Traslape espacial de la comunidad de peces dulceacuícolas diurnos en el sistema de humedal Media Luna, Rioverde, S.L.P., México
Spatial overlap of the diurnal freshwater fish community in the Media Luna wetland system, Rioverde, S.L.P., Mexico
Jorge Palacio–Núñez1,*, Genaro Olmos–Oropeza1, José R. Verdú2, Eduardo Galante2, Octavio César Rosas–Rosas1, Juan Felipe Martínez–Montoya1 y Jesús Enríquez3
1 Colegio de Postgraduados, Campus San Luis Potosí. Iturbide 73, Salinas de Hidalgo, S.L.P., 78600 México.*E–mail: jpalacio@colpos.mx
2 Instituto de Investigación de la Biodiversidad (CIBIO), Universidad de Alicante, Carretera San Vicente del Raspeig s/n, San Vicente del Raspeig, Alicante, 03690 España.
]]> 3 Colaborador externo. Ejército Nacional 100, Frac. Ignacio Loyola. Rioverde, S.L.P. 79610. México.
Recibido: 1 de junio de 2009.
Aceptado: 18 de diciembre de 2009.
RESUMEN
El humedal de la Media Luna es el más representativo y el más presionado por uso turístico y acciones de manejo en la Llanura de Rioverde, S.L.P. Contiene 13 especies de peces, dos nocturnas y 11 diurnas. Estas últimas fueron consideradas en este estudio, entre las que destacan cinco especies endémicas de la zona, una especie nativa de amplia distribución y cinco introducidas. Con la finalidad de determinar el grado de traslape espacial entre las especies, se tomaron los datos de cinco momentos de muestreo efectuados entre 1998 y 1999, con n = 54 transectos subacuáticos en cada muestreo. Se analizó tanto la distribución poblacional de los peces por clase de edad (adultos, juveniles y crías), como la similitud entre pares de especies para las variables del hábitat: sector, profundidad y vegetación. Los peces adultos y juveniles tendieron a traslaparse; las crías se segregaron en sector y se traslaparon en profundidad y vegetación. Hubo traslape considerable entre especies nativas e introducidas en al menos un estadio de vida; la especie endémica con mayor grado de superposición espacial con los introducidos fue Ataeniobius toweri, mientras que la especie introducida que mayor traslape con las nativas fue Oreochromis sp. Todas las especies endemícos se encontraron en riesgo de extinción, por lo que se considera de especial interés la posible afectación por el traslape espacial con las especies introducidas.
Palabras clave: Traslape espacial, análisis de comunidades, distribución de comunidades, peces endémicos, peces exóticos.
ABSTRACT
]]> The Media Luna wetland is the most representative and most affected by tourism and management actions in the Llanura de Rioverde, S.L.P. It contains 13 fish species, two of them are nocturnal and eleven diurnal. This study considers only the diurnal species. From these, the most relevant are five local endemic, one native widely distributed and five introduced species. In order to determine the degree of spatial overlap, 54 sub–aquatic transects per sampling session were conducted from 1998 and 1999. The fish population distribution per age class (adult, juvenile and larvae) and the similitude between species pairs were analyzed and estimated for the habitat variables: sector, depth, and vegetation. Adult and juvenile fish tended to overlap for all the variables, whereas the larvae segregated for sector but overlap for depth and vegetation. There was a high overlap between native and introduced species for at least one their life stages. The endemic species that showed the highest spatial overlap with the introduced species was Ataeniobius toweri, whereas Oreochromis sp. was the introduced one with the highest overlap with the native species. All of the endemic species are endangered; was thus, the possible affectation caused by the spatial overlap between endemic and introduced species has a special interest.Key words: Niche overlap, community analysis, communities distribution, endemic fishes, exotic fishes.
INTRODUCCIÓN
Los medios dulceacuícolas suelen ser ricos en biodiversidad y contener comunidades adaptadas a una gama de condiciones, tales como las situaciones que suelen presentarse en los manantiales de las zonas áridas, con alto contenido de solutos y grandes fluctuaciones térmicas (Contreras–Balderas, 1969). La distribución de los seres vivos depende de factores de diversa índole que abarcan escalas espaciales y temporales muy distintas, desde globales hasta locales, o debidas a interacciones. Las comunidades ecológicas son inherentemente complejas por la permanente existencia de un número importante de variables del hábitat que influyen sobre las especies, y la gran cantidad de especies que coexisten e interactúan. En general, las interacciones son amplias y diversas, involucrando a gremios multi–específicos de competidores que, en conjunto, son un mecanismo importante que estructura a las comunidades naturales (Albrecht & Gotelli, 2001; Palmer et al., 2003).
Para el estudio de especies que interactúan es fundamental el enfoque hacia el estudio de traslape de nicho. Se sugiere que al menos existen tres dimensiones principales que determinan el nicho: espacio, recurso trófico y tiempo (Pianka, 1969). Para el estudio de traslape de nicho usualmente se usa el término "eje de recursos" o "eje de nicho" para referirse al recurso particular por el cual dos o más especies compiten o comparten. Se considera que en todos los casos hay un número grande de estos ejes que las especies se reparten entre sí, siendo difícil saber cual o cuales deben ser medidos (Sale, 1974; Feinsinger et al., 1981). Schoener (1974) argumenta que el microhábitat, la dieta y la actividad temporal son los ejes de nicho más importantes, y que la mayor diferenciación ocurre a lo largo de los dos primeros ejes. Numerosos estudios son enfocados sobre gradientes ambientales, tales como microhábitat, y entonces los nichos quedan referidos como preferencias diferenciales del hábitat (Dolédec et al., 2000).
En la región semiárida de la Llanura de Rioverde, en el NE de México, existe una serie de al menos 16 manantiales que forman pequeños lagos. De estos, el humedal originado por el manantial de la Media Luna es el mayor de la zona y el más representativo, tanto en diversidad paisajística como en presión antrópica y condiciones de manejo inadecuadas. Este sistema se encuentra bajo constantes modificaciones para favorecer la actividad turística, con afectación a las poblaciones de al menos 13 especies de peces, debida a mala gestión y a presión del turismo. La sobrevivencia de estas especies ha estado en función de su propia tolerancia y adaptabilidad (Palacio–Núñez et al., 2007), sin embargo, la presión ejercida por las especies introducidas también ha manifestado afectación sobre las locales (Miller, 1984). El objetivo de este trabajo fue determinar el grado de traslape espacial, enfatizando en las interrelaciones entre especies nativas e introducidas, considerando a cada una de las especies en tres estadios de vida: adultos, juveniles y crías.
MATERIALES Y MÉTODOS
Descripción general del área de estudio. El sistema de la Media Luna se localiza en la Llanura de Rioverde, en el estado de San Luis Potosí (entre las coordenadas 21° 51' 34.29" – 21° 51' 47.67" N y 100° 00' 47.67" – 100° 01' 42.72" O. El lago–manantial de la Media Luna está formado por una continuidad de seis cráteres de manantial, con profundidad máxima de 36 m y flujo medio de 4350 l.seg–1. Del manantial parten tres canales principales de corriente muy lenta, que suman 86,200 m2 de espejo de agua. Con base en variaciones en la morfología de laguna y canales, por cambios en la profundidad del agua y en la cobertura vegetal subacuática, el sistema de la Media Luna fue dividido en 13 sectores (Figura 1).
]]> El criterio para establecer estratificaciones debe proponerse en función de las variables de cada hábitat particular. En peces, variables tales como la profundidad pueden ser consideradas como la mejor variable para estratificar el muestreo (Schwarz & Seber, 1999). En el humedal de la Media Luna la profundidad del agua (Fig. 2) tuvo gran relevancia para la distribución de la vegetación subacuática y para los peces (Palacio–Núñez et al., 2007).El fondo del sistema de la Media Luna presenta aproximadamente el 91% de cobertura vegetal de Nymphaea sp. en tres tipos morfológicos, el remanente está ocupado por suelo desnudo, propiciado principalmente por la actividad turística y en menor escala debido a una profundidad mayor a 12 m (Figs. 1 y 2). Los tipos morfológicos son: 1) alfombra pequeña, formada por plantas que presentan un tamaño menor a 0.25 m que crecen muy hacinadas, 2) alfombra grande, con plantas más dispersas y porte marcadamente mayor (0.5 – >2.00 m) y 3) tipo maduro, con plantas todavía más dispersas, porte similar a las anteriores pero que alcanzan la superficie con hojas flotantes y flores (Palacio–Núñez, 1997). La profundidad del agua (Fig. 2) fue de ocho niveles, considerando la distribución de las diferentes especies de peces en sus clases de edad: 1) 0.00 – 0.15 m, 2) 0.16 – 0.30 m, 3) 0.31 – 0.50 m, 4) 0.51 – 1.00 m, 5) 1.01 – 1.50 m, 6) 1.51 – 2.80 m, 7) 2.81 – 14.00 m y 8) > 14.00 m.
Muestreo de los peces. En la Media Luna habitan 13 especies de peces, de estas, Ictalurus mexicanus (Meek, 1904) e Ictalurus furcatus (Valenciennes in Cuvier Valenciennes, 1840) se ocultan durante las horas diurnas, la primera endémica de la región y la segunda introducida. En contraparte, las 11 especies restantes son activas durante ese mismo lapso y fueron las consideradas en este estudio, las cuales presentan diferente origen, distribución y riesgo de extinción y cuyas especificaciones se presentan en la Tabla 1.
Para la estimación de abundancia de peces, dos de los métodos más ampliamente usados actualmente son la pesca eléctrica o la observación directa mediante buceo (Bonar et al., 2000; Thompson, 2003). Se eligió el segundo método por ser recomendable para estimar distribución y abundancia de poblaciones en distintas circunstancias; además, se ha probado que tal método es eficiente, confiable, preciso y seguro (Ensign et al., 1995).
Para el presente trabajo se realizaron cinco sesiones de muestreo sistematizado mediante n = 54 transectos subacuáticos de ancho fijo, de 10 m de longitud por de 2 m de ancho, en cada sesión, distribuidos homogéneamente en todo el sistema, los cuales fueron trazados en forma perpendicular a la rivera para incluir los gradientes de profundidad y vegetación acuática (Ej. Palacio–Núñez et al., 2007). Los muestreos se hicieron en: primavera y verano de 1998, invierno 1998–1999 y verano y otoño de 1999. Las variables de campo, para cada avistamiento de los peces fueron: a) de tipo ambiental, cobertura de vegetación acuática, b) de tipo espacial, sector y profundidad del agua y c) de los peces, especie, clase de edad y abundancia, quedando la variable abundancia como respuesta general. Cabe mencionar que la talla corporal es un atributo importante desde un punto de vista ecológico y evolutivo. Algunas poblaciones con especímenes de diferente talla, o clase de edad, pueden comportarse como diferentes especies (Gotelli, 1997), lo cual es acentuado para el grupo de los peces (Werner & Gilliam, 1984), por lo que los muestreos fueron enfocados a las especies considerándolas en tres clases de edad: adultos, juveniles o crías.
Traslape espacial entre las especies. Numerosos autores se han enfocado al estudio de peces en función de sus interacciones, específicamente al traslape de nicho. En años recientes, varios de ellos se han apoyado en el programa EcoSim para estos fines (Ej. Fayram et al., 2005; Novakowski et al., 2008). Una de las mejores herramientas para estimar traslape de nicho es el módulo "Niche Overlap", que incluye el modo Pairwise para la comparación entre pares de especies, del programa informático EcoSim 7.0, el cual se basa en una tabla de medias que muestra la prueba de probabilidad en la cual el traslape de nicho medio observado se compara con el de las comunidades simuladas, asumiendo que cuando la media observada es mayor que la esperada hay traslape; en caso contrario, hay segregación (Gotelli & Graves, 1996).
La amplitud de nicho puede ser retenida o reasignada, cuando es retenida se preserva el grado de especialización de cada especie y cuando es reasignada usualmente conduce a un espectro mucho más amplio de utilización y, por lo tanto, un mayor traslape de nicho en la comunidad simulada contra la comunidad real. De la misma manera, los estados en cero son retenidos o reasignados: la participación de los ceros en la matriz observada es mantenida o no en cada matriz simulada.
Para el presente trabajo se realizaron comparaciones entre los algoritmos RA2, RA3 y RA4 utilizando el índice de Pianka. El algoritmo RA2 flexibiliza la amplitud del nicho observado y retiene la condición de ceros; RA3 retiene amplitud y flexibiliza ceros, mientras RA4 retiene ambas condiciones. Cada prueba fue corrida de manera aleatoria 1000 veces usando el programa informático EcoSim 7.0 (Gotelli & Entsminger, 2001). Para estos análisis se usó la abundancia total de peces por clase de edad. En todos los casos las filas contuvieron a las especies y las columnas a la presencia de los peces entre las variables del hábitat: profundidad (Prof), sector (Sec) y tipo de vegetación acuática (TipVeg), y estas tres variables combinadas (Prof+Sec+TipVeg).
RESULTADOS
]]> Durante el periodo de estudio las diferentes especies de peces, en sus tres clases de edad, no se distribuyeron de manera homogénea entre las variables del sistema de la Media Luna. En la Tabla 2 se muestra la abundancia media de cada una de las especies y la clase de edad entre los sectores; las áreas con vegetación muy poblada (en los sectores del 2 al 10) tuvieron las mayores abundancias. En la Tabla 3 se muestran datos similares para la distribución entre los niveles de profundidad y los tipos de cobertura subacuática; las mayores abundancias de crías y juveniles ocurrieron en profundidades someras (0.00 – 0.50 m y 0.16 – 1.00 m respectivamente), mientras los adultos fueron más abundantes entre 0.50 y 1.50 m. De los tipos de cobertura subacuática la mayor abundancia, en las tres clases de edad, ocurrió entre la alfombra grande y el tipo maduro.Referente al estudio espacial, para la comunidad de peces en estado adulto (Tabla 4), hubo traslape significativo para las variables profundidad (Prof) y tipo de vegetación acuática (TipVeg) y para la suma de las tres variables (Prof+Sec+TipVeg). Para la variable sector (Sec) los resultados fueron opuestos y el único algoritmo significativo fue RA2 mostrando segregación; donde la condición de los estados en ceros parece ser importante.
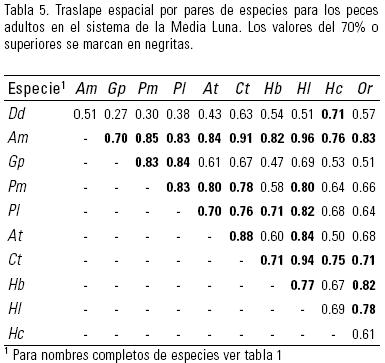
Se analizó en primer lugar la similitud entre los pares de especies de peces adultos pertenecientes a las nativas de este sistema (Tabla 5), encontrando traslapes de hábitat superiores al 70% entre la mayoría de las especies: A. mexicanus (Am), A. toweri (At), H. labridens (Hl), H. bartoni (Hb) y C. tessellatus (Ct). Sólo en el par para A. toweri con H. bartoni el traslape disminuyó a 60%. Dionda dichroma (Dd) mostró el traslape más bajo con las demás especies nativas.
Los adultos de las especies introducidas mostraron similitudes entre 51 y 84%. Los poecílidos G. panuco (Gp), P. latipunctata (Pl) y P. mexicana (Pm) tuvieron valores superiores a 80%. Los dos cíclidos introducidos, H. cyanoguttatum (Hc) y Oreochromis sp. (Or), mostraron menos traslape entre ellos (61%) y con los poecílidos (51 – 68%). En todos los casos, los valores fueron superiores a 50%.
En la comparación entre los adultos de las especies nativas con las introducidas, A. mexicanus se traslapó en más de 70% con todas las introducidas, mientras A. toweri sólo con Poecilia spp. (70 – 80%) al igual que C. tessellatus (76 –78%). Gambusia panuco habita preferentemente en profundidades muy bajas (Tabla 3) y la similitud con las demás especies disminuyó por el efecto de esa variable particular. Oreochromis sp. mostró fuerte similitud con H. bartoni (82%) y con H. labridens (78%).
Para los análisis de peces juveniles y crías no se consideró a D. dichroma, ya que durante los muestreos no fueron detectados individuos en estos estados (Tabla 2). Los peces juveniles tuvieron un patrón muy similar al de los adultos (Tabla 4). Los tres algoritmos mostraron traslape significativo para Prof, TipVeg y Prof+Sec+TipVeg mediante RA3, lo que enfatiza que varias especies de peces juveniles usan espacios en común, pero mediante RA2 se observó que no usan los mismos sectores.
La similitud entre pares de especies de peces juveniles nativos sólo fue considerable en el caso de A. mexicanus (75 – 91%) con las demás especies, así como entre C. tessellatus y H. labridens (89%). En todos los casos los porcentajes superaron el 60% (Tabla 6). Ataeniobius toweri fue el de menor traslape (55 – 68%) por su restricción ante los sectores carentes de vegetación (Tabla 3).
]]>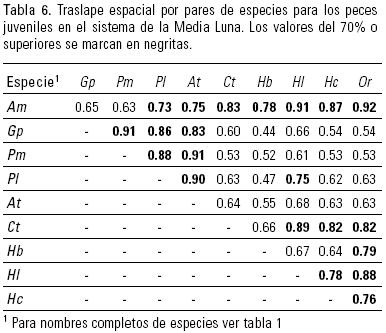
Para el traslape de juveniles entre las especies introducidas, se observaron porcentajes altos para los tres poecílidos (86 – 91%), así como entre H. cyanoguttatum y Oreochromis sp. (76%).
El traslape entre los juveniles de las especies nativas y las introducidas en general fue alto, salvo H. bartoni y G. panuco (44%) cuyas abundancias difieron en Prof y en Sec, todos los demás valores estuvieron por encima del 50%. Al igual que en adultos, A. mexicanus tuvo similitud alta con las especies introducidas (63 – 92%); para A. toweri fue mayor del 80% con los poecílidos, mientras Oreochromis sp. mostró niveles superiores a 78% con C. tessellatus, H. bartoni y H. labridens.
Las crías presentaron resultados contrastantes (Tabla 4), con segregación espacial significativa por RA2 para Prof y Sec, pero para Prof+Sec+TipVeg hubo segregación significativa mediante RA2 y traslape significativo por RA3 y RA4; ambas condiciones existen en función de las especies a las que pertenecen.
La similitud entre pares de especies nativas para las crías (Tabla 7) en general mostró valores menores que en adultos y juveniles, tales como A. mexicanus con H. bartoni (22%), H. labridens (29%) y C. tessellatus (15%) y A. toweri con H. bartoni (23%). Hubo valores más altos (60 – 62%) para los cíclidos nativos entre sí. Esto confirmó los valores generales de segregación de la Tabla 4.

Para las crías de las especies introducidas hubo traslape alto para los poecílidos entre sí (85 – 91%), pero valores muy inferiores para las demás combinaciones de especies (7 – 28%). Al igual que entre los nativos, predominó la segregación entre las especies.
Otro contraste de las crías fueron los valores entre nativos e introducidos muy inferiores, salvo para A. toweri con respecto a los poecílidos (70 – 85%) y para H. labridens respecto a Oreochromis sp. (85%). Para esto influyó también la segregación para Prof y Sec.
]]> DISCUSIÓN
Aunque desde 1992 hasta 2009 hubo eventuales y conspicuas fluctuaciones poblacionales debidas a acciones de gestión mal enfocadas, o por aparición de epizootias por motivos no antrópicos, en el periodo comprendido entre 1996 y 2000 no se detectó ninguno de estos eventos. Sin embargo, en el periodo de estudio (entre 1998 y1999), sin motivo detectando, hubo fluctuación significativa en la población general de peces entre la primavera y el verano de 1998, sin diferencias en las demás épocas de muestreo (Palacio–Núñez, 2007).
La regulación poblacional es un tema controvertido en ecología, ya que intervienen factores como la preferencia de las especies a determinado microhábitat, siendo difícil determinar las interrelaciones y otros procesos ecológicos que influyen en la distribución (Schoener, 1974; Feinsinger et al., 1981; Dolédec et al., 2000). Para una comunidad de peces es de esperar que las variables del hábitat (tamaño del cuerpo de agua, profundidad, tipo y velocidad de flujo, pH y solutos) expliquen una porción significativa de la varianza (Angermeier & Winston, 1999). En los bancos de depósitos con pendientes suaves y poca profundidad los peces pueden conseguir significativamente más comida, contrastando con las menores abundancias en sitios erosionados o a mayor profundidad (McIvor & Odum, 1998). En los sectores del 2 al 9 del sistema de la Media Luna se presentan estas condiciones, con muy alta cobertura vegetal subacuática. Es donde se registraron de manera abundante todas las especies, especialmente sus crías (con excepción de Oreochromis sp.), y es lo que explicó el traslape significativo mostrado por los peces adultos y juveniles para Prof y TipVeg en estos sectores. Sin embargo, existe una segregación significativa para las tres clases de edad respecto a Sec, lo que confirma que la abundancia no es homogénea dentro del sistema.
Se observó una separación de microhábitat entre los estadios de vida de una misma especie. Para la distribución de las crías de los cíclidos, el sitio es elegido por los padres que los protegen, siempre cerca del fondo y a mayor profundidad que las demás especies. Herichthys bartoni suele reproducirse en sitios abiertos; los demás cíclidos buscan algún elemento del paisaje que cubra algún flanco. Para las demás especies, las mayores concentraciones de crías se encontraron en sitios muy vegetados a escasa profundidad y esta combinación sólo se encuentra en algunos sectores, lo que explica una segregación significativa para Prof y para Sec, pero bajo esta condición, gran cantidad de crías de varias especies se traslapan.
Hay una estrecha relación entre las especies de los géneros endémicos de la Llanura de Rioverde, A. toweri y C. tessellatus, con similitud de hábitat de 88% entre adultos, 64% entre juveniles y 37% en crías. En estados adulto y en menor grado para juvenil, C. tessellatus es muy generalista, con amplia distribución entre sitios vegetados o no, y en profundidad, mientras A. toweri depende de sitios más someros y muy vegetados en sus tres estadios de vida (Palacio–Núñez, 2007).
Ataeniobius toweri está muy relacionado con las demás especies nativas, sobre todo en estado adulto, pero mostró traslape de consideración con Poecilia spp. en los tres estadios de vida (de 70 a 91%). Cualac tessellatus adulto se traslapa en más del 70% con Poecilia sp. y con los cíclidos introducidos; su relación con los poecílidos es menor en juveniles pero mayor con los cíclidos introducidos. Para C. tessellatus cría, el traslape es mucho menor con todos los introducidos. Ante estos resultados, se concluye que A. toweri es más susceptible que C. tessellatus ante la presencia de introducidos, ya que es más dependiente de la calidad del hábitat en sus tres estados de edad; además de ser la especie nativa con mayor traslape con las especies introducidas en la suma de todas sus etapas de vida.
Herichthys bartoni adulto mostró traslape espacial con los nativos H. labridens (77%) y C. tessellatus (71%) y con Oreochromis sp. de 82%. En estado juvenil sólo mostró traslape alto (79%) con este mismo cíclido introducido.
Los poecílidos mostraron valores de traslape entre sí mayores al 80% en adultos y al 85% en juveniles y crías, pero menores a lo que podría esperarse entre P. mexicana y P. latipunctata, dado que lo normal es que aparezcan en cardúmenes compartidos, las crías suelen compartir las mismas variantes del hábitat.
En el caso de D. dichroma no hubo suficientes datos para determinar el traslape; se puede suponer antagonismo con alguno de los introducidos, pero el único indicio es que sólo se observó en el sector 10, en ausencia de Oreochromis sp.
La introducción de especies es un problema creciente, particularmente cuando las especies nativas son incapaces de mantener la integridad con la presencia de las invasoras. El papel de estas introducciones ha crecido en proporciones pandémicas (Campbell et al., 2005) y la Media Luna no es la excepción. Todas las especies de peces son susceptibles a depredación constante al menos sus primeros estadios de vida y todas deben mantener un dinamismo reproductivo alto para sobrevivir. No hay datos de la población de los peces de la Media Luna en ausencia de especies introducidas, pero con estos resultados quedan establecidos los aspectos más relevantes sobre la situación ecológica en función de la distribución poblacional entre las variantes del hábitat. Los resultados muestran una abundancia poblacional crítica para D. dichroma y baja para H. bartoni, y se considera que la población de los endemismos no está al nivel más deseable, y que el grado de traslape con las especies introducidas es considerable.
]]>REFERENCIAS
Albrecht, M. & N. J. Gotelli. 2001. Spatial and temporal niche partitioning in grassland ants. Oecologia 126: 134–141. [ Links ]
Angermeier, P. L. & M. R. Winston. 1999. Characterizing fish community diversity across Virginia landscapes: prerequisite to conservation. Ecological Applications 9 (1): 335–349. [ Links ]
Bonar, S. A., B. D. Bolding, & M. Divens. 2000. Standard fish sampling guidelines for Washington State ponds and lakes. Report No. FPT 00–28, Washington Department of Fish and Wildlife, Olympia, Washington. 28 p. [ Links ]
Campbell, L M., S. B. Wandera, R. Thacker, G. D. Dixon & R. E. Heüky. 2005. Trophic niche segregation in the Nilotic ichthyofauna of Lake Albert (Uganda, Africa). Environmental Biology of Fishes 74: 247–260. [ Links ]
Contreras–Balderas, S. 1969. Perspectivas de la ictiofauna en las zonas áridas del norte de México. In: Memorias – Primer Simposio Internacional de Aumento de Producción en Zonas Áridas. ICASALS, Texas, Tech. Publ. 3. USA. pp. 293–304. [ Links ]
Dolédec, S., D. Chessel, & C. Gimaret. 2000. Niche separation in community analysis: a new method. Ecology 81 (10): 2914–1927. [ Links ]
Ensign, W. E., P. L. Angermeier & C. A. Dolloff. 1995. Use of line transect methods to estimate abundance of benthic stream fishes. Canadian Journal of Fisheries and Aquatic Sciences 52: 213–222. [ Links ]
Fayram, A. H., M. J. Hansen & T. J. Ehlinger. 2005. Interactions between walleyes and four fish species with implications for walleye stocking. North American Journal of Fisheries Management 25: 1321–1330. [ Links ]
Feinsinger, P., E. E. Spears & R. W. Poole. 1981. A simple measure of niche breadth. Ecology 62: 27–32. [ Links ]
Gotelli, N. J. 1997. Competition and coexistence of larval ant lions. Ecology 78(6): 1761–1773. [ Links ]
Gotelli, N. J. & G. R. Graves. 1996. Null models in ecology. Smithsonian Institution Press. Washington, DC. USA. 156 p. [ Links ]
Gotelli, N. J. & G. L. Entsminger. 2001. EcoSim: Null models software for ecology. Version 7.0. Acquired Intelligence Inc. & Kesey–Bear. http://homepages.together.net/~gentsmin/ecosim.htm. [ Links ]
IUCN, 2009. IUCN Red list of Threatened Species. Version 2009.1.www.iucnredlist.org. [ Links ]
McIvor, C. C. & W. E. Odum. 1998. Food, predation risk, and microhabitat selection in a marsh fish assemblage. Ecology 69 (5): 1341–1351. [ Links ]
Miller, R. R. 1984. La Media Luna, San Luis Potosí, at edge of Chihuahua Desert, México. In: Pister, E.P (Ed.). Proceedings of the Desert Fishes Council. Volumes XVI–XVIII of Annual Symposia. Bishop, CA. USA. pp. 67–72. [ Links ]
Novakowski, G. C., N. S. Hahn & R. Fugi. 2008. Diet seasonality and food overlap of the fish assemblage in a pantanal pond. Neotropical Ichthyology 6(4):567–576. [ Links ]
Palacio–Núñez, J. 1997. Caracterización del ecosistema de la laguna Media Luna, Rioverde, S. L. P. y su influencia en la ornitofauna acuática y ribereña. Tesis de Maestría. Colegio de Postgraduados. Montecillo, México. 140 p. [ Links ]
Palacio–Núñez, J. 2007. Designación de zonas prioritarias de conservación en el Parque Estatal de la Media Luna, (México): utilización de aves y peces como bioindicadores. Tesis Doctoral. Instituto de Investigación de la Biodiversidad (CIBIO). Universidad de Alicante, España. 155 p. [ Links ]
Palacio–Núñez, J., J. R. Verdú, E. Galante, D. Jiménez–García & G. Olmos–Oropeza. 2007. Birds and fish as bioindicators of tourist disturbance in springs in semiarid regions in Mexico: a basis for management. Animal Biodiversity and Conservation 30 (1): 29–41. [ Links ]
Palmer, T. M., M. L. Atanton & T. P. Young. 2003. Competition and coexistence: exploring mechanisms that restrict and maintain diversity within mutualist guilds. The American Naturalist 162: S63–S79. [ Links ]
Pianka, E. R. 1969. Sympatry of desert lizards (Ctenotus) in Western Australia. Ecology 50: 1012–1030. [ Links ]
Sale, P. F. 1974. Overlap in resource use, and interspecific competition. Oecologia 17: 245–256. [ Links ]
Schoener, T. W. 1974. Resource partitioning in ecological communities. Science 185: 27–39. [ Links ]
Schwarz, C. J. & G. A. F. Seber. 1999. Estimating animal abundance: Review III. Statistical Science 14 (4): 427–456. [ Links ]
Thompson, W. L. 2003. Hankin and Reeves' approach to estimating fish abundance in small streams: limitations and alternatives. Transactions of the American Fisheries Society 132: 69–75. [ Links ]
Werner, E. E. & J. F. Gilliam. 1984. The ontogenetic niche and species interactions in size–structured populations. Annual Review in Ecology and Systematics 15: 393–425. [ Links ] ]]>