
Productividad primaria del fitoplancton en la zona eufótica del Sistema de la Corriente de California estimada mediante imágenes del CZCS
Phytoplankton primary productivity in the euphotic zone of the California Current System estimated from CZCS imagery
OA Barocio-León, R Millán-Núñez*, E Santamaría-del-Ángel, A González-Silvera
Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Km 103 Carretera Tijuana-Ensenada, Ensenada CP 22800, Baja California, México. *E-mail: rmillan@uabc.mx.
]]> Recibido en mayo de 2006;
Resumen
Se estimó la productividad primaria (PP) del fitoplancton en la zona eufótica del Sistema de la Corriente de California utilizando la concentración de clorofila (Chl) obtenida de imágenes del CZCS (Coastal Zone Color Scanner). Se dividió el área en dos regiones, sur de California (SC) y Baja California (BC), cada una con una subregión costera y otra mar adentro, y en dos temporadas, la fría y la cálida. Los valores de Chl y PP fueron en general mayores y más variables en las subregiones costeras que mar adentro. La temporada fría mostró valores más altos de PP, excepto en la subregión de mar adentro de BC. El efecto de El Niño (ENSO) 1982-1983 fue más evidente en las subregiones costeras, propiciando valores más bajos de PP durante la temporada cálida de 1983 y la fría de 1984. En las subregiones costeras dominaron las aguas mesotróficas y eutróficas, mientras que en las subregiones de mar adentro la mayor contribución fue de aguas oligitróficas. En promedio, se estimó una PP total 23.78% debajo de los correspondientes datos in situ de CalCOFI, 51.82% en las subregiones costeras y 5.93% mar adentro, lo que concuerda con las diferencias esperadas entre datos in situ y los provenientes de modelos. El promedio diario de PP para la subregión costera y mar adentro de SC fue de 0.80 y 0.61 g C m-2, respectivamente, mientras que para la subregión costera y mar adentro de BC fue de 1.03 y 0.18 g C m-2, respectivamente. La PP total de la zona eufótica del área estudiada (de enero de 1979 a junio de 1986) fue de 2.39 Gt C. Se calculó una PP media anual de 0.32 Gt C para toda el área de estudio.
Palabras clave: productividad primaria, CZCS, clorofila a, Corriente de California.
Abstract
We estimated phytoplankton primary productivity (PP) in the euphotic zone of the California Current System using chlorophyll concentration (Chl) from CZCS imagery. The area was divided into two regions, Southern California (SC) and Baja California (BC), with inshore and offshore subregions, and into two seasons, cool and warm. The Chl and PP values were in general higher and more variable inshore than offshore. The cool season had higher PP values, except in the offshore subregion of BC. The effect of El Niño (ENSO) 1982-1983 was more evident inshore, leading to lower PP values during the warm season of 1983 and the cool season of 1984. Mesotrophic and eutrophic waters dominated in the inshore subregions, whereas the major contribution in the offshore subregions was from oligotrophic waters. The total PP estimated was on average 23.78% below the corresponding in situ CalCOFI PP data, 51.82% inshore and 5.93% offshore, which is within the expected agreement between in situ and modeling methods. The average daily PP for SC inshore and offshore was 0.80 and 0.61 g C m-2, respectively, and for BC inshore and offshore it was 1.03 and 0.18 g C m-2, respectively. The total PP in the euphotic zone of the study area (January 1979 to June 1986) was 2.39 Gt C. We calculated an annual average PP for the whole study area of 0.32 Gt C.
Key words: primary productivity, CZCS, chlorophyll a, California Current.
]]>Introducción
Uno de los objetivos de la oceanografía actual es medir y entender los procesos que controlan las fluctuaciones en los flujos de carbono a nivel global. Por tanto, la estimación de la productividad primaria (PP) del fitoplancton es de gran importancia, ya que es el proceso que transforma carbono inorgánico en materia orgánica mediante la asimilación fotosintética del CO2, introduciendo carbono en la cadena trófica acuática. Ya se ha determinado el papel de la PP en el ciclo del carbono, observándose que 75% de la diferencia en la concentración de carbono inorgánico disuelto entre la superficie y los océanos profundos se debe a la bomba biológica (Sarmiento y Bender 1994). Tales observaciones sugieren que la PP puede regular el creciente efecto invernadero causado por las emisiones antropogénicas de CO2 a la atmósfera desde la revolución industrial, lo que ha fomentado el interés de los científicos por estimar la tasa a la cual el carbono inorgánico se transforma en biomasa fitoplanctónica y es introducido en la cadena trófica marina a nivel global.
La concentración de clorofila ([Chl]) ha sido utilizada como indicador de la biomasa fitoplanctónica y, por tanto, la cuantificación de su distribución resulta esencial para la determinación de la PP. Aunque han mejorado los métodos para realizar estimaciones de [Chl] in situ, las técnicas clásicas de muestreo desde embarcaciones no son adecuadas para obtener datos de áreas extensas y de largo plazo. En consecuencia, existe incertidumbre alrededor de las características de la variabilidad planctónica a gran escala determinada mediante muestreos in situ (Longhurst et al. 1995).
El uso de sensores remotos para la detección de pigmentos fitoplanctónicos ha resultado ser la mejor forma de obtener una visión sinóptica de regiones extensas (Longhurst et al. 1995). Las imágenes de los sensores remotos proporcionan datos de la concentración media de pigmentos fotosintéticos en la primera profundidad óptica (Kirk 1994), lo que permite estudiar grandes áreas del océano de forma simultánea. Mediante estas imágenes se han observado filamentos, meandros y manchas irregulares, indicando mayores concentraciones cerca de la costa que mar adentro, con concentraciones máximas en las zonas de surgencia (Platt y Sathyendranath 1988, Longhurst et al. 1995).
El radiómetro conocido como Coastal Zone Color Scanner (CZCS) fue el primer sensor del color del océano instalado a bordo de un satélite en órbita (Nimbus 7) y funcionó de noviembre de 1978 a junio de 1986. Las imágenes que generó fueron usadas para estimar la concentración de clorofila a más feofitina a en la capa superior del océano dentro de cuadrículas muy finas sobrepuestas sobre áreas muy grandes (Longhurst et al. 1995). Esta capa superior corresponde a la primera profundidad óptica y puede variar de centímetros a varias decenas de metros, dependiendo de la concentración de los constituyentes del agua (Kirk 1994).
El uso de sensores remotos para estimar la PP requiere establecer una conexión entre un indicador de biomasa (concentración de pigmentos) y una tasa de fijación de carbono. Tal conexión se realiza mediante modelos matemáticos con base en la relación entre la producción primaria y la luz (e.g., Platt et al. 1988, Morel 1991, Behenfeld y Falkowski 1997). La capacidad del fitoplancton para utilizar la luz para transformar carbono inorgánico en materia orgánica se mide mediante la construcción de curvas de fotosíntesis-irradiancia (P-E). Se ha observado que los parámetros fotosintéticos que describen tales curvas varían en espacio y tiempo, y no pueden ser inferidas directamente de los datos tomados por sensores remotos. Como una forma de resolver este problema se sugirió la división del océano en provincias biogeoquímicas (Longhurst et al. 1995). Se podrían estimar los promedios de los parámetros fotosintéticos obtenidos de datos de campo para describir cada provincia biogeoquímica y utilizarse para estimar el PP a partir de las imágenes de satélite (Platt et al. 1991, Longhurst et al. 1995), o se podrían modelar como función de los sensores remotos (Morel et al. 1996).
A pesar de que las imágenes de satélite proporcionan una muy buena caracterización espacial de la capa superficial del océano, la distribución vertical de [Chl] en la zona eufótica no es homogénea y en general presenta un máximo profundo de clorofila (MPC) (Cullen y Eppley 1981), que a veces se localiza por debajo de la primera profundidad óptica y, por tanto, es invisible para el sensor. Para estimar la PP sobre toda la zona eufótica se necesita conocer la distribución vertical de los pigmentos, ya que suponer un perfil de biomasa homogéneo puede producir errores graves en las estimaciones (Platt et al. 1988). A fin de evitar este error potencial, Platt et al. (1988) propusieron un modelo de curvas gaussianas que describe la distribución vertical de [Chl]. Se han desarrollado modelos empíricos para estimar los parámetros gaussianos que describen tal curva a partir de la clorofila superficial para el Sistema de la Corriente de California (SCC) (Millán-Núñez et al. 1997), apoyándose en la base de datos del programa California Cooperative Oceanic Fisheries Investigations (CalCOFI). Estos modelos empíricos permiten determinar perfiles típicos de [Chl] para diferentes regiones biogeoquímicas y temporadas utilizando [Chl] de datos satelitales como datos de entrada.
Un modelo para estimar la producción primaria a partir de sensores remotos requiere considerar al menos los siguientes siete puntos (Platt et al. 1991): (1) la estimación de la luz disponible en la superficie del mar considerando las pérdidas en la interface aire/mar; (2) la estimación de la biomasa fitoplanctónica en la superficie del mar expresada como [Chl] usando sensores remotos; (3) la regionalización del área de estudio según las diferencias en los perfiles de [Chl] y la estacionalidad; (4) la estimación del perfil de biomasa desde la superficie hasta la base de la zona eufótica; (5) el establecimiento de los parámetros de fotosíntesis-luz, αB (pendiente inicial de la curva fotosíntesis-luz) y PmB (número de asimilación); (6) la estimación del coeficiente de atenuación de la luz (Kd); y (7) la estimación de la PP en la columna de agua.
Longhurst et al. (1995) dividieron los océanos del mundo en 57 provincias biogeoquímicas y, mediante imágenes del CZCS, estimaron la PP global en 45-50 Gt C año-1 con el modelo mencionado arriba. Dada la naturaleza de sus estimaciones, la escala espacial que definieron se basa en provincias bastante grandes, dos de las cuales incluyen el SCC. En un esquema local en el SCC, Millán-Núñez et al. (1997) dividieron las grandes provincias de Longhurst et al. (1995) en regiones y subregiones según las diferencias en los perfiles de [Chl].
]]> El objetivo del presente trabajo consistió en estimar la variabilidad espacial y temporal de la PP en la zona eufótica del SCC utilizando las concentraciones de pigmentos obtenidas de imágenes del CZCS.
Área de estudio
El área de estudio incluye el SCC del sur de California y Baja California, desde la costa hasta 122°00' W. El límite norte se encuentra entre Punta Concepción (EUA) y 34°30' N, y el límite sur entre Punta Eugenia (México) y 20°45' N (fig. 1).
En el SCC se han realizado muestreos oceanográficos periódicos (1951 hasta la fecha) desde que se inició el programa CalCOFI en 1949, los cuales han generado las series de tiempo oceanográficas más completas y extensas del mundo. Lynn y Simpson (1987) analizaron esta base de datos y definieron tres dominios: el oceánico, el costero, y una zona de transición intermedia centrada de 200 a 300 km mar adentro, paralela a la costa y coincidiendo con el núcleo del SCC.
La Cuenca del Sur de California, desde Punta Concepción hasta aproximadamente la frontera entre Estados Unidos y México, es una región costera relativamente productiva, delimitada al norte y este por la línea de costa de Norteamérica y al oeste por la Corriente de California. Es una región con una topografía de fondo compleja, con islas y bancos a partir del talud continental hacia la costa y con un patrón de circulación ligeramente relacionado con el flujo principal de la Corriente de California (Smith et al. 1988). El Frente Ensenada, una región frontal que separa las aguas eutróficas (norte) y oligotróficas (sur), es una estructura persistente a lo largo del año (Santamaría-del-Ángel et al. 2002), y puede considerarse el límite sur de la Cuenca del Sur de California. Este frente es detectable durante la mayor parte del año, pero es fuerte desde finales de marzo hasta principios de junio y muestra un desplazamiento latitudinal de alrededor de 150 km a lo largo de todo el año (Haury et al. 1993, Santamaría-del-Ángel et al. 2002).
La costa de Baja California, desde la frontera entre México y Estados Unidos hasta Punta Eugenia, está más expuesta y es influenciada por el SCC. Las condiciones eutróficas generalmente se limitan a una franja costera asociada con los eventos de surgencia, especialmente durante los primeros meses del año (invierno y primavera) en la parte sur, y durante fines de primavera y verano en la parte norte (Millán-Núñez y LaraLara 1995).
]]> Material y métodos
Se estimó la PP del fitoplancton mediante la ecuación de Platt et al. (1988):

donde P(z) es la tasa de PP a una profundidad Z (mg C nr3 Ir1), B(z) es [Chl] a una profundidad Z (mg Chl nr3), αB es la pendiente inicial de la curva P-E (mg C (mg Chl)-1 h-1 (umol quanta m-2 s-1)-1), E(z) es la irradiancia a una profundidad Z (umol quanta m-2 s-1) y PmB es el número de asimilación (mg C (mg Chl)-1 h-1).
La concentración de clorofila en la superficie ([Chl]s) se obtuvo de imágenes del color de mar del CZCS disponibles de enero de 1979 a junio de 1986. Se procesaron estas imágenes usando la corrección atmósferica y los algoritmos bioópticos de Gordon et al. (1983, 1988). Se crearon composiciones mensuales con una resolución espacial de 4 km.
A fin de calcular la PP para toda la zona eufótica a partir de los datos del CZCS, obtenidos sólo de la primera profundidad óptica, se generaron perfiles de [Chl] a partir de [Chl]s usando los parámetros gaussianos obtenidos con las ecuaciones de Millán-Núñez et al. (1997).
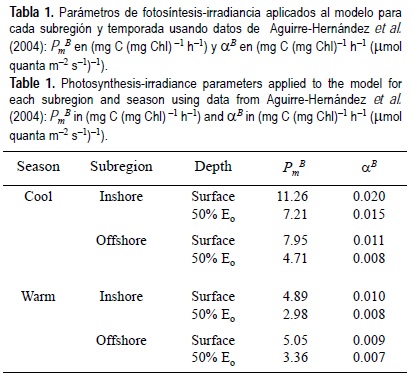
Se utilizaron diferentes parámetros de P-E de las subregiones costeras y mar adentro para cada temporada, de acuerdo con los valores publicados por Aguirre-Hernández et al. (2004) para el área de estudio; sin embargo, sus valores corresponden sólo al nivel de 50% de Eo. Debido a la falta de valores superficiales de PmB y αB, se emplearon los promedios máximos para 50% de Eo como valores superficiales, decreciendo linealmente hasta este nivel (tabla 1). Los valores se mantuvieron constantes desde la profundidad de 50% de Eo hasta la base de la zona eufótica.
El área de estudio abarcó dos regiones, el sur de California (SC) y Baja California (BC), y éstas a su vez se dividieron en dos subregiones, la costera (SCi, BCi) y mar adentro (SCo, BCo) (fig. 1). Además, se clasificaron los datos de cada región y subregión por temporada fría y cálida, como explican Millán-Núñez et al. (1997). La temporada fría en SC es de enero a mayo y en BC, de enero a junio. La temporada cálida comprendió los demás meses. Según estas divisiones, y utilizando composiciones mensuales del CZCS, se obtuvieron dos juegos de composiciones estacionales, uno para SC (fig. 2a) y otro para BC (fig. 2b).
]]> Las concentraciones de pigmentos se clasificaron en siete categorías (Millán-Núñez et al. 1997), correspondiendo a siete diferentes intervalos de [Chl]s (fig. 2). Se calculó la [Chl]s media para cada categoría en cada composición estacional para cada subregión. Subsecuentemente se determinó el perfil vertical medio para cada categoría de [Chl]s empleando las ecuaciones propuestas por Millán-Núñez et al. (1997).La irradiancia total incidente diaria (W m-2) y la duración del día fueron calculadas con la paquetería desarrollada por Platt y Sathyendranath (1995). Se calcularon la irradiancia media incidente en la superficie del mar (Eo, μmol quanta m-2 s-1) y la duración media de la luz del día (horas) para cada composición y subregión (SCi y SCo, o BCi y BCo), escogiendo un punto cerca del centro de cada subregión como representativo para estos cálculos (tabla 2).
A fin de establecer la profundidad de la zona eufótica para cada categoría de [Chl]s, se empleó la relación entre el coeficiente de atenuación (Kd) y [Chl]s, que explica 89% de la variabilidad de Kd:
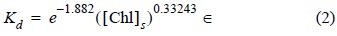
donde ∈ corresponde a los componentes que no son clorofila que contribuyen al Kd y que no fueron considerados. Esta ecuación fue establecida tras analizar la base de datos del programa CalCOFI (1979-1992) en la profundidad del disco de Secchi (ZSD) (Millán-Núñez 1996).
Se calculó una Kd típica para cada categoría de [Chl]s (ecuación 2) usando la [Chl]s media estacional de las imágenes compuestas del CZCS (fig. 2). La base de la zona eufótica se estableció en 1% y 0.1% de la irradiancia justo debajo de la superficie (Eo).
Para cada composición estacional se calcularon la PP integrada (mg C m-2 h-1) y el área (km2) cubierta por cada categoría de [Chl]s para cada subregión (SCo, SCi, BCo y BCi), a fin de estimar la PP total anual (mg C m-2 año-1) y estacional (mg C m-2 temporada-1) de cada subregión.
Resultados y discusión
La tendencia general de las imágenes del CZCS mostró valores de [Chl]s mayores en las subregiones costeras que en las subregiones mar adentro (fig. 2). El intervalo de los promedios de [Chl]s para las subregiones costeras (SCi y BCi) fue de 0.42 a 3.06 mg m-3 y para las de mar adentro (SCo y BCo) fue de 0.08 a 0.42 mg m-3. El promedio mínimo de [Chl]s para todas las subregiones se presentó durante la temporada cálida de 1983, mientras que los máximos se presentaron en 1981 y 1986 para las subregiones de SC y BC, respectivamente. La disminución de [Chl]s durante 1983 es consecuencia de la influencia de uno de los eventos ENSO más fuertes del siglo (Trenberth y Stepaniak 2001), el cual se pudo observer inclusive durante la temporada cálida de 1984.
]]> En BC la temporada fría de 1986 mostró un área ampliada de aguas eutróficas (fig. 2). En particular durante junio de 1986, las concentraciones de pigmentos mayores que 1 mg m-3 cubrieron una zona más grande de las subregiones costeras. Este valor concuerda con lo observado por González-Morales y Gaxiola-Castro (1991), quienes realizaron experimentos sobre PP en junio y julio de 1986 justo afuera de la Bahía de Todos Santos (Ensenada, México) y encontraron una [Chl] integrada muy alta (>110 mg m-2) durante los últimos días de junio. Estos autores atribuyeron estos valores altos a eventos sucesivos de surgencia moderada a intensiva, los cuales fueron evidentes en sus datos de temperatura y nutrientes, y observaron que la máxima [Chl] integrada se presentó durante el periodo de relajación de la surgencia. Aunque ellos encontraron la MPC a la profundidad de 50% de Eo, nuestra imagen compuesta del CZCS para la temporada fría de 1986 muestra florecimientos masivos de fitoplancton ([Chl]s > 2 mg m-3) en la capa superficial para toda la costa de BC (fig. 2).Los perfiles de [Chl] ajustados con el model gaussiano fueron significativamente diferentes cuando se ajustaron las integraciones a 1% ó 0.1% de Eo (prueba de bondad de ajuste, α = 0.05 y g.l. = 390); sin embargo, cuando se compararon sus valores integrados de PP correspondientes no se observó ninguna diferencia significativa (prueba de bondad de ajuste, α = 0.05 y g.l. = 390), por lo que sólo se presentarán los resultados para la integración de 1%. Varios autores (Venrick et al. 1973, Millán-Núñez et al. 1997) han encontrado casos aislados donde el fitoplancton entre 1% y 0.1% de Eo contribuye hasta 10% a la PP integrada, pero al trabajar con promedios de áreas extensas del océano, como en nuestro caso, los extremos de estos casos aislados son modulados por los valores predominantes.
Los valores diarios estimados de PP para toda el área de estudio fueron entre 0.16 y 2.63 g C m-2. En BC los valores fueron mayores en la subregión costera que en mar abierto en ambas temporadas. En SC se presentó un patrón similar en la temporada fría, pero en la temporada cálida se observó lo contrario (fig. 3). Este patrón durante la temporada cálida puede explicarse por: (a) una mayor [Chl]s en la subregión de mar abierto en SC que en BC (fig. 2), (b) un MPC más somero en SC que en BC (Millán-Núñez et al. 1997), y (c) la diferencia estacional entre los parámetros de P-E aplicados.
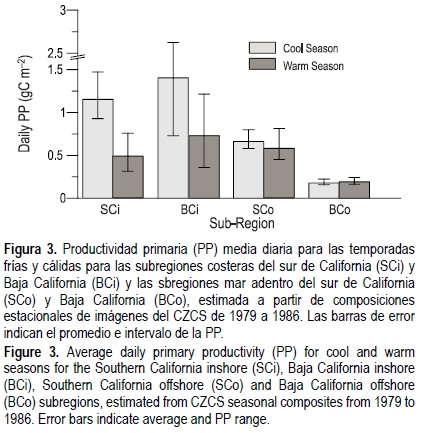
En cada subregión, los promedios de PP fueron mayores durante la temporada fría que durante la cálida, excepto en mar abierto en BC (fig. 3). Los valores más altos se presentaron en la subregión costera de BC. Este patrón de una PP alta en la temporada fría coincide con mayores [Chl] y las surgencias costeras durante los primeros meses del año y a finales de la primavera (Millán-Núñez et al. 1996), una condición persistente también registrada por Kahru y Mitchell (2001).
En BC los valores diarios de PP fueron considerablemente más bajos mar adentro que en la subregión costera, con diferencias pequeñas entre la temporada fría y la cálida y un intervalo restringido de 0.16 a 0.24 g C m-2 (fig. 3). En SC también se observó poca diferencia entre ambas temporadas, pero los valores diarios de PP fueron mayores (0.45-0.81 g C m-2). Este patrón se debe principalmente a los parámetros de P-E usados (tabla 1), que fueron mayores en la temporada fría que en la cálida pero con menor diferencia entre sí que en las sub-regiones costeras.
Otro factor a considerar en las subregiones mar adentro es la presencia y frecuencia de remolinos de mesoescala u otras estructuras que presentan una [Chl]s alta. Estudios anteriores (Simpson y Koblinsky 1986, Haury et al. 1986, Peláez et al. 1986, Jerónimo y Gómez-Valdés 2005) encontraron remolinos de mesoescala recurrentes frente a Punta Concepción y a Punta Eugenia, y observaron agua de la Corriente Subsuperficial de California en sus núcleos (Peláez et al. 1986, Simpson y Lynn 1990). Estos remolinos de mesoescala muy probablemente se relacionan con la estacionalidad de esta corriente subsuperficial y fertilizan, hasta cierto punto, las subregiones mar adentro debido a la advección vertical asociada con ellos (Simpson y Lynn 1990). En las imágenes diarias (no presentadas) se encontró una mayor frecuencia de estos remolinos de mesoes-cala o filamentos en las subregiones mar adentro durante las temporadas cálidas. Se observó que se forman dentro de las subregiones costeras o cerca del límite entre ambos dominios y que luego viajan mar adentro, moviendo aguas con una [Chl]s alta en esa dirección. Esto contribuye a una mayor [Chl] media integrada durante la temporada cálida que durante la fría en la subregión de mar adentro, especialmente en BC.
En la figura 4 se muestra la PP diaria (g C m-2), de cada año y temporada, para las subregiones costeras (a) y mar adentro (b). En las subregiones costeras, la PP en SC y BC fue mayor durante la temporada fría que durante la cálida, con excepción de 1984 en BC, probablemente debido al ENSO. Para la subregión mar adentro se encontró el mismo patrón en SC, con excepción de 1981, mientras que en BC ésta presentó menor variación entre temporadas y una PP media considerablemente menor que la de SC sin ningún patrón estacional claro. Como ya se mencionó anteriormente, esta diferencia puede estar relacionada con la diferencia en [Chl]s, la profundidad de la MPC y la variabilidad estacional de los parámetros de P-E.
]]>
Kahru y Mitchell (2000) observaron que durante el evento ENSO de 1997, las condiciones mesotróficas incrementaron mar adentro en BC de alrededor de 100 a 700 km probablemente debido a los florecimientos de cianobacterias fijadoras de nitrógeno. A fin de comparar ambos periodos de ENSO, se agruparon las categorías de [Chl]s según Kahru y Mitchell (2000) en aguas oligotróficas ([Chl]s = 0.2 mg m-3), mesotróficas (0.2 < [Chl]s =1.0 mg m-3) y eutróficas ([Chl]s > 1.0 mg m-3). Se calculó la contribución anual de cada categoría de [Chl]s a la PP para las subregiones costeras (fig. 5) y mar adentro (fig. 6). Nuestros resultados concuerdan con esta observación, presentándose un incremento de las condiciones mesotróficas en mar abierto durante la temporada cálida de 1981 en SC, mientras que en BC este incremento de las condiciones mesotróficas ocurrió durante la temporada cálida de 1982 (fig. 6). El efecto del ENSO se observó claramente en SC durante 1983, mientras que en BC éste podía observarse hasta en 1984 (fig. 6), enfatizando las diferencias entre estas dos subregiones.
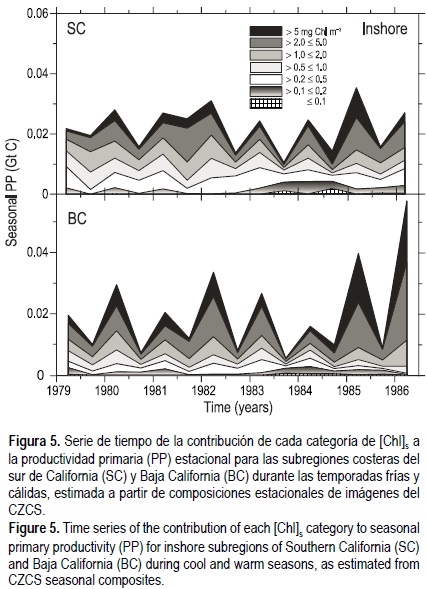

En las subregiones costeras, las aguas mesotróficas y eutróficas dominaron durante todo el periodo, excepto desde la temporada cálida de 1983 hasta la temporada fría de 1984 (fig. 5), cuando se registró una reducción de las surgencias (Kahru y Mitchell 2000, Santamaría-del-Ángel et al. 2002) y se incrementó la contribución relativa de las aguas oligotróficas a la PP cerca de la costa (fig. 5) debido a la falta de aguas de surgencia frías y ricas en nutrientes.
Aparentemente las variaciones interanuales para las sub-regiones costeras están relacionadas con los factores físicos y climatológicos que afectan las surgencias, mientras que las variaciones interanuales mar adentro son mínimas, asociadas con la circulación oceánica principal y las condiciones climatológicas y de circulación favorables para estructuras de [Chl] que se alejan del continente, presentando un patrón estacional. Es importante tomar en cuenta, sin embargo, el hecho de que el algoritmo utilizado para estimar [Chl]s es susceptible a sesgos. Kahru y Mitchell (2000) encontraron que las imágenes del CZCS tienden a sobreestimar para concentraciones altas y a subestimar para concentraciones bajas, en comparación con los datos de SeaWifs. Al comparar la PP estimada mediante imágenes del CZCS con los datos de CalCOFI tomados in situ y ordenados temporal y espacialmente de la misma forma que las imágenes, se encontró una subestimación en todos los casos, excepto para la subregión mar adentro de SC, donde las estimaciones de PP fueron mayores que las mediciones integradas de CalCOFI.
A fin de detectar las diferencias en el patrón de distribución de las subregiones costeras vs. mar adentro, se estimaron los valores anuales de PP para ambas subregiones (fig. 7). Como ya se indicó, los valores fueron mayores en las subregiones costeras que mar adentro. La subregión mar adentro presentó menor variabilidad y una PP más alta en 1981 (antes del ENSO) y más baja en 1984 (al final del ENSO) (fig. 7b). El Índice Multivariado ENSO (Wolter y Timlin 1998) evalúa la magnitud relativa de los eventos de El Niño y La Niña con base en seis variables del Pacífico tropical. Este índice se graficó junto con los resultados de la PP (fig. 7a) para el periodo de estudio y mostró evidencia del evento de 1983. El efecto del ENSO es evidente en ambas subregiones y aparece como un decremento en los valores de PP especialmente durante 1983 y 1984.
]]>
Para 1986 sólo se tienen datos para la temporada fría y, como tan sólo en ella se alcanzó el 85% de la PP para todo 1985 (fig. 7b), se pudo inferir que 1986 fue el año más productivo.
Es difícil llevar a cabo una comparación rigurosa entre las series de tiempo de PP derivadas por satélite y las mediciones in situ debido a las muy diversas características temporales y espaciales de ambas (Kahru y Mitchell 2002); sin embargo, para contar con alguna referencia, comparamos nuestras estimaciones con las de experimentos de PP de CalCOFI disponibles para el periodo de 1979 a 1986, las cuales fueron anteriormente clasificadas de acuerdo con las siete categorías de [Chl]s, las subregiones y las temporadas utilizadas en nuestro modelo. El PP total que estimamos para toda el área de estudio y todo el periodo estuvo 23.78% por debajo de los datos de CalCOFI; las regiones costeras y mar adentro estuvieron en promedio 51.82% y 5.93%, respectivamente, por debajo de los datos de CalCOFI, diferencias que se encuentran dentro de las esperadas entre métodos de medición in situ y de modelación (Platt et al. 1988, Kahru y Mitchell 2002).
Se realizó otra comparación con un modelo a escala global (Longhurst et al. 1995) y nuestra estimación diaria media de 0.80 g C m-2 para las subregiones costeras fue 16% menor y la de 0.61 g C m-2 para las subregiones mar adentro resultó 20% mayor. En esta comparación se tiene que considerar que utilizamos divisiones espaciales más detalladas que las de Longhurst et al. (1995) y que el modelo se alimentó con parámetros locales (perfil de [Chl] y parámetros P-E).
Los parámetros fotosintéticos empleados en este estudio para el modelo fueron tomados de experimentos realizados en 50% de Eo durante dos cruceros (julio de 1998 y octubre de 1998) en la zona del programa IMECOCAL (Aguirre-Hernández et al. 2004); sin embargo, se sabe que estos parámetros (PmB y αB) pueden presentar una gran variabilidad espacial y temporal (Banse y Postel 2003, Aguirre-Hernández et al. 2004), lo que hace difícil establecer un promedio para toda la columna de agua y para áreas muy grandes. Ciertamente una contribución substancial para lograr mejores estimaciones de la PP será el uso de parámetros medios de P-E más precisos para cada región y subregión y variarlos en la columna de agua, tal como fue hecho con los perfiles de clorofila (Millán-Núñez et al. 1996), siendo esto una de las bases de la determinación de provincias biogeoquímicas (Platt et al. 1991, Longhurst et al. 1995). De hecho, Banse y Postel (2003) observaron que, en la estimación de la PP mediante imágenes de satélite, el incorporar datos de campo al modelo en vez de valores promediados regional o estacionalmente, da lugar a una correlación mayor al comparar con datos in situ.
El uso de datos del CZCS puede ser materia de discusión en vista de que este sensor dejó de funcionar hace varios años. No obstante, Antoine et al. (2003) recientemente presentaron un resumen del reprocesado de estos datos usando algoritmos mejorados y enfatizaron la importancia del archivo del CZCS como referencia para el estado bioóptico del océano en la década de los ochenta, como fuente para la evaluación de las tendencias a largo plazo del color del océano.
En resumen, la PP media diaria en SC para las subregiones costeras y mar adentro fue de 0.80 y 0.61 g C m-2, respectivamente, para SC, y de 1.03 y 0.18 g C m-2, respectivamente, para BC. Los promedios diarios para todo el periodo de estudio fueron 0.29 g C m-2 para las subregiones mar adentro y 0.90 g C m-2 para las subregiones costeras. La producción total en la zona eufótica del área de estudio de enero de 1979 a junio de 1986 fue de 2.39 Gt C, lo que corresponde a una PP media anual de 0.32 Gt C.
Agradecimientos
Se agradece el apoyo brindado por el CONACYT al primer autor durante sus estudios de maestría. Agradecemos a R Hernández-Walls sus sugerencias y ayuda durante el procesamiento de los datos. De manera especial se agradecen a los revisores sus comentarios y sugerencias que hicieron que mejorar sustancialmente este escrito.
]]> Traducido al español por Christine Harris.
References
Aguirre-Hernández E, Gaxiola-Castro G, Nájera-Martínez S, Baumgartner T, Kahru M, Mitchell BG. 2004. Phytoplankton absorption, photosynthetic parameters, and primary production off Baja California: Summer and autumn 1998. Deep-Sea Res. II 51: 799-816. [ Links ]
Antoine A, Morel A, Gentili B. 2003. In search of long-time trends in ocean color. EOS 84: 301-309. [ Links ]
Banse K, Postel JR. 2003. On using pigment-normalized, light-saturated carbon uptake with satellite-derived pigment for estimating column photosynthesis. Global Biogeochem. Cycles 17: 1-11. [ Links ]
Behrenfeld MJ, Falkowski PG. 1997. Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol. Oceanogr. 42: 1-20. [ Links ]
Cullen JJ, Eppley RW. 1981. Chlorophyll maximum layers of the Southern California Bight and possible mechanisms of their formation and maintenance. Oceanol. Acta 4: 23-32. [ Links ]
González-Morales TA, Gaxiola-Castro G. 1991. Daily variation of physico-chemical characteristics, biomass and phytoplankton primary production in an upwelling coastal zone of Baja California. Cienc. Mar. 17(3): 21-37. [ Links ]
Gordon HR, Clark DK, Brown JW, Brown OB, Evans RH, Broenkow WW. 1983. Phytoplankton pigment concentrations in the Middle Atlantic Bight: Comparison of ship determinations and CZCS estimates. Appl. Opt. 22: 20-36. [ Links ]
Gordon HR, Brown OB, Evans RH, Brown JW, Smith RC, Baker KS, Clark DK. 1988. A semianalytic radiance model of ocean color. J. Geophys. Res. 93: 10909-10924. [ Links ]
Haury LR, Simpson JJ, Peláez J, Koblinsky CJ, Wiesenhahn D. 1986. Biological consequences of a recurrent eddy off Point Conception, California. J. Geophys. Res. 91: 12937-12956. [ Links ]
Haury LR, Venrick E, Fey CL, McGowan JA, Neil PP. 1993. The Ensenada front: July 1985. Cal. Coop. Oceanic Fish. Invest. Rep. 34: 69-88. [ Links ]
Jerónimo G, Gómez-Valdés J. 2005. Mean temperatura and salinity along an isopycnal surface in the upper ocean off Baja California. Cienc. Mar. 32: 663-671. [ Links ]
Kahru M, Mitchell G. 2000. Influence of the 1997-1998 El Niño on the surface chlorophyll in the California Current. Geophys. Res. Lett. 27: 2937-2940. [ Links ]
Kahru M, Mitchell G. 2001. Seasonal and nonseasonal variability of satellite-derived chlorophyll and colored dissolved organic matter concentrations in the California Current. J. Geophys. Res. 160: 2517-2529. [ Links ]
Kahru M, Mitchell G. 2002. Influence of the El Niño-La Niña cycle on satellite-derived primary production in the California Current. Geophys. Res. Lett. 29: 1-1. [ Links ]
Kirk JO. 1994. Light and Photosynthesis in Aquatic Ecosystems. Cambridge Univ. Press, Cambridge, 401 pp. [ Links ]
Longhurst A, Sathyendranath S, Platt T, Caverhill C. 1995. An estimate of global primary production in the ocean from satellite radiometer data. J. Plankton Res. 17: 1245-1271. [ Links ]
Lynn RJ, Simpson JJ. 1987. The California Current System: The seasonal variability of its physical characteristics. J. Geophys. Res. 92: 12947-12966. [ Links ]
Millán-Núñez R. 1996. Caracterización de los parámetros que determinan el perfil vertical de clorofila a en la Corriente de California. Tesis doctoral, CICESE, Ensenada, México, pp. 99. [ Links ]
Millán-Núñez R, Lara-Lara JR. 1995. Productividad primaria del fitoplancton del Pacífico mexicano: Un diagnóstico. En: González-Farías F, de la Rosa-Vélez J. (eds.), Temas de Oceanografía Biológica en México II. Universidad Autónoma de Baja California, Ensenada, México, pp. 31-60. [ Links ]
Millán-Núñez R, Álvarez-Borrego S, Trees CC. 1996. Relationship between chlorophyll maximum and surface chlorophyll concentration in the California Current System. Cal. Coop. Oceanic Fish. Invest. Rep. 37: 241-250. [ Links ]
Millán-Núñez R, Álvarez-Borrego S, Trees CC. 1997. Modeling the vertical distribution of chlorophyll in the California Current System. J. Geophys. Res. 102: 8587-8595. [ Links ]
Morel A. 1991. Light and marine photosynthesis: A spectral model with geochemical and climatological implications. Prog. Oceanogr. 26: 263-306. [ Links ]
Morel A, Babin M, Dandonneau Y. 1996. Measured and modeled primary production in the northeast Atlantic (EUMELI JGOFS program): The impact of natural variations in photosynthetic parameters on model predictive skill. Deep-Sea Res. I 43: 1273-1304. [ Links ]
Peláez J, Haury LR, Wiesenhahn D. 1986. Temperature-plant pigment-optical relations in a recurrent offshore mesoscale eddy near Point Conception, California. J. Geophys. Res. 91: 12919-12936. [ Links ]
Platt T, Sathyendranath S. 1988. Oceanic primary production: Estimation by remote sensing at local and regional scales. Science 241: 1613-1620. [ Links ]
Platt T, Sathyendranath S. 1995. Software for use in calculation of primary production in the oceanic water column. http://www.ioccg.org/software/Ocean_Production. [ Links ]
Platt T, Sathyendranath S, Caverhill CM, Lewis MR. 1988. Oceanic primary production and available light: Further algorithms for remote sensing. Deep-Sea Res. 35: 855-879. [ Links ]
Platt T, Caverhill C, Sathyendranath S. 1991. Basin-scale estimates of oceanic primary production by remote sensing: The North Atlantic. J. Geophys. Res. 96: 15147-15159. [ Links ]
Santamaría-del-Ángel E, Millán-Núñez R, González-Silvera A, Müller-Karger F. 2002. The color signature of the Ensenada Front and its seasonal and interannual variability. Cal. Coop. Oceanic Fish. Invest. Rep. 43: 156-161. [ Links ]
Sarmiento JL, Bender M. 1994. Carbon biogeochemistry and climate change. Photosynthesis Res. 39: 209-234. [ Links ]
Simpson JJ, Koblinsky CJ. 1986. Temperature-plant pigment-optical relations in a recurrent offshore mesoscale eddy near Point Conception, California. J. Geophys. Res. 91: 12919-12936. [ Links ]
Simpson JJ, Lynn RJ. 1990. A mesoscale eddy dipole in the offshore California Current. J. Geophys. Res. 95: 13009-13022. [ Links ]
Smith R, Zhang X, Michaelsen J. 1988. Variability of pigment biomass in the California Current system as determined by satellite imagery 1. Spatial variability. J. Geophys. Res. 93: 10863-10882. [ Links ]
Trenberth KE, Stepaniak DP. 2001. Indices of El Niño evolution. J. Climate 14: 1697-1701. [ Links ]
Venrick EL, McGowan JA, Mantyla AW. 1973. Deep maxima of photosynthetic chlorophyll in the Pacific Ocean. Fish. Bull. 71: 41-52. [ Links ]
Wolter K, Timlin MS. 1998. Measuring the strength of ENSO events: How does 1998/1997 rank? Weather 53: 315-324. [ Links ]
]]>