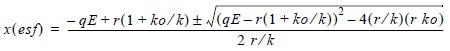
Comparación entre el uso de dos derechos de acceso pesquero, concesiones y permisos, en la pesquería de erizo rojo de mar, Strongylocentrotus franciscanus (Agassiz), en Santo Tomás, Baja California, México
Modelling the use of two fishery access rights, concessions and licences, in the red sea urchin, Strongylocentrotus franciscanus (Agassiz), fishery at Santo Tomás, Baja California, Mexico
Evlin Ramírez-Félix1*, Héctor G. Manzo-Monroy2
1 Dirección General de Investigación Pesquera en el Pacífico Norte Instituto Nacional de la Pesca Apartado postal 1177 Mazatlán, CP 82010, Sinaloa, México. *E-mail: evlinr@yahoo.com.mx
2 Facultad de Ciencias Marinas Universidad Autónoma de Baja California Apartado postal 453 Ensenada, CP 22800, Baja California, México.
]]> Recibido en junio de 2003;
Resumen
Se simularon diversos escenarios pesqueros con situaciones de riesgo para la pesquería del erizo rojo de mar, Strongylocentrotus franciscanus, y se evaluó el efecto de dos tipos de derechos de uso en la legislación pesquera mexicana: permisos y concesiones. La biomasa del recurso se calculó con dos modelos, el de depensación crítica y el de Schaefer, tanto dinámica como estáticamente. Los parámetros requeridos por cada modelo se determinaron por dos vías: información de oficinas gubernamentales y generando números estocásticos por el Método Monte Carlo ajustándose a un modelo dinámico. El ajuste de los valores de los parámetros de ambos modelos fue por medio de generación de números estocásticos por el méodo de Monte Carlo. En general, ambos modelos pronosticaron capturas menores a las obsevadas. En las simulaciones hechas para cinco años con ambos modelos para el sistema de permisos la biomasa disminuye y con concesiones se mantiene al alza con excepción de los años con temperaturas altas, por lo que la mejor opción para preservar la pesquería es otorgar concesiones en comparación con permisos.
Palabras clave: erizo rojo de mar, efecto Allee, depensación crítica, Schaefer, Bayes, concesión, permiso.
Abstract
Various stages of the red sea urchin, Strongylocentrotus franciscanus (Agassiz), fishery were simulated to evaluate the effect of two types of access rights under Mexican law: concessions and licences. The resource biomass was evaluated using the critical depensation and Schaefer models. Each model was evaluated as a static or dynamic system. The parameters required for each model were determined from government data and by generating stochastic numbers using the Monte Carlo method to adjust to a dynamic model. In general, both models predicted catches smaller than those observed. The five-year-period simulation for both models showed a drop in biomass when licences are issued, whereas a similar simulation for concessions resulted in increased biomass, except when high temperatures occurred. Our results suggest that issuing concessions rather than licences is the best option to conserve the fishery.
Key words: red sea urchin, Allee effect, critical depensation, Schaefer, Bayes, concessions, licences.
]]>Introducción
El erizo rojo de mar, Strongylocentrotus franciscanus (Agassiz), se distribuye geográficamente desde Alaska, EUA, hasta Isla Cedros, Baja California (BC), México (Mottet, 1976). Su extracción en Baja California inició en la década de 1960 (Malagrino-Lumare, 1972). A partir de los años setenta se otorgaron permisos para su explotación comercial y, hacia fines de 1994, aparecieron publicadas en el Diario Oficial de la Federación (DOF, 23 de diciembre de 1994) concesiones pesqueras del recurso. Las capturas (en peso entero) de erizo en Baja California han presentado grandes fluctuaciones con máximos en 1979 (5700 t) y 1986 (8500 t), y capturas mínimas en 1981 y 1983 (de 986 y 929 t respectivamente). De 1987 a 2001, de acuerdo a la Carta Nacional Pesquera, la producción ha oscilado entre 1000 y 5500 t (DOF, 15 de marzo, 2004). A partir de 1993 se estableció la época de veda en Baja California del 1ro. de marzo al 30 de junio de cada año (DOF, 18 de octubre de 1993).
Los equinodermos son animales exclusivamente marinos. Todos los miembros de este filo son dióicos y su fecundación es externa (Barnes, 1977). Comúnmente se localizan en mantos de macroalgas (Macrocystis pyrifera) por ser éstas su alimento preferido, aunque también pueden habitar sustratos rocosos (Palleiro, 1987) a profundidades entre diez a treinta metros (o mayores) (Mottet, 1976) creciendo hasta un diámetro de testa cercano a 18 cm (Palleiro, 1982).
Las medidas administrativas comúnmente empleadas en la pesquería del erizo en algunos países como México y Estados Unidos incluyen límite de talla, cierres temporales/área y acceso limitado (Haaker, 1992; Halman y Rudie, 1992; Botsford et al., 1993; Botsford y Quinn, 1994), pero estas varían de país a país. Con la finalidad de promover la conservación de las poblaciones de invertebrados, los administradores han definido como componentes de su conservación los siguientes: protección del potencial reproductivo, control del esfuerzo e incremento del reclutamiento y sobrevivencia (Jamieson y Caddy, 1986).
Existen dos tipos de derechos de uso comercial de recursos pesqueros en la legislación mexicana: permisos y concesiones (Ley de Pesca y su Reglamento, 1999). La diferencia entre permisos y concesiones es la temporalidad de los derechos. La concesión requiere del solicitante cumplir con un mayor número de requistos como (1) un estudio económico, (2) la solicitud debe explicar qué se va a hacer con el recurso, (3) deberá indicar cuál será la inversión y su tasa de recuperación y (4) deberá demostrar la propiedad o disposición legal de las artes de pesca y embarcaciones (redes, barcos) requeridas. Los permisos solamente requieren los números 2 y 4 (Ley de Pesca y su Reglamento, 1999) y, mientras que éstos tienen una duración de hasta 4 años, las concesiones pueden durar hasta 50 años. Por ello, es menos frecuente que se solicite una concesión que un permiso. Las concesiones se otorgan regularmente para recursos bentónicos y en una área definida geográficamente.
Con una concesión el poseedor asegura la explotación de una zona adquieriendo cierto sentido de propiedad de la misma, lo que le brinda una mayor seguridad al invertir e incentiva en él la protección y el uso eficiente del recurso. En cambio, al caducar la vigencia del permiso de extracción su poseedor no tiene ninguna seguridad de que éste se renueve, lo que puede traer consigo un uso ineficiente o abuso del recurso. Esto se conoce también como el valor del recurso en el tiempo que, para el caso de las concesiones es a futuro (largo plazo) y para los permisos vale más en el presente (corto plazo).
El aprovechamiento del erizo rojo en México se regula con la Norma Oficial Mexicana (NOM) 007-PESC-1993, misma que establece una talla mínima de 8 cm de diámetro de caparazón, el uso de una embarcación menor de hasta 7.3 metros (24 pies) de eslora con motor fuera de borda equipada con compresor para buceo semi autónomo, y con una tripulación que no exceda de tres personas (DOF, 16 de diciembre de 1993). Además, la extracción del erizo será únicamente mediante buceo semiautónomo y los organismos deberán entregarse enteros a las plantas de procesamiento, entre otras reglas. Por su parte, la Carta Nacional Pesquera estipula que no se deberán otorgar nuevas concesiones además de disminuir el esfuerzo de pesca hasta que la pesquería muestre signos de recuperación (DOF, 15 de marzo, 2004).
Entre los modelos más sencillos usados en pesquerías se encuentran los de dinámica de biomasa, que requieren pocos parámetros y variables observadas. En los modelos de producción pesquera comúnmente se emplea una función de producción biológica de tipo compensatoria, sin embargo, para organismos bentónicos y de poca movilidad se ha propuesto la existencia de mortalidad de tipo depensatoria y el efecto Allee. Los adultos ofrecen un refugio parcial a los juveniles (Tegner y Dayton, 1981), lo cual indica que el reclutamiento es denso-dependiente con un gran asentamiento en áreas donde existe un gran número de adultos (Baker, 1973). Al dispersarse las larvas existen dos posibles causas del efecto Allee en su reclutamiento: la eficiencia de fertilización disminuye con densidades bajas de desovantes (Levitan, 1991; citado por Botsford y Quinn, 1994); y el grado de protección de nuevos reclutas bajo las espinas de los adultos también disminuye cuando éstos presentan menor densidad (Tegner y Dayton, 1977, citado por Botsford y Quinn, 1994). Ambos factores hacen a la especie muy susceptible a la sobrepesca (Botsford y Quinn, 1994).
La relación depensatoria en la función de crecimiento poblacional puede ser causada tanto por una relación depredador-presa como por una disminución en el éxito de la fertilización en densidades muy bajas de organismos adultos (efecto Allee). Estos dos mecanismos pueden actuar simultáneamente, o aún más, reforzarse. Neave (1953) fue el primero en utilizar el término depensación y propuso tres tipos de mortalidad: (1) compensatoria, cuando la población tiende a equilibrarse en un nivel de capacidad de carga; (2) depensatoria, cuando la mortalidad tiende a ser muy alta conforme la densidad de población disminuye; y (3) extrapensatoria, la mortalidad es densoindependiente.
]]> La depredación total se ve afectada por dos variables esenciales, la densidad de la presa y la del depredador. Las características principales de la depredación pueden entenderse por los cambios dinámicos de ambas variables. La respuesta funcional se define como el incremento en el número de presas consumidas por depredador, conforme la densidad de la presa se incrementa. El cambio en la densidad del depredador se denomina respuesta numérica (Solomon, 1949 citado por Holling, 1959).Ricker (1954) identificó que el proceso de mortalidad depensatoria puede resultar de que la línea de reemplazo cruce la curva de reproducción (reclutamiento) en dos o tres puntos, lo cual da origen a más de un punto de equilibrio de la población. Si la curva de crecimiento cruza la línea de reemplazo por abajo existe un equilibrio inestable y el tamaño de la población tiende a cambiar hasta otro punto de equilibrio estable, el cual se alcanza cuando la curva de crecimiento cruza la de reemplazo por arriba. Éstos son los llamados dominios de atracción de la población (Peterman, 1977).
El presente trabajo evalúa los efectos de una concesión pesquera, en comparación con los de un permiso de extracción, en la pesquería de erizo del área de Santo Tomás a Punta China, Baja California (México), y calcula la biomasa por medio de los modelos de depensación crítica y de Schaefer.
Materiales y métodos
El área de estudio comprendió de Punta Santo Tomás a Punta China, BC, México, a aproximadamente 55 km al sur del Puerto de Ensenada. Es una zona protegida de la acción de las corrientes y el oleaje, con grandes zonas con fondo arenoso pero con una franja rocosa de bajo relieve y con mantos de algas cafés, Macrocystis pyrifera (Agardh), que dependiendo de las condiciones ambientales pueden cubrir gran parte del área (Romero y Bertsch, 1984).
Los parámetros se calcularon para el período de 1983-1994, sin embargo, la información para estimarlos se obtuvo de diferentes fuentes para distintos períodos. Para el periodo de 1983-1987 se obtuvo con base en la contribución porcentual de la zona a la captura estatal, para el de 1988-1992 de Palleiro et al. (1994), y para el de 1993-1994 de la Subdelegación Federal de Pesca en Baja California. El coeficiente de capturabilidad (q) se obtuvo con los datos de biomasa determinados para la zona por Palleiro (1982).
Para describir el comportamiento de la biomasa poblacional en el tiempo y predecir las capturas con el modelo de producción de Schaefer y de depensación crítica tanto estática como dinámicamente, se determinaron los parámetros para cada uno de los modelos: tamaño de la población en equilibrio natural (k), biomasa de la población capturable (B), tasa intrínseca de crecimiento poblacional (r), coeficiente de capturabilidad (q) y población crítica (ko). El modelo de Schaefer no utiliza este último parámetro.
Para determinar los parámetros poblacionales se generaron números estocásticos por el Método Monte Carlo (Sóbol, 1983). El rango de cada uno de ellos fue r = 0.3-1.1, k = 1800-5500 t, B = 1300-3000 t, ko = 65-200 t, y q = 7x10-5-1.4 X 10-4, y por medio de un ajuste de mínimos cuadrados se determinó el valor óptimo de cada parámetro, al que se le denominó parámetro base.
Para el modelo estático de Schaefer (Hilborn y Walters, 1992):
]]> Y = q k [1- q/r E] = q k E - k q2/r E2 = a E - b E2donde Y es la captura en equilibrio ó la producción sustentable que le corresponde al esfuerzo E, k es la capacidad de carga, r la tasa intrínseca de crecimiento poblacional, q el coeficiente de capturabilidad, a el intercepto y b la pendiente. Para el modelo estático de depensación crítica (Clark, 1976) la biomasa en equilibrio estático está dada por:
Para el modelo dinámico de Schaefer (Hilborn y Walters, 1992) tenemos:
xt+1 = xt + r xt [1 - xt/k] - ct
donde ct es la captura observada en el tiempo t, xt es la biomasa en el tiempo t y xt+1 es la biomasa en el tiempo t + 1.
La captura calculada es
Ĉ t = q xt Et
donde Ĉ t es la captura calculada y Et el esfuerzo observado en el tiempo t.
Para el modelo dinámico de depensación la biomasa es
]]> xt+1 = xt + r (xt - k0) (1 - xt/k) - ctLa captura calculada en el tiempo es ĉt = q xt Et.
Para predecir la biomasa y captura con eventos riesgosos se utilizó la probabilidad de Bayes en situación de explotación pesquera con permisos y concesiones sólo con los parámetros base.
Para determinar los riesgos asociados se realizaron encuestas entre los actores involucrados en la pesquería incluyendo permisionarios, industriales, buzos y administradores. A partir de ello se definieron posibles escenarios con relación a los porcentajes de pesca furtiva, riesgo de cambio de zona al renovar permisos y ocurrencia de temperaturas anómalas (fig. 1). Sobre la cuota de captura previamente establecida se definieron dos porcentajes de pesca furtiva, 30% y 10%, y para ambos casos tres posibles intervalos de temperatura (alta, baja y normal). Con relación al comportamiento de los permisionarios se consideraron dos alternativas, que el permisionario capture 30% más de la cuota de captura establecida (P en la fig. 1) o que respete y ejerza únicamente su cuota de captura (I en la fig. 1). La combinación de estas alternativas da un total de doce posibles escenarios para cada modelo (fig. 1). De estos posibles escenarios solamente se tomaron en cuenta el 5, el más negativo y el 10, el más benigno.
Para calcular la biomasa con los modelos dinámicos en el caso de permiso de pesca se utilizaron los parámetros base con información de captura y esfuerzo de 1994 y se consideró una mortalidad por pesca del 20% de la biomasa como captura calculada y posteriormente restándola a la biomasa calculada para el año posterior. En todos los casos se calculó el riesgo con base en la biomasa para obtener la captura asociada a la misma.
En el caso de la pesca furtiva se sumaron los porcentajes de extracción ilegal (estimado de las encuestas, 30% y 10%) a la captura observada, se corrieron los nuevos valores de captura con cada modelo y se determinó la probabilidad previa para 0, 10%, 20% y 30% de pesca ilegal asociada al valor de la nueva biomasa con una prueba z, posteriormente se multiplicó por la función de verosimilitud (con los parámetros base y ambos modelos) y se dividió por la probabilidad total para obtener la probabilidad posterior de Bayes y sus capturas asociadas.
Para determinar el riesgo debido al cambio de zona se calcularon las probabilidades previas de manera similar a lo explicado anteriormente, sólo que aquí el supuesto es que, dada la incertidumbre de cambiar de área la siguiente temporada, el permisionario tiende a capturar 30% más de su cuota asignada. Estas probabilidades previas se multiplicaron por las anteriormente obtenidas.
En los casos de las temperaturas anómalas altas y bajas se consideró que estos fenómenos tuvieron una duración de dos años a partir de 1997 (Wolter y Timlin, 1997). Con respecto al efecto por variabilidad ambiental, las encuestas mencionaron que en años cálidos la calidad de la gónada de erizo disminuye porque M. pyrifera, el principal alimento del erizo, decae por una inhibición de nutrientes relacionada con temperaturas altas (Zimmerman y Robertson, 1985; Dean y Jacobsen, 1986). Se buscó una asociación entre las capturas anuales de S. franciscanus y M. pyrifera durante 1977-1993 (Molina, 1986) para Baja California, así como de éstas con la temperatura (NOAA, 1991) media anual superficial del mar de 1960-1990, con correlaciones de Pearson a un nivel de significancia del 95%.
Se determinaron los niveles de biomasa asociados con temperaturas bajas (<16°C), normales (entre 16 y 18°C) y altas (>18°C) obteniendo también sus probabilidades previas, que se multiplicaron por las dos anteriores para obtener los nuevos valores esperados de biomasa y captura con cada escenario.
]]> En el caso de la concesión solamente se consideraron los riesgos relacionados a fenómenos naturales (temperaturas), por lo que se obtuvo la probabilidad previa asociada a la biomasa en cada rango de temperatura, después la total y, con éstas y la función de verosimilitud, la probabilidad posterior (Bayes).La descripción completa de la simulación con modelos dinámicos, riesgos y parámetros base es:
1. Una vez obtenidos los parámetros base se determinaron los riesgos asociados a la pesquería.
2. Se calculó la probabilidad previa con los riesgos mencionados en las encuestas.
3. Se obtuvo la función de verosimilitud con los modelos de Schaefer y de depensación crítica con parámetros base utilizando una mortalidad de 20% de la biomasa para considerarla en la captura calculada.
4. Se multiplicó la probabilidad previa de los escenarios 5 y 10 (fig. 1) por la función de verosimilitud y se dividió entre la probabilidad total.
Para comparar los escenarios 5 y 10 (fig. 1) y los derivados de concesiones con temperaturas normal, baja y alta, se calcularon los valores esperados haciendo la sumatoria de los valores obtenidos de multiplicar la probabilidad de cada escenario por la captura esperada en el mismo (Berry, 1996) de acuerdo a la siguiente ecuación:
VE = Σ capti, X prob. capti
donde capti es la captura calculada de cada escenario y prob. capti la probabilidad de que la captura se obtenga en ese escenario particular.
]]> Resultados
El modelo estático de depensación crítica con parámetros base (fig. 2; tablas 1 y 2) presenta dos puntos de equilibrio poblacional para el mismo nivel de esfuerzo y muestra que el tamaño de la población cambia con diferentes niveles de esfuerzo hasta llegar al esfuerzo crítico de 3500 viajes de pesca alcanzando una captura sostenible cercana a 180 t. Con parámetros provenientes de la fuente oficial, tenemos un esfuerzo crítico máximo cercano a 5000 viajes de pesca con una captura de casi 270 t y 400 t de biomasa poblacional.
El modelo estático de Schaefer con los parámetros base muestra el nivel de esfuerzo en que ocurre un tamaño poblacional en equilibrio y muestra una relación inversa entre el esfuerzo pesquero y el tamaño de la población en equilibrio. En los 2500 viajes de pesca se alcanza la máxima captura sostenible de casi 400 t (fig. 2, tablas 1 y 2), mientras que con los parámetros calculados a partir de una fuente oficial se encuentra casi en los 4000 viajes de pesca con cerca de 470 t de captura.
La figura 3 muestra la biomasa calculada con el modelo de depensación crítica y, a temperaturas altas, se observa un marcado decremento en la biomasa. El comportamiento de las curvas de captura para las predicciones de biomasas anteriores son similares y menores, con un mínimo de captura esperada menor a 200 t. Comparado con el modelo de Schaefer son muy similares e iguales para el último año.
Las biomasas (fig. 4) con temperaturas normales muestran un ligero descenso con el modelo de Schaefer y son casi constantes con el de depensación. Sus correspondientes capturas calculadas oscilan entre 400 y 500 t para ambos modelos.
En los escenarios de concesión (fig. 5) la biomasa baja al simular una temperatura alta durante dos años con ambos modelos, y después aquella se incrementa al normalizarse ésta. Con el modelo de depensación sus capturas son menores a 400 t, pero son poco mayores con el de Schaefer. En el escenario de una concesiones pesqueras, si la temperatura es baja durante dos años, ambos modelos siempre aumentan sus biomasas (casi 3000 t) al igual que sus capturas (casi 600 t) aunque la condición se normalice. Con temperaturas normales el comportamiento es similar pero de menor magnitud.
Discusión
]]> Hace aproximadamente 10 años en México era difícil obtener una concesión, por lo que era necesario determinar si en términos de conservación del recurso ésta es una mejor opción de manejo que los permisos de extracción. Se eligieron los modelos descritos porque muchos investigadores utilizan el modelo de Schaefer para la pesquería del erizo rojo de mar y, debido al comportamiento particular de esta especie (efecto Allee) era necesario considerar una población crítica con el modelo de depensación para después comparar los resultados de ambas aproximaciones.La curva de equilibrio poblacional de los modelos estáticos de depensación crítica muestra cuánto cambiará la población a diferentes niveles de esfuerzo, mostrando el nivel de esfuerzo que producirá una captura igual a la tasa de crecimiento para varios tamaños de la población y, alternativamente, los tamaños de equilibrio poblacional que corresponderán a varios niveles de esfuerzo. Con ambos parámetros, la parte con pendiente positiva de la curva, a bajos tamaños de población, muestra una disminución de la población a la que se debe oponer otra en el esfuerzo para asegurar que el nuevo tamaño de la población esté en equilibrio. Esto no significa que un incremento en el esfuerzo aumente el tamaño de la población en equilibrio. Un incremento en el esfuerzo en un punto de equilibrio poblacional dado hará disminuir la población debido a que la captura será mayor que el crecimiento.
Las curvas de captura sostenible muestran dos capturas para cada nivel de esfuerzo menor a 3500 viajes (o 5000 viajes, dependiendo los parámetros), en donde solamente hay un equilibrio poblacional y, por lo tanto, una captura sostenible (esfuerzo crítico). Por lo mismo, para niveles de esfuerzo mayores a 3500 viajes (o a 5000 viajes) no existe población en equilibrio, y de aquí que la captura sostenible sea cero.
En la porción de la curva sobre la parte de la curva de equilibrio poblacional con pendiente negativa, al incrementar el esfuerzo decrece la curva de equilibrio poblacional de manera que la captura es mayor que el crecimiento. No se puede decir cuál de los dos modelos es mejor debido a que hay que alcanzar los límites críticos del esfuerzo para compararlos y ver cómo se ajustan. Quizá ambos son buenos en la parte casi lineal de la curva superior de biomasa del de depensación crítica antes de llegar a la inflexión como en el de Schaefer ya que son muy similares sus pendientes; probablemente el primero es mejor por tener mayores consideraciones (como la población mínima viable) ya que los erizos requieren densidades mínimas en su reclutamiento (Botsford y Quinn, 1994).
Entre los factores físicos que pueden afectar la capacidad de carga de la pesquería del erizo está la disminución de alimento disponible debida a la variabilidad de temperaturas (Tegner, 1980) y a tormentas (Dayton y Tegner, 1984). Otro efecto denso-dependiente es la presencia o ausencia de erizos adultos, ya que los jóvenes utilizan las espinas de los adultos para protegerse de los depredadores y compartir su alimento.
El nivel de esfuerzo necesario para extinguir el recurso en el área de estudio, según el modelo de Schaefer y los parámetros obtenidos de la fuente oficial, es más de la mitad del esfuerzo anual medio observado. De acuerdo con el modelo logístico y los valores de los parámetros provenientes de la fuente oficial, el nivel de esfuerzo para eliminar completamente el recurso es más de la mitad del esfuerzo medio calculado para Baja California (12000 viajes de pesca). Este esfuerzo es excesivo para un área tan pequeña en relación al área total de pesca de erizo.
Cuando se determinaron los parámetros a partir de una fuente oficial solamente se contó con cuatro años de capturas y sus esfuerzos respectivos, lo cual es un período corto de tiempo para obtener parámetros poblacionales. Otra fuente de error también puede deberse a que la extracción de macroalgas se realiza durante todo el año, mientras que la de erizo rojo sólo es durante 8 meses al año.
El cálculo de los parámetros de crecimiento y sobrevivencia de los erizos jóvenes es difícil debido a que la tasa intrínseca de crecimiento poblacional es específica para su tamaño, creciendo los pequeños más lentamente (Tegner, 1989).
Si el alga a la deriva está limitada, los erizos abandonarán su hábitat escondido y ramonearán en las algas fijas (Tegner, 1989) lo cual los hace más vulnerables a la depredación. Además, con el incremento de la temperatira del agua debido a El Niño, el erizo disminuye su tasas de alimentación (Kato y Schroeter, 1985). También, el tiempo de fase planctónica depende de la temperatura (Tegner, 1989).
Las simulaciones de riesgo se hicieron para seis años debido a que se consideró que la condición ambiental anómala de temperatura tardaría dos años en desaparecer, que las tasas de reclutamiento de los erizos son muy lentas durante y después de El Niño (Tegner, 1992) y que les toma de 4 a 5 años en alcanzar la talla comercial (Kato y Schroeter, 1985). Por otra parte, el promedio de vida de las embarcaciones menores utilizadas para su pesca según las encuestas es de siete años y de los motores de tres.
]]> Al simular una situación anómala de temperatura alta, era de esperar que el escenario de permisos arrojara las biomasas más bajas, sin embargo llama la atención que éstas no se recuperan aún después de que la condición anómala haya terminado.Al simular temperaturas bajas, el modelo de depensación después de los ligeros incrementos de biomasa por las bajas temperaturas, éstas tienden a descender y al igual que en los casos anteriores las probabilidades posteriores son menores que las que resultan del modelo de Schaefer. En el escenario de concesión solamente se incluyó lo relacionado con riesgos en los cambios de temperatura y como se esperaba, las temperaturas altas hacen que la biomasa disminuya más con el modelo de depensación crítica que con el de Schaefer y, en general, sus probabilidades previas y posteriores son ligeramente mayores para este último, pero con ambos modelos tiende a recuperarse. Las probabilidades previas con temperaturas bajas y normales fueron similares para ambos modelos y la tendencia es a elevar la biomasa.
Es muy difícil determinar las probabilidades previas de todos los escenarios de riesgo porque interviene la subjetividad, pero puede ser útil cuando se desea simular una situación difícilmente cuantificable, como es el efecto de la pesca furtiva y de los cambios de zona al renovar permisos. En este caso particular, es evidente (con ambos modelos y parámetros) que de manera general la biomasa de la población de erizo rojo se encuentra disminuyendo y esto habrá que tomarlo en consideración dependiendo de los eventos adversos que se prevea que puedan afectar cada temporada.
Desafortunadamente no existen trabajos similares para el erizo rojo de mar. Es importante recalcar que si no se toman medidas de manejo adecuadas para el recurso, debido a la presión que representa su alto valor comercial y su importancia económica para Baja California éste podría acabarse o disminuir drásticamente su captura.
Agradecimientos
Se agradece a los revisores anónimos, a los pescadores de Baja California que proporcionaron información, comentarios y gentilmente respondieron la encuesta. A Ellis Glazier por traducir el escrito. ERF agradece la beca otorgada por CONACYT.
Referencias
Baker, S. (1973). Growth of the red sea urchin Strongylocentrotus franciscanus (Agassiz) in two natural habitats. M.Sc. thesis, California State University, San Diego, 83 pp. [ Links ]
Barnes, D.R. (1977). Zoología de los Invertebrados. 3ra ed. Interamericana, México, DF, 826 pp. [ Links ]
Berry, A.D. (1996). Statistics. A Bayesian Perspective. Duxbury Press. Wadsworth Co., 518 pp. [ Links ]
Botsford, L.W. and Quinn, J.F. (1994). Spatial management of the northern California red sea urchin fishery. University of California, Davis, R/F-150, 4 pp. [ Links ]
Botsford, L.W., Quinn, J.F., Wing S.R. and Brittnacher, J.G. (1993). Rotating spatial harvest of a benthic invertebrate, the red sea urchin, Strongylocentrotus franciscanus. Proceedings of the International Symposium on Management Strategies of Exploited Fish Populations. Alaska Sea Grant College Program, AK-SG-93- 02. pp. 409-428. [ Links ]
Clark, C.W. (1976). Mathematical Bioeconomics. John Wiley, 352 pp. [ Links ]
Dayton, P.K. and Tegner, M. (1984). Catastrophic storms, El Niño, and patch stability in a southern California kelp community. Science, 224(4646): 283-285. [ Links ]
Dean, T.A. and Jacobsen, F.R. (1986). Nutrient limited growth of juvenile kelp, Macrocystis pyrifera, during the 1982-1984 "El Niño" in southern California. Mar. Biol., 90: 597-601. [ Links ]
Haaker, P. (1992). The southern California red sea urchin: A case history. The management and enhancement of sea urchins and other kelp bed resources: A Pacific rim perspective publication. T-CSGCP-028. California Sea Grant College, University of California, pp. 1-43. [ Links ]
Halman, P. and Rudie, D. (1992). The southern California red sea urchin fishery, 1972-1992. The management and enhancement of sea urchins and other kelp bed resources: A Pacific rim perspective publication. T-CSGCP-028. California Sea Grant College, University of California, pp. 1-43. [ Links ]
Hilborn, R. and Walters, C.J. (1992). Quantitative Fisheries Stock Assessment: Choice, Dynamics and Uncertainty. Chapman and Hall, New York, 570 pp. [ Links ]
Holling, C.S. (1959). The components of predation as revealed by a study of small-mammal predation of the European pine sawfly. Can. Entomol., XCI: 293-320. [ Links ]
Jamieson, G.S. and Caddy, J.F. (1986). Research advice and its application to management of invertebrate resources: An overview. In: G.S. Jamieson and N. Bourne (eds.), North Pacific Workshop on Stock Assessment and Management of Invertebrates. Can. Spec. Publ. Fish. Aquat. Sci., pp. 416-424. [ Links ]
Kato, S. and Schroeter, S.C. (1985). Biology of the red sea urchin, Strongylocentrotus franciscanus, and its fishery in California. Mar. Fish. Rev., 47(3), 1-19. [ Links ]
Ley de Pesca y su Reglamento (1999). Secretaría de Medio Ambiente, Recursos Naturales y Pesca. México, DF, 113 pp. [ Links ]
Malagrino-Lumare, G. (1972). Extracción y preservación de las gónadas de erizo. Tesis de licenciatura, Universidad Autónoma de Baja California, Ensenada, México, 79 pp. [ Links ]
Molina, M.J. (1986). Notas sobre tres especies de algas marinas: Macrocystis pyrifera, Gelidium robustum y Gigartina canaliculata de interés comercial en la costa occidental de Baja California, México. Contribuciones Biológicas y Tecnológico-Pesqueras. SEPESCA, Instituto Nacional de la Pesca. Ensenada, BC. Doc. Téc. No. 3, 39 pp. [ Links ]
Mottet, M.C. (1976). The fishery biology of the sea urchin in the family Strongylocentrotidae. Washington Dept. Fish. Tech. Rep., No. 20, 66 pp. [ Links ]
Neave, F. (1953). Principles affecting the size of pink and chum salmon populations in British Columbia. J. Fish. Res. Board Can., 9(9): 450-491. [ Links ]
NOAA (1991). CD-ROMs NODC-02 and NODC-03: Global ocean temperature and salinity profiles. US Dept. Commerce, Washington, DC, Informal Rep. No. 12. [ Links ]
Palleiro, J. (1982). Estimación de la densidad y crecimiento del erizo rojo Strongylocentrotus franciscanus (Agassiz) para la zona de Santo Tomás, Baja California, México. Tesis de licenciatura, Universidad Autónoma de Baja California, Ensenada, México, 68 pp. [ Links ]
Palleiro, J. (1987). Biología del erizo rojo Strongylocentrotus franciscanus y su pesquería en Baja California. Proyecto: Erizo de Mar. Instituto Nacional de la Pesca, Centro Regional de Investigaciones Pesqueras de Ensenada. Documento Interno, 19 pp. [ Links ]
Palleiro, J., Aguilar, D. y Navarrete, M. (1994). Cambios en la densidad de población de erizo rojo Strongylocentrotus franciscanus, en un área sujeta a explotación comercial, Santo Tomás, BC. Instituto Nacional de la Pesca, Centro Regional de Investigación Pesquera de Ensenada. Documento Interno, 17 pp. [ Links ]
Peterman, R.M. (1977). A simple mechanism that causes collapsing stability regions in exploited salmon populations. J. Fish. Res. Board Can., 34: 1130-1142. [ Links ]
Ricker, W.E. (1954). Stock and recruitment. J. Fish. Res. Board Can., 11(5): 559-623. [ Links ]
Romero, J. and Bertsch, H. (1984). Efectos de la pesquería del erizo rojo, Strongylocentrotus franciscanus (Echinordermata), sobre la estructura de su población en Santo Tomás, Baja California, México, con una comparación de métodos de muestreo. Cienc. Mar., 10(2): 85-95. [ Links ]
Sóbol, I.M. (1983). Método de Montecarlo. Ed. MIR, Moscow, 79 pp. [ Links ]
Tegner, M.J. (1980). Multispecies considerations of resource management in southern California kelp beds. Can. Tech. Rep. Fish. Aquat. Sci., No. 954: 125-143. [ Links ]
Tegner, M.J. (1989). The feasibility of enhancing red sea urchin, Strongylocentrotus franciscanus, stocks in California: An analysis of the options. Mar. Fish. Rev., 51(2): 1-22. [ Links ]
Tegner, M.J. (1992). Red urchin recruitment patterns in southern California. In: The Management and Enhancement of Sea Urchins and Other Kelp Bed Resources: A Pacific Rim Perspective. Calif. Sea Grant. Rep. No. T-CSGCP-028, 43 pp. [ Links ]
Tegner, M.J. and Dayton, P.K. (1981). Population structure, recruitment and mortality of two sea urchins: Strongylocentrotus franciscanus and S. purpuratus, in a kelp forest near San Diego, California. Mar. Ecol. Prog. Ser., 5: 255-268. [ Links ]
Wolter, K. and Timlin, M. (1997). Multivariate ENSO index for the six strongest historic El Niño events vs. the current event. NOAA-CIRES. Climate Diagnostics Center, University of Colorado at Boulder, USA. Internet (www.srh.noaa.gob/lch/research/BPTLCH.HTM). [ Links ]
Zimmerman, R.C. and Robertson, D.L. (1985). Effects of El Niño on local hydrography and growth of the giant kelp, Macrocystis pyrifera, at Santa Catalina Islands, California. Limnol. Oceanogr., 30(6): 1298-1302. [ Links ]
]]>

