
Cofactor FeMco (M = Mo, V, Fe) en la nitrogenasa
Cofactor FeMco (M = Mo, V, Fe) in nitrogenase
Brenda Lizette Ruiz Herrera, Jorge Arturo Campos González Angulo, Norah Barba Behrens1
1 Departamento de Química Inorgánica, Facultad de Química, Universidad Nacional Autónoma de México, C.U. Coyoacán, México, DF. 04510, México. Correo electrónico: norah@servidor.unam.mx, isyobrend@hotmail.com y knowliege@yahoo.com.mx.
]]>Recibido: 21 de junio de 2007.
Aceptado: 9 de octubre de 2007.
Abstract
Nitrogen fixation is a process widely investigated due to its great relevance to several fields, especially in agriculture. Nitrogen fixation is the process, which, as a component of the biogeochemical nitrogen cycle, drives the conversion of atmospheric N2 into ammonium ions. The enzyme nitrogenase has been studied with the purpose of developing systems capable of emulating (mimicking) it. It has been shown that it is a metallo-enzyme with a cofactor, constituted by two different metallo-clusters, one iron-sulfur and the other an iron-metal cluster, where the second metal atom may be molybdenum, vanadium or iron itself. Several reaction mechanisms have been formulated based upon structural, theoretical and experimental evidence, but the comprehension of the whole process is quite far from being fulfilled.
Estudio de la fijación de nitrógeno a lo largo del tiempo (Leigh, 1999)
A lo largo de los años, por lo menos los últimos dos mil, los humanos hemos hecho uso de la fijación de nitrógeno en los procesos agrícolas, a pesar de no tener la comprensión adecuada del proceso involucrado.
La primera técnica de mejora agrícola fue la rotación de cultivos, la cual, aunque se sabe de su uso en el continente americano, se cree fue originada en el imperio romano y ampliada al resto de Europa en el siglo XI. Dicha técnica era completamente empírica, por lo que la primera asociación del nitrógeno con el enriquecimiento del suelo fue meramente casual.
]]> A finales del siglo XVII se empezó a usar nabo como alimento para ganado, observándose que los campos donde éstos habían sido sembrados, daban muy buenos resultados con los cultivos posteriores. Así, en 1694 se asoció el éxito de los campos de nabo con una sustancia encontrada en ellos denominada nitro.A pesar de esta evidencia, cuando en 1772 Rutherford logró aislar nitrógeno y éste fue reconocido como gas elemental en 1776 por Lavoisier, se encontró que su molécula era prácticamente inerte, por lo que muchos científicos descartaron la idea, surgida en los agricultores, de que las plantas eran capaces de fijar nitrógeno convirtiéndolo en amoniaco.
Sin embargo, después de la unificación alemana en 1872 y tras el incremento en el apoyo a la investigación científica por parte de la nueva nación —impulsada por el deseo de superar a las otras potencias como Francia y Gran Bretaña— los científicos Hellriegel y Wilfarth lograron probar la fijación biológica del nitrógeno en 1886.
Hellriegel y Wilfarth encontraron que la fijación del nitrógeno estaba relacionada con los nódulos de las raíces que crecían en las plantas, denominadas por los agricultores "fijadoras de nitrógeno". Dichos nódulos eran resultado de una infección en la planta por bacterias del género Rhizobuim, por lo que concluyeron que éstas eran los verdaderos agentes fijadores.
Al empezar la Primera Guerra Mundial, el interés por la reducción de nitrógeno molecular ya no fue el desarrollo de la agricultura, si no la obtención barata de amoniaco, y con ello de nitrato de amonio, para la generación de explosivos.
Desde muchos años atrás, se tenía el conocimiento de que el amoniaco podía ser sintetizado a partir de sus elementos predecesores. Sin embargo, también se sabía que para lograr esto empleando nitrógeno e hidrógeno molecular se requería de condiciones muy drásticas.
Debido a ello, científicos como Nernst y Otswald creían que la obtención de amoniaco a nivel industrial sería prácticamente imposible. Sin embargo, fue Fritz Haber quien en 1912 logró demostrar lo contrario, pudiendo sintetizar grandes cantidades de amoniaco a partir de nitrógeno molecular, proceso conocido como Haber-Bosch, debido a que fue este último quien desarrolló la técnica de altas presiones necesaria para llevarlo a cabo.
El uso del proceso Haber-Bosch para la obtención de amoniaco continúa siendo el más importante hasta nuestros días, ya que representa la manera más económica de obtener fertilizantes a gran escala. Sin embargo, debido a las condiciones en que la reducción se lleva cabo:
requiere de instalaciones muy costosas, tanto en inversión inicial, como en mantenimiento, inversión que muchos países en vías de desarrollo no están dispuestos a realizar, por lo que el desarrollo de otros procesos capaces de generar amoniaco a partir de nitrógeno molecular continúan siendo de gran importancia.
]]>Fijación bioquímica del nitrógeno (Lehninger, 1993, p. 690-692; Voet, 1995, pp. 343-355, pp. 774-775; Metzler, 1981, pp. 824-828)
Existe un importante número de organismos capaces de reducir la molécula de N2. Entre los más conocidos se encuentran las bacterias que se alojan en los nódulos de las raíces de leguminosas y algunos vegetales. La reducción realizada por éstos es posible gracias a que dichos organismos poseen una enzima denominada nitrogenasa.
Para poder hablar de la nitrogenasa primero tenemos que entender lo que son las enzimas: éstas son proteínas globulares solubles que catalizan reacciones biológicas en los organismos vivos. Una enzima es, en su forma más esquemática, un catalizador bioquímico.
Las enzimas actúan sobre sustancias denominadas sustratos, uniéndose a ellos por medio de interacciones tipo Van der Waals, electrostáticas e hidrofóbicas, o por medio de enlaces de hidrógeno, enlaces covalentes transitorios, reacciones ácido-base, reacciones de óxido-reducción o reacciones de transferencia de grupo. Forman complejos enzima-sustrato que dan como resultado productos de importancia biológica, así como la enzima regenerada.
Los complejos enzima-sustrato reaccionan generalmente a través de procesos unimoleculares, es decir, mediante reacciones cuya cinética es de primer orden. El mecanismo por el cual las enzimas actúan sobre los sustratos se conoce como 'hipótesis llave-cerradura', en la cual el sitio de unión sustrato-enzima está constituido por una 'grieta' en la superficie de la enzima, de forma complementaria a la superficie del sustrato; de igual manera, los aminoácidos que forman la enzima, están constituidos de forma que actúen atractivamente sobre el sustrato. Esto nos lleva a que, para que una enzima sea selectiva a un sustrato, debe existir entre ellos una complementariedad electrónica y geométrica.
La parte más importante de una enzima es el lugar donde se lleva a cabo la reacción con el sustrato, es decir su sitio activo, que en algunas de ellas se denomina cofactor. Éste puede estar constituido por iones metálicos o por moléculas orgánicas, en cuyo caso se denomina coenzima.
Los cofactores pueden estar asociados de manera permanente a la enzima, conociéndose entonces como grupos prostéticos, o de forma transitoria, denominándose cosustratos.
Las enzimas se clasifican en función del tipo de reacción que catalizan. De acuerdo con ello, en una primera clasificación podemos agruparlas en seis tipos de enzimas básicas: oxidorreductasas, transferasas, hidrolasas, liasas, isomerasas y ligasas. Así, las transferasas, por ejemplo, serán enzimas que catalicen reacciones de transferencia de grupos funcionales.
]]> Las nitrogenasas son enzimas oxidoreductasas que catalizan reacciones de tipo óxido-reducción, específicamente la reducción de nitrógeno molecular; son de especial importancia para las plantas ya que efectúan en ellas el proceso de fijación de nitrógeno convirtiéndolo en amoniaco fácilmente asimilable.De manera general, las nitrogenasas están formadas por dos componentes (Chan, Jongsun, 1993):
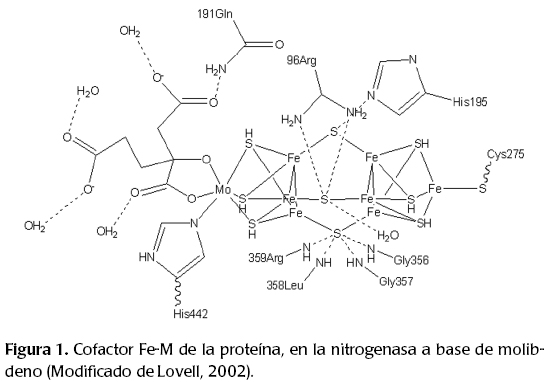
— La proteína Fe-M, donde el metal (M) puede ser molibdeno, vanadio o hierro. Es un tetrámero α2β2 constituido por 30 átomos de hierro y dos del metal correspondiente, distribuidos en dos tipos de cúmulos: los empaques cúbicos o tipo "P" (8Fe, 7S2-) y el cofactor Fe-M.
— La proteína sólo hierro, sensible al oxígeno, contiene únicamente cúmulos tipo 4Fe4S, así como dos sitios de enlace MgATP.
Para formar la enzima activa nitrogenasa se requiere una molécula del componente I o proteína Fe-M, y dos moléculas del componente II o proteína sólo hierro. En la figura 2 se muestran la proteína y el cofactor en su entorno proteico, aislada de la bacteria Klebsiella pneumoniae.
Las nitrogenasas se encuentran en bacterias como: Clostridium pasteurianum, Rhizobium y Azotobacterias. Las Rhizobia se encuentran en las raíces de las leguminosas, de manera que la fijación de nitrógeno se lleva a cabo en los nódulos de éstas; allí, la bacteria degenera en bacterioides y en una hemoglobina especial, denominada leghemoglobina, que atrapa el oxígeno, protegiendo a la nitrogenasa y proveyendo de éste al resto de la planta.
La reacción principal que efectúan las nitrogenasas es la reducción de nitrógeno molecular:
N2 + 8H+ + 8e–+ 16MgATP → 2NH3 + H2 + 16MgADP + 16P1
Sin embargo, es capaz de catalizar otras reacciones de reducción, como son:
]]>
No se ha determinado con exactitud la forma en que las nitrogenasas llevan a cabo el proceso de fijación; sin embargo, uno de los mecanismos más recientemente aceptados es el Lowe-Thorneley (1996), derivado de estudios cinéticos, y que está enfocado en la nitrogenasa Mo-Fe, ya que ésta ha sido la más estudiada. Se cree que el mecanismo es aplicable a todos los tipos de nitrogenasas.
El mecanismo supone un cambio en la configuración o estado de oxidación de la proteína Fe-Mo, debido a la transferencia de electrones de la proteína sólo hierro a los cúmulos "P", asociados al cofactor, en los cuales dichos electrones se "almacenan" hasta que son requeridos por el cofactor para la reducción del dinitrógeno; este proceso es energéticamente compensado por la hidrólisis de ATP en la proteína sólo hierro (Leigh, 2002, pp. 223-261).
En la figura 3 se muestra el esquema de reducción de la proteína Fe-Mo, en el que cada uno de los estados intermediarios corresponde a un ciclo en que la proteína sólo hierro cede un electrón a la Fe-Mo, a la par que lleva a cabo la hidrólisis de dos equivalentes de ATP. El ciclo completo involucra ocho electrones y 16 moléculas de ATP.

Según este mecanismo, los factores que influyen en la especificidad de la enzima, así como en la cantidad de nitrógeno reducido, frente a la de hidrógeno producido, son:
— El ambiente o aminoácidos que rodean al cofactor, los cuales participan en el proceso cediendo hidrógenos que se coordinan al metal presente en el cofactor, para después ser donados por éste a la molécula de dinitrógeno durante su reducción.
— El metal encontrado en el cofactor, ya que de las nitrogenasas existentes (Fe-Mo, Fe-V, Fe-Fe), la Fe-Mo es la que realiza más eficientemente el proceso de reducción.
]]> — El flujo de electrones, de manera que un alto flujo alto favorece la reducción, mientras que uno bajo da como único producto hidrógeno molecular.
Nitrogenasas y cofactores
De acuerdo con el metal representativo en el centro activo se distinguen tres nitrogenasas, cuya diferencia radica en el catión hexacoordinado presente en el cofactor Fe-M:
1. Nitrogenasa de Mo. La más abundante, encontrada en todas las especies que contienen esta enzima.
2. Nitrogenasa de V. La segunda en abundancia y en ser descubierta. Se encuentra en Azotobacter Vinelandii.
3. Nitrogenasa de Fe. Se encuentra en una proporción mucho menor a las anteriores. Aislada de Rhodobacter Capsulat.
En la representación dada en la figura 4 puede observarse que el sitio del heterometal (diferente de FeII) es octaédrico, lo que le confiere particularidades bien estudiadas desde el punto de vista de la química de coordinación.
]]>
De acuerdo con Lee (1997), en la nitrogenasa molibdeno, el heterometal en el cofactor tiene un estado de oxidación IV por lo que mantiene dos electrones en su capa d. Análogamente se tiene el escenario isoelectrónico del cofactor hierro-vanadio (FeVco), en el que el vanadio se encuentra en estado de oxidación III.
Para este tipo de especies se sabe que el sitio suele presentar paramagnetismo, al ser imposible aparear sus dos únicos electrones d. Las transiciones electrónicas de la proteína Fe-Mo, en medio ácido pueden observarse en el espectro de la figura 5.

Teniendo en cuenta las peculiaridades mencionadas para el sitio del heterometal, vale la pena describir las propiedades del cofactor completo como entidad, y diferenciarlas de acuerdo con el catión presente en el sitio octaédrico. La descripción que se da a continuación se enfoca en los efectos que produce la simple diferencia en la especie atómica que habita en el sitio de interés, por lo que no se hace una discusión más profunda sobre la diversidad de nitrogenasas conocidas y diferenciadas.
La presencia de FeII intrínseco en el sistema le confiere de propiedades particulares, pero no es sino hasta la inclusión de la especie en mayor estado de oxidación que éstas pueden ser discutidas. Los cationes que pueden ser sustituidos difieren en su configuración electrónica, de modo que la distribución de la densidad a lo largo del cofactor está marcadamente distinguida por el metal que distingue al cofactor. Esta dependencia del carácter electrónico repercute no sólo en las propiedades magnéticas, sino también en las estructurales, las cuales también se ven afectadas por las particularidades estéricas del catión que participa.
Propiedades electrónicas (Lovell, 2002)
Los estados basales de las nitrogenasas Fe-Mo y Fe-V exhiben un señal de EPR en S = 3/2, resultante de los cofactores, lo cual indica la existencia de tres electrones desapareados.
]]> De acuerdo con la fórmula propuesta (Lee, 1997), [MoIVFeIII6FeII9S2–]+ se tiene un conteo formal de 43 electrones d. Asumiendo que los iones metálicos son de alto espín, existen muchos patrones de alineación de espín que satisfacen S = 3/2. Considerando al MoIV, con configuración d2, en un entorno octaédrico de bajo espín con ligantes unidos por O, N y S, el estado de espín de S = ±3/2 resulta del apareamiento de un ión férrico (FeIII, d5, alto espín, S = 5/2) y seis iones ferroso (FeII, d6, alto espín, S = 2). La condición de tres electrones desapareados puede satisfacerse alineando los vectores de espín en los sitios Fe de la siguiente manera:
El anterior representa uno de varios posibles alineamientos de espín correspondientes a los valores Ms = 3/2 y una proporción de 4:3 con respecto a las direcciones de orientación de espín en los sitios Fe.
De acuerdo con la espectroscopia de absorción de rayos X, el vanadio en el FeVco puede estar como VII, VIII o VIV en un entorno de octaedro distorsionado, y a diferencia del FeMoco, el vanadio no presenta interacciones hiperfinas observables. Sin embargo, las configuraciones d 3 y d 1 introducen un número impar de electrones en el sitio heterometálico, junto con interacciones hiperfinas características del vanadio en diferentes estados de oxidación, además de no permitir que el sistema total de espín coincida con S = 3/2. Considerando la relación diagonal y las similitudes conocidas entre MoIV y VIII, la situación más probable es aquella en la que el sitio octaédrico es isoelectrónico (d2). La fórmula propuesta [VIIIFeIII6FeII9S2–] sigue muy de cerca al esquema de apareamiento de espín descrito para el FeMoco, con un estado de ruptura de simetría (BS = broken symmetry) de espín más probable, compuesto por un cúmulo V3Fe con espín Sa = 2, apareado antiferro-magnéticamente a un cúmulo 4Fe' con espín Sb = 7/2.
Se sabe que el estado basal del FeFeco es diamagnético (S = 0). Tomando en cuenta los estados de oxidación asignados por Lee (1997) al FeMoco, el cofactor de hierro cumple con la fórmula 2FeIII6FeII. Con las consideraciones anteriores, el que S = 0 surge del apareamiento antiferromagnético de los espines en los sitios Fe de la siguiente manera:

que también es sólo una de diversas alineaciones de espín posibles, todas correspondientes a Ms(total) = 0. Considerando que la simetría aproximada del cúmulo cumple con un C3, hay seis patrones simples de alineación de espín para el FeFeco, mostradas en la figura 6.

El estado BS3 representa el de menor energía; los otros cinco estados son excitados con entre 10.7 y 26.4 kcal/mol respecto al BS3. Se observa que el estado de menor energía es también el que presenta un mayor número de interacciones antiferromagnéticas, y se sabe que el estado de alineación ferromagnética es mucho más energético que los mostrados. Ello implica que el antiferromagnetismo juega un papel importante en la estabilización del cofactor hierro-hierro. Ya que las interacciones antiferromagnéticas son más energéticas en el caso Fe(III)/Fe(II) que en Fe(II)/Fe(II), el fenómeno observado se asocia con el carácter férrico de los sitios en los extremos.
]]> Con espectroscopia Mösbauer se ha encontrado un estado BS7 con dos interacciones ferromagnéticas y una interacción antiferromagnética a través de los átomos μS2 centrales en el cúmulo, lo que resulta del apareamiento antiferromagnético de dos mitades de cubano con espines Sa = 0 y Sb = 0. El estado BS3 surge del apareamiento antiferromagnético de dos mitades cubano con S = 4.
Propiedades estructurales (Lovell, 2002)
Las geometrías optimizadas de los cofactores FeMo, FeV y FeFe en el estado BS3 se comparan en la figura 7. En ésta se observa que el entorno químico permanece invariante, de modo que las estructuras observadas se mantienen considerablemente similares. Los parámetros geométricos, obtenidos mediante Teoría de Funcionales de Densidad, espectroscopia de absorción extendida de rayos-X de estructura fina y por caracterización biológica se comparan en la tabla 1.

La sustitución de MoIV por VIII en el cofactor da lugar a ciertos efectos estructurales, incluyendo una expansión de la longitud total del cofactor, de 7.27 a 7.62 Letra mayúscula A con diéresis Å. Lo anterior se explica tomando en cuenta que la carga total del cofactor cambia de –4 en el FeMoco a –5 en el FeVco, pues la repulsión electrónica distribuida por el cofactor se ve incrementada, lo que resulta en una ampliación en las longitudes VFe con respecto a las MoFe en el cubano M3Fe. Además, las distancias promedio Fe–Fe' y los ángulos Fe–μS 2-Fe' se incrementan en el congénere V. Sin embargo, las geometrías de los cubanos 4Fe' parece ser insensible al cambio total en la carga del cofactor.
Existe evidencia experimental de que los elementos estructurales principales en los FeMoco y FeVco también prevalecen en el FeFeco. Los parámetros geométricos en el FeFeco son ligeramente mayores que en el cofactor FeMoco. Esto se debe a que los orbitales 3d en el FeIII están contraídos con respecto a los 4d en el MoIV por lo que el traslape de orbitales entre el ión férrico hexacoordinado y los iones ferrosos está reducido con respecto al que se da entre estos últimos y el molibdeno. La expansión del FeFeco también se hace evidente en las distancia promedio entre los cubanos FeFe' (2.89 Å) comparada con el caso del FeMoco (2.75 Å). Como la distancias Fe–μS2 y Fe'–μS2 son casi idénticas a las observadas en el FeMoco y en el FeVco, la fuente de la expansión se adjudica a la abertura del ángulo Fe–μS2 Fe'.
El apareamiento antiferromagnético domina el esquema de espín en el FeFeco, particularmente a través de los sitios trigonales que ocupan la cavidad central del cúmulo. Las interacciones cortas del tipo Fe–Fe usualmente ocurren en pares de FeII apareados ferromagnéticamente donde se presenta una trayectoria de electrones σ deslocalizados con espín minoritario, como en el caso del FeMoco en estado BS6 (Lovell, 2001). Cuando en un par ferromagnético de centros Fe ocurre una inversión en uno solo de los vectores de espín, el apareamiento antiferromagnético prevalece entre los dos sitios y la trayectoria para los electrones σ deslocalizados se remueve. Con ello, los sitios Fe se mueven más lejos para dar una estructura del cofactor mucho más elongada.
El patrón de apareamiento BS3 que ocurre en el FeFeco tiene un efecto claro en la magnitud de las densidades de espín en los sitios Fe. Los sitios FeIII de alto espín ocupan las posiciones hexacoordinada y tetracoordinada con sus vectores de espín alineados de manera opuesta. Esta característica es consistente con la noción de que se requiere un número par de sitios entre férrico y ferroso para satisfacer el criterio diamagnético del cofactor.
]]> Si se toma la distancia entre la posición del heterometal con la de FeIII tetracoordinado como un indicador, la de FeFeco (7.87 Å) es significativamente mayor que las de los otros cofactores (7.27 Å en FeMoco y 7.62 Å en FeVco). La misma distancia para las diferentes alineaciones de espín en el FeFeco son: BS1 7.61 Å, BS2 7.41 Å, BS4 7.50 Å, BS5 7.40 Å y BS6 8.29 Å. La mayoría de estas distancias son menores que la de BS3 y se parecen más a las de los cofactores en las nitorgenasas molibdeno y vanadio; sin embargo, son de muy alta energía como para ser considerados como estados basales.
Reactividad (Hinneman, Nørskov, 2004)
En la mayoría de los casos la reactividad de las nitrogenasas alternativas disminuye en el orden:
Mo > V > Fe
Se observa que la nitrogenasa sólo hierro tiene la menor actividad. Además esta enzima cataliza una evolución de hidrógeno inusualmente elevada. Estas diferencias todavía no son comprendidas en términos de la estructura molecular; es decir, si surgen directamente del intercambio del metal o a las modificaciones estructurales a las que conduce.
Estudios teóricos han sugerido que los sitios de adsorción de H2 en el cofactor FeMo son el átomo μS2 y el interior del cúmulo, mediante un átomo de hierro, si bien no hay acuerdo sobre cuál es energéticamente más favorable, aunque para el FeFeco todos los cálculos concuerdan con que es el μS2 el sitio de unión; sin embargo, la energía del proceso se ha calculado con magnitudes tan diferente como 10 y 30 kcal/mol. No obstante, se ha observado experimentalmente que la nitrogenasa con molibdeno termina con una reducción de la His195, que se mantiene constante en la enzima sólo hierro. Esto hace pensar que los sitios de hierro en la sección trigonal forman parte importante del proceso, aun cuando los cálculos arrojan que la adsorción por estos sitios no es favorable.
En cuanto a la adsorción de nitrógeno, las consideraciones teóricas llevan a resultados que sugieren que la unión no se lleva a cabo, pues es endotérmica en todos los casos. Resultados más parecidos a la realidad se han conseguido modelando la acción de la histidina con una contribución de amonio.
En la figura 8 se muestran los resultados de la energía involucrada en la hidrogenación de la molécula de nitrógeno. Puede observarse que sin importar el modelo utilizado, la reacción con la nitrogenasa de molibdeno requiere mayor energía que la de sólo hierro.
]]>
Conclusiones
A pesar de los avances logrados en los últimos años, la fijación natural de nitrógeno es aún un proceso en aras de comprenderse. Aun tomando en cuenta las evasivas a las que los investigadores se enfrentan con la enzima en su estado natural, se han logrado realizar una infinidad de estudios analíticos y teóricos con el propósito de conocer el mecanismo mediante el cual la naturaleza logra llevar a cabo esta importante actividad.
La importancia de conocer a fondo la estructura y mecanismo de acción de la metaloproteína recae en la búsqueda de nuevos compuestos capaces de realizar la fijación de nitrógeno molecular de manera más económica y accesible, ya que como vimos, el proceso Haber-Bosch, por el que actualmente se lleva acabo, requiere de una costosa inversión inicial y de mantenimiento.
En el terreno de la química de coordinación se han logrado importantes avances en la obtención de análogos sintéticos a la acción de las nitrogenasas, desde la síntesis accidental del [Ru(NH3)5(N2)], compuesto que demuestra la coordinación del dinitrógeno a un centro metálico.
Un claro ejemplo lo proporciona el caso del ligante de campo débil tris(pirazolil)hidroborato (Berlinguette, 2006; Zuo, 2003), un ligante tridentado, coordinado por nitrógenos, usado para simular los ligantes unidos a los sitios oxidados en los cofactores. En combinación con otros ligantes, éste ha demostrado tener la capacidad de mantener la estructura del cúmulo análoga a la de la enzima. Sin embargo, al igual que otros compuestos sintetizados con la misma finalidad, es degradado en el proceso, por lo que su uso a nivel industrial se ve imposibilitado debido a los altos costos que genera.
Es por ello que el estudio de esta enzima natural y la búsqueda de compuestos análogos a su acción (Tatsumi, Ohki, 2007; Coucouvanis, 2007; Mayer, 1999) continúa siendo una importante área de investigación científica, con una inmediata aplicación y repercusión en la economía de países en vías de desarrollo.
Agradecimientos
]]> Este artículo no hubiera sido posible sin la participación del Prof. G.Jefferey Leigh y su invaluable conocimiento sobre el tema.De igual forma se agradece la valiosa aportación de Carlos Mauricio Maldonado Domínguez a la realización del artículo, así como de Gustavo Antonio Urrutia y de Edgar Omar Rodríguez Hernández por su apoyo en el diseño de los elementos gráficos.
Referencias
Arber,J. M.; Dobson, B. R.; Eady, R. R.; Stevens, P.; Hasnain, S. S.; Garner, C. D.; Smith, B. E., Vanadium K-edge X-ray absorption spectrum of the VFe protein of the Vanadium nitrogenase of Azotobacter chroococcum, Nature, 325, 372-374, 1987. [ Links ]
Berlinguette, C. P., Precursors to Clusters with the Topology of the PN Cluster of Nitrogenase: Edge-Bridged Double Cubane Clusters [(Tp)2Mo2Fe6S8L4]z: Synthesis, Structures, and Electron Transfer Series, Inorg. Chem, 45, 1997-2007, 2006. [ Links ]
Chan, Michael K.; Jongsun, Kim Rees, D. C., The Nitrogenase FeMo-Cofactor and P-Cluster Pair: 2.2 Å Resolution Structures, Science, 260, 792-794, 1993. [ Links ]
]]>Christiansen, J.; Tittsworth, R. J.; Hales, B. J.; Cramer, S. P. Fe and Mo EXAFS of Azotobacter vinelandii Nitrogenase in Partially Oxidized and Singly Reduced Forms, J. Am. Chem. Soc., 117, 10017, 1995. [ Links ]
Cotton, F. A.; Wilkinson, G.; Advanced Inorganic Chemistry, John Wiley, 5th ed., New York, 1988. [ Links ]
Coucouvanis, D. Exploration and Analyses of Possible Pathways for the Activation and Reduction of Dinitrogen by Nitrogenase, J. Biol. Inorg. Chem., 12, S157-S177, 2007. [ Links ]
Hinneman, B., J. K. Nørskov, Structure of the FeFe-cofactor of the iron-only nitrogenase and possible mechanism for nitrogen reduction, Phys. Chem. Chem.. Phys., 6, 843-853, 2004. [ Links ]
Krahn, E.; Weiss, B. J. R.; Krockel, M.; Groppe, J.; Henkel, G.; Cramer, S. P.; Trautwein, A. X.; Schneider, K.; Müller, A., The Fe-only nitrogenase from Rhodobacter capsulatus: Identification of the cofactor, an unusual, high-nuclearity iron-sulfur cluster, by Fe K-edge exafs and 57Fe Mössbauer spectroscopy, Biol. Inorg. Chem., 7, 37-45, 2002. [ Links ]
]]>Kraulis, P.J., MOLSCRIPT: A program to produce both detailed and schematic plots of protein structures, J. Appl. Crystalloggr., 24, 946, 1991. [ Links ]
Lee, H-I-; Hales, B.J.; Hoffman, B. M. Metal-Ion Valencies of the FeMo Cofactor in CO-Inhibited and Resting State Nitrogenase by 57Fe Q-Band ENDOR, J. Am. Chem. Soc., 119, 11395-11400, 1997. [ Links ]
Lehninger, Albert L., David L. Nelson, Principios de Bioquímica, Omega S.A., Barcelona, 1993. [ Links ]
Leigh, G. J., A Brief History of Nitrogen Fixation, Educ. Quím., 10(2), 86-91, 1999. [ Links ]
Leigh, G. J., Nitrogen Fixation at the Millennium, Elsevier, 2002. [ Links ]
]]>Lovell T., Metal Substitution in the Active Site of Nitrogenase MFe7S9 (M = MoIV, VIII, FeIII), Inorg. Chem., 41, 5744-5753, 2002. [ Links ]
Lovell, T.; Li, J.; Liu, T.; Case, D. A.; Noodleman, L., FeMo Cofactor of Nitrogenase: A Density Functional Study of States MN, MOX, MR, and MI, J. Am. Chem. Soc., 123, 12392-12410, 2001. [ Links ]
Mayer, S.M., Lawson, D. M., Gormal, C. A., Roe, S. M., Smith, B. E., New insights into structure-function relationships in nitrogenase: A 1.6 Å resolution X-ray crystallographic study of Klebsiella pneumoniae MoFe-protein, J. Mol. Biol., 292, 871-891, 1999 . [ Links ]
Metzler, David E., Bioquímica. Las reacciones químicas en las células vivas, Omega SA, Barcelona, 1981. [ Links ]
Tatsumi, K., Ohki, Y., A New Aspect of Fe/S-Cluster Synthesis. Relevance to the Active Site of Nitrogenase and Hydrogenase, J. Biol. Inorg. Chem., 12, S211-S213, 2007. [ Links ]
]]>Thorneley, R. N. F., Lowe, D.J., Nitrogenase: substrate binding and activation, J. Biol. Inorg. Chem., 1, 576-580, 1996. [ Links ]
Voet y Voet, Bioquímica, Omega SA, EUA, 1995. [ Links ]
Zumft, W. G., Isolation of Thiomolybdate Compounds from Molybdenum-Iron protein of Clostridial Nitrogenase, J. Biochem., 91, 345-350, 1978. [ Links ]
Zuo, J-L., H-C. Zhoi, R. H. Holm, Vanadium-Iron-Sulfur Clusters Containing the Cubane-type [VFe3S4] Core Unit: Synthesis of a Cluster with the Topology of the PN Cluster of Nitrogenase, Inorg. Chem., 42, 4624-4631, 2003. [ Links ]
]]>