinin, dinoxanthin and diatoxanthin to Chla ratios there were not significantly different at 50 and 150 μmol quanta m–2 s–1 and similar results were observed for diadinoxanthin/Chla at 300 and 750 μmol quanta m–2 s–1. Our results indicate important changes in average carotenoids/Chla ratios in A. carteri cultured under different irradiances.
ACKNOWLEDGMENTS
The authors are grateful to CONACYT for supporting the first author during his graduate studies. This research was supported by the Universidad Autónoma de Baja California. Special thanks are extended to the Center for Hydro–Optics and Remote Sensing at San Diego State University for processing the HPLC samples. We also thank two anonymous reviewers, and special thanks to Elizabeth Ortega for their suggestions and positive criticisms.
]]>
REFERENCES
Bidigare, R. R., Van Heükelem, L. & Trees, C. C. 2003. HPLC phytoplankton pigments: Sampling, laboratory methods, and quality assurance procedures. In: Muller, J. L., Fargion, G. S., and McClain, C. R. (Eds.). Ocean Optics Protocols for Satellite Ocean Color Sensor, Revision 4, volume V. NASA Technical memorándum 2003–211621. NASA Goddard space Flight Center, Greenbelt, Maryland, pp. 5–14. [ Links ]
Bucciarelli, E. & W. G. Sünda. 2003. Influence of CO2, nitrate, phosphate, and silicate limitation on intracellular dimethylsulfoniopropionate in batch cultures of the coastal diatom Thalassiosirapseudonana. Limnology and Oceanography 48 (6): 2256–2265. [ Links ]
Clark, D. R. 2001. Growth rate relationships to physiological indices of nutrient status in marine diatoms. Journal of Phycology 37: 249–256. [ Links ]
Clark, D. R. & K. J. Flynn. 2002. N–assimilation in the noxious flagellate Heterosigma carterae (Raphidophyceae): dependence on light, N–source, and physiological state. Journal of Phycology 38: 503–512. [ Links ]
]]>
Dae–IL, K., Y. Matsüyama, S. Nagasoe & M. Yamagüchi. 2004. Effects of temperature, salinity and irradiance on the growth of the harmful red tide dinoflagellate Cochlodinium polykrikoides (Margalef) Dinophyceae. Journal of Plankton Research 26 (1): 61–66. [ Links ]
Damjanovi, A., T. Ritz & K. Schülten. 2000. Excitation transfer in the peridinin chlorophyll–protein of Amphininium carterae. Biophysical Journal 79: 1695–1705. [ Links ]
Fietz, S. & A. Nicklisch. 2004. An HPLC analysis of the summer phytoplankton assemblage in Lake Baikal. Freswater Biology 49: 332–345. [ Links ]
Fogg, G. E. & B. Thake. 1987. Algal cultures and phytoplankton ecology. Third Ed. University of Wisconsin Press. 257 p. [ Links ]
García, V. M. T. & D. A. Pürdie. 1992. The influence of irradiance on growth, photosynthesis and respiration of Gyrodinium cf. aureolum. Journal of Plankton Research 14 (9): 1251–1265. [ Links ]
]]>
Goericke, R. & J. P. Montoya. 1998. Estimating the contribution of micro–algal taxa to chlorophyll a in the field variations of pigments ratios under nutrient and light limited growth. Marine Ecology Progress Series 169: 97–112. [ Links ]
Guillard, R. R. L. 1975. Culture of phytoplankton for feeding marine invertebrates. In: Smith, W. L. & M. H. Chanley (Eds). Culture of marine invertebrate animals. Plenum Publishing Corp. New York. pp. 29–60. [ Links ]
Henriksen, P., B. Riemann, H. Kaas, H. M. Sørensen & H. L. Sørensen. 2002. Effect of nutrient–limitation and irradiance on marine phytoplankton pigments. Journal of Plankton Research 4 (9): 835–858. [ Links ]
Jeffrey, S. W. & M. Vesk. 1997. Introduction to marine phytoplankton and their pigment signatures. In: Jeffrey, S. W., R. F. C. Mantoura & S. W. Wright (Eds.). Phytoplankton Pigments in Oceanography. UNESCO, Paris, pp. 37–84. [ Links ]
Kirk, J. T. O. 1994. New York, NY, USA. Light and photosynthesis in aquatic ecosystems. 2nd Ed. Cambridge University Press. 509 p. [ Links ]
]]>
Lean, D. R. S. & F. R. Pick. 1981. Photosynthetic response of lake plankton to nutrient enrichment: a test for nutrient limitation. Limnology and Oceanography 26: 1001–1019. [ Links ]
Leonardos, N. & R. J. Geider. 2004. Response of elemental and biochemical composition of Chaetoceros muelleri to growth under varing ligh and nitrate:phosphate supply ratio and therir influence on critical N: P. Limnology and Oceanography 49 (6) : 2105–2114. [ Links ]
Lemasson, L., J. Pages & J. L. Cremoüs. 1980. Inorganic phosphate uptake in brackish tropical lagoon. Estuarine Coastal Marine Science 11: 547–561. [ Links ]
Llewellyn, C. A., J. R. Fishwick & J. C. Blackford. 2005. Phytoplankton community assemblage in the English Channel: a comparison using chlorophyll a derived from HPLC–CHEMTAX and carbon derived from microscopy cell counts. Journal of Plankton Research 27 (1): 103–119. [ Links ]
Lohuis, T. M. R. & D. J. Miller. 1998. Light–regulates transcription of genes encoding peridinin chlorophyll a protein and the major intrinsic light–harvesting complex proteins in the dinoflagellate Amphidinium carterae Hulburt (Dinophyceae). Changes in cytosine methylation accompany photoadatation. Plant Physiology 117: 189–196. [ Links ]
]]>
Mackey, D. M., D. J. Mcakey, H. W. Higgins & S. W. Wright. 1996. CHEM–TAX– a program for estimating class abundance from chemical markers: application to HPLC measurements of phytoplankton. Marine Ecology Progress Series 144: 265–283. [ Links ]
Nalewajko, C. & K. Lee. 1983. Light simulation of phosphate uptake in marine phytoplankton. Marine Biology 74: 9–15. [ Links ]
Needoba, J. A. & P. J. Harrison. 2004. Influence of low light and a light: dark cycle on NO3– uptake, intracellular NO3–, and nitrogen isotope frac–tionation by marine phytoplankton. Journal of Phycology 40: 505–516. [ Links ]
Parsons, T. R., Y. Maita & C. M. Lalli. 1985. A manual of chemical and biological methods for seawater analysis. First edition. (Eds.). Pergamon Press. Inc. New York. 173 p. [ Links ]
Polívka, T., T. Pascher, V. Sundström & R. G. Hiller. 2005. Tuning energy transfer in the peridinin–chlorophyll complex by reconstitution with different chlorophylls. Photosynthesis Research 86: 217–227. [ Links ]
]]>
Schlüter, L., F. Møhlenberg, H. Havskum & S. Larsen. 2000. The use of phy–toplankton pigments for identifying and quantifying phytoplankton groups in coastal areas: testing the influence of light and nutrients on pigments/chlorophyll a ratios. Marine Ecology Progress Series 192: 49–63. [ Links ]
Sciandra, A., L. Lizzara, H. Claustre & M. Babin. 2000. Responses of growth rate, pigment composition and optical properties of Cryptomonas sp. to light and nitrogen stresses. Marine Ecology Progress Series 201:107–120. [ Links ]
Sharples, F. P., P. M. Wrench, K. Ou & R. G. Hiller. 1996. Two distinct forms of the peridinin–chlorophyll a–protein from Amphidinium carterae. Biochimica et Biophysica Acta. Bioenergetics 1276 (2):117–123. [ Links ]
Sosik, H. M. & S. W. Chisholm. 1989. Chlorophyll fluorescence from single cells: Interpretation of flow cytometric signals. Limnology and Oceanography 34 (8): 1749–1761. [ Links ]
Stón, J. & A. Kosakowska. 2000. Qualitative and quantitative analysis of Baltic phytoplankton pigments. Oceanología 42 (4): 449–471. [ Links ]
]]>
Stón, J. & Kosakowska. 2002. Pigment composition in relation to phytoplankton community structure and nutrient content in the Baltic Sea. Oceanologia 44 (4): 419–437. [ Links ]
Trees, C. C., D. K. Clark, R. R. Bidigare, M. E. Ondrüsek & J. L. Mueller. 2000. Accessory pigments versus chlorophyll a concentrations within the euphotic zone: A ubiquitous relationship. Limnology and Oceanography 45 (5): 1130–1143. [ Links ]
 ) uptake and carotenoids/chlorophyll a ratio we determined in Amphidinium carteri Hulburt, cultured under conditions of continuous light (50, 150, 300 and 750 μmol quanta m-2 s-1) and three nutrient concentrations NaNO3/NaH2PO4 at 441.5/18.1 μM (low) 883/36.3 μM (medium) and 1766/72.6 μM (high). Both nutrient and irradiance had a significant effect (p < 0.05) on cellular abundance during the period of culture, except for the fourth and seventh day for nutrients (p > 0.05). In cultures under low nutrient condition, NO- and PO4- were almost depleted by the fifth day and in cultures with medium nutrient this condition occurred in the sixth day; whereas, at high nutrient condition the nutrients were not depleted. We concluded that A. carteri had higher growth rates and nutrient consumption at 300 μmol quanta m-2 s-1 during the first five days and in general the effect of nutrients on the pigment ratios was not significant (p > 0.05). However, the average peridinin/Chla ratio decreased up to 72% from the lowest and the highest irradiance. The opposite was observed for the average of diadinoxanthin/Chla ratio that increased almost two-fold, and the averages dinoxanthin and diatoxanthin to Chla ratios that increased from low to high irradiances. The average diatoxanthin/Chla ratio at 750 μmol quanta m-2 s-1 increased up to 2.7-fold from the exponential to the stationary phase only in the low and medium nutrient concentration. Likewise, the average peridinin, dinoxanthin and diatoxanthin to Chla ratios were not significantly different at 50 and 150 μmol quanta m-2 s-1. These results indicate important changes in average carotenoids/Chla ratios in A. carteri cultured under different irradiances.]]>
) uptake and carotenoids/chlorophyll a ratio we determined in Amphidinium carteri Hulburt, cultured under conditions of continuous light (50, 150, 300 and 750 μmol quanta m-2 s-1) and three nutrient concentrations NaNO3/NaH2PO4 at 441.5/18.1 μM (low) 883/36.3 μM (medium) and 1766/72.6 μM (high). Both nutrient and irradiance had a significant effect (p < 0.05) on cellular abundance during the period of culture, except for the fourth and seventh day for nutrients (p > 0.05). In cultures under low nutrient condition, NO- and PO4- were almost depleted by the fifth day and in cultures with medium nutrient this condition occurred in the sixth day; whereas, at high nutrient condition the nutrients were not depleted. We concluded that A. carteri had higher growth rates and nutrient consumption at 300 μmol quanta m-2 s-1 during the first five days and in general the effect of nutrients on the pigment ratios was not significant (p > 0.05). However, the average peridinin/Chla ratio decreased up to 72% from the lowest and the highest irradiance. The opposite was observed for the average of diadinoxanthin/Chla ratio that increased almost two-fold, and the averages dinoxanthin and diatoxanthin to Chla ratios that increased from low to high irradiances. The average diatoxanthin/Chla ratio at 750 μmol quanta m-2 s-1 increased up to 2.7-fold from the exponential to the stationary phase only in the low and medium nutrient concentration. Likewise, the average peridinin, dinoxanthin and diatoxanthin to Chla ratios were not significantly different at 50 and 150 μmol quanta m-2 s-1. These results indicate important changes in average carotenoids/Chla ratios in A. carteri cultured under different irradiances.]]> se agotaron casi totalmente al quinto día, y en cultivos con medio nutriente ocurrió al sexto día; mientras que en alta concentración de nutrientes, éstos no se agotaron. Se concluye que A. carteri tuvo la mayor tasa de crecimiento y consumo de nutrientes en 300 μmol quanta m-2 s-1 durante los primeros cinco días y en general el efecto de nutrientes sobre las proporciones de pigmentos no fue significativo (p > 0.05). Sin embargo, la proporción promedio de peridinina/Chla disminuyó hasta un 72% de bajas a altas irradianzas. Lo contrario ocurrió para el promedio de las proporciones de diadinoxantina/Chla, que se incrementó casi 2 veces, y los promedios de las proporciones de dinoxantina/Chla y diatoxantina/Chla se incrementaron de altas a bajas irradianzas. Los promedios de las proporciones de diatoxantina/Chla en 750 μmol quanta m-2 s-1 aumentó casi 2.7 veces de la fase exponencial a la estacionaria sólo en la condición baja y media de nutrientes. Así mismo, no hubo diferencias significativas en los promedios de las proporciones de peridinina, dinoxantina y diatoxantina respecto a la clorofila a en las irradianzas de 50 y 150 μmol quanta m-2 s-1. Estos resultados indican cambios importantes en los promedios de las proporciones de carotenoides/Chla en A. carteri cultivada en diferentes irradianzas.]]>
se agotaron casi totalmente al quinto día, y en cultivos con medio nutriente ocurrió al sexto día; mientras que en alta concentración de nutrientes, éstos no se agotaron. Se concluye que A. carteri tuvo la mayor tasa de crecimiento y consumo de nutrientes en 300 μmol quanta m-2 s-1 durante los primeros cinco días y en general el efecto de nutrientes sobre las proporciones de pigmentos no fue significativo (p > 0.05). Sin embargo, la proporción promedio de peridinina/Chla disminuyó hasta un 72% de bajas a altas irradianzas. Lo contrario ocurrió para el promedio de las proporciones de diadinoxantina/Chla, que se incrementó casi 2 veces, y los promedios de las proporciones de dinoxantina/Chla y diatoxantina/Chla se incrementaron de altas a bajas irradianzas. Los promedios de las proporciones de diatoxantina/Chla en 750 μmol quanta m-2 s-1 aumentó casi 2.7 veces de la fase exponencial a la estacionaria sólo en la condición baja y media de nutrientes. Así mismo, no hubo diferencias significativas en los promedios de las proporciones de peridinina, dinoxantina y diatoxantina respecto a la clorofila a en las irradianzas de 50 y 150 μmol quanta m-2 s-1. Estos resultados indican cambios importantes en los promedios de las proporciones de carotenoides/Chla en A. carteri cultivada en diferentes irradianzas.]]> ,
, ) uptake and carotenoids/chlorophyll a ratio we determined in Amphidinium carteri Hulburt, cultured under conditions of continuous light (50, 150, 300 and 750 μmol quanta m–2 s–1) and three nutrient concentrations NaNO3/NaH2PO4 at 441.5/18.1 μM (low) 883/36.3 μM (medium) and 1766/72.6 μM (high). Both nutrient and irradiance had a significant effect (p < 0.05) on cellular abundance during the period of culture, except for the fourth and seventh day for nutrients (p > 0.05). In cultures under low nutrient condition, NO– and PO4– were almost depleted by the fifth day and in cultures with medium nutrient this condition occurred in the sixth day; whereas, at high nutrient condition the nutrients were not depleted. We concluded that A. carteri had higher growth rates and nutrient consumption at 300 μmol quanta m–2 s–1 during the first five days and in general the effect of nutrients on the pigment ratios was not significant (p > 0.05). However, the average peridinin/Chla ratio decreased up to 72% from the lowest and the highest irradiance. The opposite was observed for the average of diadinoxanthin/Chla ratio that increased almost two–fold, and the averages dinoxanthin and diatoxanthin to Chla ratios that increased from low to high irradiances. The average diatoxanthin/Chla ratio at 750 μmol quanta m–2 s–1 increased up to 2.7–fold from the exponential to the stationary phase only in the low and medium nutrient concentration. Likewise, the average peridinin, dinoxanthin and diatoxanthin to Chla ratios were not significantly different at 50 and 150 μmol quanta m–2 s–1. These results indicate important changes in average carotenoids/Chla ratios in A. carteri cultured under different irradiances.
) uptake and carotenoids/chlorophyll a ratio we determined in Amphidinium carteri Hulburt, cultured under conditions of continuous light (50, 150, 300 and 750 μmol quanta m–2 s–1) and three nutrient concentrations NaNO3/NaH2PO4 at 441.5/18.1 μM (low) 883/36.3 μM (medium) and 1766/72.6 μM (high). Both nutrient and irradiance had a significant effect (p < 0.05) on cellular abundance during the period of culture, except for the fourth and seventh day for nutrients (p > 0.05). In cultures under low nutrient condition, NO– and PO4– were almost depleted by the fifth day and in cultures with medium nutrient this condition occurred in the sixth day; whereas, at high nutrient condition the nutrients were not depleted. We concluded that A. carteri had higher growth rates and nutrient consumption at 300 μmol quanta m–2 s–1 during the first five days and in general the effect of nutrients on the pigment ratios was not significant (p > 0.05). However, the average peridinin/Chla ratio decreased up to 72% from the lowest and the highest irradiance. The opposite was observed for the average of diadinoxanthin/Chla ratio that increased almost two–fold, and the averages dinoxanthin and diatoxanthin to Chla ratios that increased from low to high irradiances. The average diatoxanthin/Chla ratio at 750 μmol quanta m–2 s–1 increased up to 2.7–fold from the exponential to the stationary phase only in the low and medium nutrient concentration. Likewise, the average peridinin, dinoxanthin and diatoxanthin to Chla ratios were not significantly different at 50 and 150 μmol quanta m–2 s–1. These results indicate important changes in average carotenoids/Chla ratios in A. carteri cultured under different irradiances. et al., 2000; Polívka et al., 2005). However, few studies have been conducted in relation to nutrient uptake and the individual pigment composition in this species under different growth conditions in laboratory cultures. In this respect, the objective of the present work was to evaluate daily changes in macronutrient uptake and carotenoid/chlorophyll a ratios of A. carteri cultured under different nutrient and light conditions.
et al., 2000; Polívka et al., 2005). However, few studies have been conducted in relation to nutrient uptake and the individual pigment composition in this species under different growth conditions in laboratory cultures. In this respect, the objective of the present work was to evaluate daily changes in macronutrient uptake and carotenoid/chlorophyll a ratios of A. carteri cultured under different nutrient and light conditions.