
> in the beginning of XX century, was also relevant during this period, and was represented by scientists like Pavlov, Watson, Skinner, and Thorndike, who detailed a variant of learning recognized in our days as associative learning. In turn, this variant can be divided into classic conditioning (associates a stimulus with a response) and instrumental or operant conditioning (associates a stimulus with a specific behavior). Behaviorists claimed that only on observational basis of behavior is possible to know processes linked to learning. However, these authors clearly made a mistake when they stated that knowledge on processes occurring in the brain will always be far of the understanding of the investigator. Later on, one of the hardest evidences for the study of memory processes came from clinic studies of patients with focal cerebral lesions. Penfield, Scoville, and Milner, during the 60's, documented the effects of surgical lesions of temporal lobe on declarative memory, finding selective alterations, such as severe anterograde amnesia and retrograde amnesia with temporal gradient. These findings were accompanied also by a description of memory subsystems remaining intact, despite of those temporal lobe lesions (procedural memory, long-term memory, etc.). Based on these findings, medial temporal lobe was described as a key structure for the acquisition of declarative information. In parallel to clinic studies, a first attempt to coordinate psycho-psychiatric research with the knowledge and scientific protocol of biology occurred in the 60's, thus allowing the emergence of disciplines known as cognitive neuroscience and cognitive psychology or psychophysiology, both created in an effort to explore the cellular and molecular mechanisms responsible for the storage of memory information. The list of significant findings provided by these two scientific disciplines is extensive, but among the most important are probably those derived from the study of the most elementary memory processes (i.e. habituation and sensitization) in invertebrate animal models, such as Aplysia spp. These researches established the basis for the basic cellular requirements for elementary learning, as well as the molecular basis for short-and long-term memory. The conjugation of these clinical descriptions together with experimental evidence led to the postulation of the first classifications of memory systems during the 80's. One of these classifications divided memory processes in two categories: declarative memory - the one containing information consciously acquired and easy to verbalize or transmit to other persons (this type of memory is also divided in semantic and episodic memories)-, and non-declarative -including that information not easy to verbalize or which acquisition is unconscious (this type of memory implies heterogeneous information in which the role of consciousness is complex and remains in discussion, and includes procedural memory, priming, classic, and operant conditioning, as well as the most elementary forms of learning, such as habituation and sensitization). To conceptualize these memory systems in an isolated form is a mistake since is results clear enough that all of them work together most of the time, and they work independently only very seldom, some other times cooperating, or even functioning in a competitive manner. Experimental studies have shown that the role of memory systems is different each time, even when two subjects perform exactly the same learning task, thus suggesting that codification, motivation and initial handling of information may determine whether this is processed as procedural, spatial or semantic information. The recent description of competitive relationships among the different memory systems (declarative vs. procedural) resulted to be an outstanding finding, although at the present there is hard clinical and experimental evidence indicating that a decrease of functions in procedural, spatial or declarative systems may induce the activation of another memory system. Initial studies in this field suggest that transient inactivation of striatal dorsal structures (implicated in performance of motor skills) or those from the hippocampus (implicated in the performance of spatial skills) facilitate learning in that system remaining active after pharmacological challenge. The real meaning of this competition among systems still remains unclear, although it has been proposed that both systems have evolved separately in response to distinct needs, which in turn might explain why eventually these systems can compete for the handling of information. Unfortunately, several studies have demonstrated that relationships among memory systems are complicated and poorly understood until now. Semantic memory Semantic memory mainly refers to information stored on characteristics defining concepts (facts lacking a well-defined spatial/ temporal frame), as well as processes allowing its efficient recovery for further utilization in language and thought. Knowledge about the anatomic location of semantic representations has gained attention with the use of new functional neuroimaging techniques. These studies have shown that the information about the features of specific objects needed to create concepts is stored in the same neuronal systems that remain active during the perception of different stimuli. The way in which this conceptual information is organized remains unclear so far, but there is evidence suggesting that its organization proceeds on the basis of grouped categories of concepts. Pathological studies in patients suffering semantic dementia have also demonstrated that some areas, such as temporal pole and perirhinal cortex, are relevant for semantic processing.]]>
Sistemas de memoria: reseña histórica, clasificación y conceptos actuales. Primera parte: Historia, taxonomía de la memoria, sistemas de memoria de largo plazo: la memoria semántica
Memory systems: historical background, classification, and current concepts. Part one: History, taxonomy of memory, long–term memory systems: Semantic memory
Paul Carrillo–Mora1
1 Laboratorio de Aminoácidos Excitadores, Instituto Nacional de Neurología y Neurocirugía Manuel Velasco Suárez.
]]> Correspondencia:
]]> Recibido: 14 de abril de 2009.
Abstract
Memory is a fascinating brain function by means of which the nervous system can codify, store, organize, and recover a variety of information relevant to the subject. The formal study of memory started more than a century ago, providing in this time a considerable amount of scientific information on memory functioning. The actual knowledge of memory allow us to consider it far from being unique, isolated or static function, but more as a complex net of memory systems working in parallel for a common goal. Historic evolution of memory concepts and their classifications have progressed simultaneously to our knowledge on these systems.
The first empiric approaches to memory description were found in ancient Greece by authors like Plato and Aristoteles, and were based on philosophy, thinking, and introspection and logic methods. However, the first real scientific approaches appeared in the XIX century, by authors such as Ebbinghaus and Lashley, who initiated the experimental study of memory in humans and animals, respectively.
The contribution of the intellectual group known as <<behaviorist>> in the beginning of XX century, was also relevant during this period, and was represented by scientists like Pavlov, Watson, Skinner, and Thorndike, who detailed a variant of learning recognized in our days as associative learning. In turn, this variant can be divided into classic conditioning (associates a stimulus with a response) and instrumental or operant conditioning (associates a stimulus with a specific behavior). Behaviorists claimed that only on observational basis of behavior is possible to know processes linked to learning. However, these authors clearly made a mistake when they stated that knowledge on processes occurring in the brain will always be far of the understanding of the investigator.
Later on, one of the hardest evidences for the study of memory processes came from clinic studies of patients with focal cerebral lesions. Penfield, Scoville, and Milner, during the 60's, documented the effects of surgical lesions of temporal lobe on declarative memory, finding selective alterations, such as severe anterograde amnesia and retrograde amnesia with temporal gradient. These findings were accompanied also by a description of memory subsystems remaining intact, despite of those temporal lobe lesions (procedural memory, long–term memory, etc.). Based on these findings, medial temporal lobe was described as a key structure for the acquisition of declarative information. In parallel to clinic studies, a first attempt to coordinate psycho–psychiatric research with the knowledge and scientific protocol of biology occurred in the 60's, thus allowing the emergence of disciplines known as cognitive neuroscience and cognitive psychology or psychophysiology, both created in an effort to explore the cellular and molecular mechanisms responsible for the storage of memory information. The list of significant findings provided by these two scientific disciplines is extensive, but among the most important are probably those derived from the study of the most elementary memory processes (i.e. habituation and sensitization) in invertebrate animal models, such as Aplysia spp. These researches established the basis for the basic cellular requirements for elementary learning, as well as the molecular basis for short–and long–term memory.
The conjugation of these clinical descriptions together with experimental evidence led to the postulation of the first classifications of memory systems during the 80's. One of these classifications divided memory processes in two categories: declarative memory — the one containing information consciously acquired and easy to verbalize or transmit to other persons (this type of memory is also divided in semantic and episodic memories)—, and non–declarative —including that information not easy to verbalize or which acquisition is unconscious (this type of memory implies heterogeneous information in which the role of consciousness is complex and remains in discussion, and includes procedural memory, priming, classic, and operant conditioning, as well as the most elementary forms of learning, such as habituation and sensitization).
To conceptualize these memory systems in an isolated form is a mistake since is results clear enough that all of them work together most of the time, and they work independently only very seldom, some other times cooperating, or even functioning in a competitive manner. Experimental studies have shown that the role of memory systems is different each time, even when two subjects perform exactly the same learning task, thus suggesting that codification, motivation and initial handling of information may determine whether this is processed as procedural, spatial or semantic information. The recent description of competitive relationships among the different memory systems (declarative vs. procedural) resulted to be an outstanding finding, although at the present there is hard clinical and experimental evidence indicating that a decrease of functions in procedural, spatial or declarative systems may induce the activation of another memory system. Initial studies in this field suggest that transient inactivation of striatal dorsal structures (implicated in performance of motor skills) or those from the hippocampus (implicated in the performance of spatial skills) facilitate learning in that system remaining active after pharmacological challenge. The real meaning of this competition among systems still remains unclear, although it has been proposed that both systems have evolved separately in response to distinct needs, which in turn might explain why eventually these systems can compete for the handling of information. Unfortunately, several studies have demonstrated that relationships among memory systems are complicated and poorly understood until now.
]]> Semantic memorySemantic memory mainly refers to information stored on characteristics defining concepts (facts lacking a well–defined spatial/ temporal frame), as well as processes allowing its efficient recovery for further utilization in language and thought. Knowledge about the anatomic location of semantic representations has gained attention with the use of new functional neuroimaging techniques. These studies have shown that the information about the features of specific objects needed to create concepts is stored in the same neuronal systems that remain active during the perception of different stimuli. The way in which this conceptual information is organized remains unclear so far, but there is evidence suggesting that its organization proceeds on the basis of grouped categories of concepts. Pathological studies in patients suffering semantic dementia have also demonstrated that some areas, such as temporal pole and perirhinal cortex, are relevant for semantic processing.
Key words: Memory, semantic, episodic, procedural, working memory.
Resumen
La memoria es una función cerebral fascinante, mediante ella el Sistema Nervioso codifica, almacena, organiza y recupera una gran variedad de tipos de información que resultan de vital importancia para el individuo en particular. Los conocimientos actuales nos permiten conceptualizar a la memoria como conformada por una red compleja de subsistemas de memoria que pueden trabajar en paralelo, cooperando e incluso en ocasiones funcionar de forma competitiva entre sí. La evolución de la clasificación de los sistemas de memoria se ha desarrollado en paralelo al conocimiento acerca del funcionamiento del los procesos mnésicos. Las primeras aproximaciones al estudio de la memoria estaban conformadas por métodos filosóficos que comprendían la observación, reflexión, lógica, etc. En el siglo XIX surgieron los primeros estudios científicos para el estudio experimental de la memoria. Autores como Ebbinghaus o Lashley estudiaron por primera vez la memoria humana y animal respectivamente. Los conductistas como Pavlov, Skinner, Thorndike y Watson sentaron las bases del aprendizaje asociativo que conocemos como condicionamiento clásico y condicionamiento operante. Más tarde los estudios neuropsicológicos de pacientes con lesiones quirúrgicas focales temporales arrojaron resultados contundentes acerca del sustrato anatómico de la memoria declarativa en el lóbulo temporal, lo que inició una avalancha de estudios y descripciones neuropsicológicas cada vez más finas sobre las consecuencias de las lesiones y patologías cerebrales en los distintos procesos de memoria. Más recientemente, los estudios de los procesos celulares y moleculares de las formas de aprendizaje más elementales (habituación y sensibilización) en modelos de animales invertebrados han demostrado los requerimientos celulares mínimos para el establecimiento del aprendizaje así como los mecanismos moleculares diferenciales involucrados en la memoria de corto y largo plazo. A últimas fechas, la introducción de los estudios de neuroimagen funcional en pacientes enfermos y sanos ha permitido la expansión de los conocimientos sobre el funcionamiento dinámico y en tiempo real de los diversos procesos de memoria. En la actualidad la clasificación más aceptada de los sistemas de memoria de largo plazo considera dos grandes esferas principales: la memoria declarativa y la no declarativa. La memoria declarativa se refiere a la que contiene información de la cual se tiene un registro consciente y que se puede verbalizar o transmitir fácilmente a través de algún medio a otro individuo. La memoria no declarativa comprende información que no se puede verbalizar fácilmente o cuyo aprendizaje puede ser inconsciente e incluso involuntario. La memoria declarativa se subdivide en memoria semántica y episódica. El ámbito de la memoria semántica es la información almacenada acerca de las características y atributos que definen los conceptos (hechos que carecen de un marco espacio temporal definido), así como los procesos que permiten su recuperación de forma eficiente para su utilización en el pensamiento y el lenguaje actual. Los estudios de imagen funcional han demostrado que la información sobre las características de objetos específicos que es necesaria para la generación de conceptos es almacenada dentro de los mismos sistemas neuronales que están activos durante la percepción de esos mismos estímulos. El rol del lóbulo temporal en esta variedad de memoria está comprobado por estudios experimentales y clínicos, pero los estudios de imagen funcional han demostrado otras áreas asociadas a la codificación y recuperación semántica cuyo papel aún no ha sido comprendido por completo.
Palabras clave: Memoria, semántica, episódica, procedimental, memoria de trabajo.
INTRODUCCIÓN
La memoria es sin duda un proceso cerebral notable pues, mediante él, el Sistema Nervioso codifica, organiza y almacena los sucesos pasados de tal forma que en ocasiones permite recordar de manera consciente eventos en el pasado distante tan vívidamente como si se experimentaran nuevamente. Estos recuerdos con frecuencia traen consigo experiencias emocionales intensas que van desde lo maravilloso hasta lo atemorizante. La experiencia presente está inexorablemente entretejida con los recuerdos, y el significado de las personas, de las cosas y de los eventos del presente depende de algún modo de la experiencia previa, es decir, de la memoria, de modo que el presente pierde su realce sin el pasado.1 La memoria no es una función cerebral estática, única o aislada, se comporta más bien como un conjunto de funciones cerebrales distintas pero estrechamente interrelacionadas que están orientadas hacia un mismo fin, por lo que resulta más correcto denominarla en términos de sistemas de memoria. Podría intentar definirse someramente a la memoria como un grupo de funciones cerebrales que tienen la tarea de clasificar, codificar, almacenar y recuperar una gran diversidad de tipos de información que resultan de importancia para el organismo en particular. La taxonomía de estos sistemas ha evolucionado paralelamente al conocimiento de las bases anatómicas y fisiológicas de la memoria.1,2 Debido a su gran extensión, el estudio correcto de los procesos de memoria ha sido tema de innumerables textos especializados a lo largo del tiempo, ya que se trata de un tema que puede abordarse desde muy distintos puntos de vista (molecular, celular, neuroquímico, anatómico, evolutivo, neuropsicológico, etc.) y se encuentra en constante actualización. El presente artículo no pretende constituirse como un tratado sobre los procesos que componen o generan la memoria, sino únicamente realizar una breve reseña sobre la evolución histórica del estudio de los procesos de memoria, enunciar las bases de la taxonomía actual de una forma sencilla y práctica, y destacar algunos puntos de polémica y debate actuales al respecto.
]]> Antecedentes y evolución históricaEl estudio teórico de la memoria y el aprendizaje se inició desde los tiempos de Platón y Aristóteles. En las teorías del conocimiento de este último se hace alusión por primera vez a distintos niveles de conocimiento (conocimiento sensible vs. entendimiento), pues para Aristóteles el verdadero saber estaba más allá de la sensación y la simple experiencia, pues además implica el conocimiento acerca de la causa y motivo de los sucesos u objetos; además señaló que el fundamento del aprendizaje y la memoria son las asociaciones entre dos sucesos (como ocurre entre el rayo y el trueno);3 sin embargo, hasta antes del siglo XIX el estudio de la memoria sólo se limitaba a métodos más empíricos y filosóficos que científicos, es decir, para su estudio se utilizaban métodos como la lógica, la introspección, la comparación, la reflexión, etc.2 Fue hasta el siglo XIX que comenzaron los primeros reportes y estudios respecto a la memoria y sus trastornos. En realidad, la noción sobre los distintos tipos o sistemas de memoria no es nueva y se puede encontrar ya en la literatura de los siglos XVIII y XIX, en escritos psicológicos que distinguen, por ejemplo, la memoria de los hábitos (habilidades mecánicas).2 De esta época son de destacar los siguientes autores: Hermann Ebbinghaus, quien en 1885 fue el primero en realizar estudios experimentales sobre la memoria de repetición verbal en seres humanos, utilizando silabas sin sentido, y además describe por primera vez la mejoría progresiva del rendimiento durante la adquisición de nuevas tareas o <<curva de aprendizaje>>;4 William James (1890), quien en su Tratado de Psicología, fue pionero al proponer la distinción entre memoria de corto y largo plazo;5 Sergei Korsakoff, quien describe junto con Carl Wernicke, el síndrome amnésico (acompañado de ataxia y oftalmoplejía), que actualmente lleva su nombre (Síndrome de Wernicke–Korsakoff) y además propone el estudio de los trastornos de memoria (amnesias) como un medio importante para conocer los procesos mnemónicos normales;6,7 pero quizá uno de los exponentes más importantes de esta época, y menos reconocidos en el medio científico, es el biólogo alemán Richard Semon, al que se atribuye la autoría de uno de los términos más utilizados en la bibliografía actual de la memoria: el engrama. De hecho, en sus teorías distinguió tres aspectos diferentes que componían los procesos de memoria: la engrafía, que representaría el proceso de codificación de la información, el engrama que representaría todos los cambios que ocurren en el Sistema Nervioso y que preservan los efectos de la experiencia, y la ecforia, que representaría la recuperación de información; en su teoría también propuso que para que ocurriera la ecforia de forma eficaz (recuperación) sería necesario que se reunieran nuevamente las condiciones que prevalecían en el momento de la engrafía (adquisición); además también propuso ideas novedosas sobre el beneficio de la repetición en la memoria, sin embargo, su teoría recibió muy poco apoyo en su época lo cual se ha reflejado en el desconocimiento de sus contribuciones hasta nuestros días.8 A pesar de todas estas contribuciones, hasta este momento a los procesos de memoria no se les reconocía un sustrato anatómico específico dentro del Sistema Nervioso. La primera asociación anatómica entre las lesiones cerebrales focales (específicamente del lóbulo temporal) y la memoria, se atribuye a Bekhterev, en 1899,9 quien demostró en un paciente cuyo principal problema era una alteración grave de la memoria reciente, la presencia de lo que él denominó <<reblandecimiento>> de áreas corticales específicas como el uncus, el hipocampo y áreas adyacentes de la corteza cerebral temporal.9,10
A principios del siglo XX, la corriente científica del <<conductismo>>, con sus principales exponentes Thorndike, Pavlov, Watson y Skinner, estudiaron las características y componentes de un tipo particular de aprendizaje y memoria: el que se deriva de la asociación repetida entre un estímulo y una respuesta (condicionamiento clásico) o entre un estímulo y una conducta (condicionamiento operante).11–13 Sin duda los conocimientos aportados por la corriente del conductismo fueron de los más importantes en los inicios del siglo XX, sin embargo, merece la pena señalar las aportaciones del novelista francés Marcel Proust (1913) quien realizó descripciones teóricas trascendentes y muy novedosas acerca del fenómeno de la experiencia consciente. En su serie de novelas que reciben el nombre de <<En busca del Tiempo perdido>> se establecen las bases de lo que él denominó <<memoria involuntaria>>, que consistía en una memoria basada en la adquisición supraconsciente de información, es decir, información cuya adquisición no ha sido dirigida por la atención o por la volición inteligente del sujeto, y a la cual eventualmente puede tenerse acceso mediante algún estímulo externo que la hace evidente.14 Estas teorías estaban sin duda adelantadas a su tiempo y conforman hasta la actualidad uno de los terrenos mas fértiles de polémica en el estudio de la memoria y su relación con la conciencia.
A pesar de todas estas aportaciones científicas fue quizá Karl Lashley, en 1929,15 el que inició formalmente el estudio experimental de la memoria con su investigación llevada a cabo en ratas, encontrando que el deterioro en el desempeño en laberintos se correlacionaba más con la extensión de las lesiones cerebrales que con la localización de las mismas. Con estos resultados se sostuvo la hipótesis de la <<equipotencialidad>> de las áreas corticales la cual establecía que, con excepción de las áreas motoras y sensitivas primarias, al resto de las funciones cerebrales complejas contribuían de igual forma todas las áreas corticales.15 Dicha hipótesis se encuentra en oposición directa con el concepto actual sobre los distintos sistemas y sustratos anatómicos para la memoria, aunque se trató sin duda de la primera aproximación realmente científica al estudio experimental de la memoria en el laboratorio. Simultáneamente a esta propuesta se describieron cada vez más casos clínicos en los que, contrariamente a lo establecido por esta teoría, se demostraba una asociación entre las lesiones del lóbulo temporal y las alteraciones de la memoria, así como diversos trastornos de memoria asociados a secuelas de infecciones cerebrales con afectación del lóbulo temporal.16,17 En 1949, Donald Hebb publicó su ya famoso libro The organization of behavior, a neuropsychological theory,18 donde propuso por primera vez que la memoria podría estar conformada por subsistemas de almacenamiento transitorio de información dependientes de circuitos neuronales redundantes, y otros sistemas más perdurables ligados a cambios estructurales en las neuronas tanto pre– como postsinápticas, siendo ésta la primera propuesta que aseguraba que la memoria se encontraba conformada por subsistemas de memoria, además de sugerir el sustrato morfofisiológico de la memoria de largo plazo.18
Sin duda, una evidencia trascendental en el estudio de la memoria provino de la observación de las alteraciones neuropsicológicas resultantes de las lesiones quirúrgicas o traumáticas del lóbulo temporal. En 1958, Wilder Penfield y Brenda Milner19 describieron dos casos en los que se había realizado resección de las estructuras mediales del lóbulo temporal como tratamiento para mejorar la intensidad de las crisis epilépticas, y encontraron como secuela postquirúrgica una severa alteración para el aprendizaje de material nuevo (amnesia anterógrada), sin alteración importante del rendimiento intelectual general. Estos pacientes conservaban además la atención y la memoria inmediata.19 Más aún, en los casos en los que la resección fue unilateral, se encontró alteración de la memoria de forma selectiva dependiendo del tipo de material recordado; esto es, para el material verbal cuando la resección fue izquierda, y para el material no verbal (visual) cuando fue derecha.19 Los reportes de estos casos llamaron la atención de William Scoville, quien también realizaba una cirugía de resección temporal–medial bilateral (como alternativa a la lobotomía frontal para el alivio de la psicosis grave en pacientes con esquizofrenia). Este autor obtuvo resultados postquirúrgicos en la memoria semejantes a los reportados por Penfield y Milner.20 A consecuencia de esto, Milner fue invitada a estudiar los casos de Scoville y logró documentar amnesia anterógrada en todos los casos en los que la cirugía incluía el hipocampo y el giro del parahipocampo; sin embargo, no encontró ninguna alteración residual de la memoria cuando la cirugía sólo se limitaba al uncus y la amígdala20 lo cual concordaba con reportes previos de Sawa et al.21 El estudio, la descripción detallada y el seguimiento de uno de los casos de Scoville (el famoso paciente HM),20 sentaron un antecedente muy importante en el estudio de la memoria, contradiciendo de forma casi definitiva la noción de la <<equipotencialidad>> cortical cerebral, además de ayudar a esclarecer la participación del lóbulo temporal en los mecanismos de la memoria explícita.22 Se pueden consultar descripciones y análisis neuropsicológicos más detallados de este caso en diversos artículos publicados con anterioridad.20,22–26
Por otro lado, el estudio de los pacientes con amnesia también puso en evidencia que al afectarse selectivamente la memoria para hechos recientes y conservarse relativamente intacta la memoria para hechos más remotos, existía (como ya se sospechaba), una diferencia entre el sustrato anatómico de las memorias de corto y de largo plazo. Además se documentaron disociaciones importantes en cuanto al manejo de la información en la memoria según el contenido de lo que es recordado; es decir, que existían algunos aspectos de ella que parecían no afectarse en los pacientes amnésicos; por ejemplo, en el estudio del paciente HM se encontró que éste era capaz de retener un número de tres cifras por un período aproximado de 15 minutos si se mantenía repitiendo dicha cifra y no se distraía su atención con otro estímulo, más allá de lo cual olvidaba por completo la información e incluso el recuerdo de haber memorizado algo.25 Esta disociación ya había sido propuesta con anterioridad por Drachman y Arbit en 1966.27 En un estudio posterior se demostró que incluso se producían resultados contrarios cuando se asignaba la tarea de recordar material que no se podía verbalizar con facilidad; en otras palabras, no podía recordar material que no era capaz de repetir y reforzar verbalmente. Más tarde también se demostró que el aprendizaje de tareas o habilidades específicas (como la escritura en espejo) se mantenía normal en las lesiones témporo–mediales, lo cual hacia suponer que el aprendizaje de hábitos y tareas motoras podía adquirirse de forma independiente del lóbulo temporal.28
Posteriormente, en 1968, Warrington y Weiskrantz,29 usando una versión de la prueba de figuras incompletas de Gollin30, demostraron que el aprendizaje de tipo perceptual está preservado en los pacientes con amnesia anterógrada, una forma de aprendizaje que posiblemente tenga su origen en áreas visuales de asociación de orden superior.29 Basándose en estos hallazgos clínicos, comprobados además en modelos experimentales de lesiones mediales–temporales,31,32 Larry Squire propuso en 1986 una taxonomía de la memoria, en la cual la dividía en dos grandes subsistemas: <<la memoria declarativa y la no declarativa>>33 (cuadro 1).
Casi al mismo tiempo, Endel Tulving propuso un modelo de la memoria que comprendía no dos, sino tres sistemas que trabajaban de forma conjunta: <<la memoria procedimental, la memoria semántica y la episódica>>.34 Tulving propuso que sólo la primera de ellas (procedimental) podía funcionar de forma aislada, siendo ésta la más elemental y primitiva y que podía estar presente desde los organismos más simples.34 La memoria procedimental comprendería el conocimiento en relación a <<saber cómo hacer las cosas>>, y según el autor, sería una forma un poco más compleja de aprendizaje que la conducta instintiva debido a que no requiere de la asociación estímulo–respuesta. La memoria semántica es la que se denomina como genérica o de hechos particulares, y se asume que no involucra ninguna estructura de eventos o ubicación témporo–espacial; un ejemplo de este tipo de memoria serían los datos concretos que memorizamos como cifras, nombres de objetos, fechas, etc. La memoria episódica es aquella que hace referencia a eventos que ocurrieron en un lugar y tiempo específicos; la recuperación de este tipo de información requiere generalmente de la creación de <<imágenes mentales>> que son evocadas en el momento y cuyo recuerdo puede incluir una gran variedad de tipos de información: visual, auditiva, olfativa, somestésica, temporal, espacial, emotiva, etc. Este proceso puede llegar a recrear el evento pasado de una forma muy vívida en el presente.34 Tulving describió, a su vez, tres submodalidades de estos tres sistemas, así como también lo que llamó el <<nivel de conciencia>> de cada uno de ellos: 1. La adquisición, que como su nombre lo indica, es la forma en que ingresa la información según el tipo de memoria; 2. La representación de la información en el sistema de memoria; es decir, la forma en que se codifica o clasifica la información y 3. La expresión de conocimiento; es decir, la forma en la que la información se exterioriza por el individuo34 (cuadro 2).
]]> Uno de los defectos que presenta este sistema de clasificación es que está basado exclusivamente en la naturaleza de la información almacenada, y no considera que estos sistemas pueden funcionar de forma complementaria y conjunta, siendo a veces difícil distinguirlos en la evaluación sistemática de la memoria. En una clasificación más reciente e incluyente, Tulving y Schacter35 definen cinco sistemas de memoria según los mecanismos cerebrales involucrados, el tipo e información que se maneja y los principios con los que operan cada uno de los sistemas, difiriendo de la propuesta de Squire en lo siguiente: 1. Esta clasificación sólo esta basada en estudios neuropsicológicos realizados en seres humanos; 2. para Tulving, la memoria de corto plazo y la episódica serían de tipo explícito, mientras que las otras serían implícitas; 3. todos los sistemas de memoria expresan funciones cognitivas a excepción de la procedimental; 4. no se incluyen formas más elementales de memoria como la habituación y la sensibilización y, además, 5. Considera la memoria episódica y la semántica como dos subsistemas independientes, proponiendo que la información accede primero a lo que llamó <<sistema de representación perceptual>> (SRP) y posteriormente a la memoria semántica, la cual a su vez, alimentaría la memoria episódica35 (cuadro 3).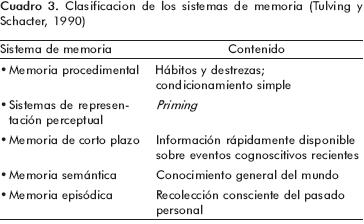
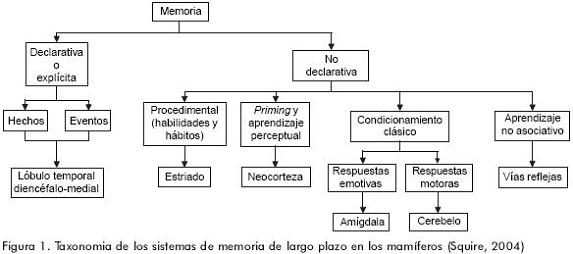
Una de las aportaciones más importantes de la clasificación de Tulving y Schacter es justamente este sistema de representación perceptual (SRP), cuyas expresiones neuropsicológicas serían la identificación perceptual y el fenómeno del priming perceptual, a cuyas características haremos referencia más adelante. En una clasificación más reciente de los sistemas de memoria de largo plazo, Larry Squire2 incluye además las principales estructuras anatómicas relacionadas con cada subtipo de memoria, las categorías de aprendizaje asociativo y no asociativo, así como una distinción entre los componentes emotivos y motores asociados con el condicionamiento clásico2 (figura 1).
El conocimiento sobre las bases moleculares de la memoria
Los estudios iniciales acerca de la memoria fueron llevados a cabo principalmente por filósofos y psicólogos, y aunque desde un inicio algunos biólogos habían ya realizado aportaciones importantes al estudio de la memoria, éstas solían recibir poco apoyo y difusión por la comunidad científica.8 Hasta antes de los años 1960 no existía una cooperación real y productiva entre las disciplinas biológicas y la psicología, hasta que surgió la psicología cognitiva o neurociencia cognitiva, que trataba, por primera vez, no sólo de explicar los procesos mnésicos basados en la conducta experimental observable y manipulable, sino en establecer cuáles eran los procesos celulares, y más tarde moleculares, responsables de generar y almacenar esa conducta, es decir, buscar el verdadero engrama de la memoria en el cerebro.36 Los fundadores de esta neurociencia cognitiva fueron George Miller, Ulric Neisser, Herbert Simons, entre otros, en la década de 1960,36 sin embargo, haciendo un análisis retrospectivo, quizá la primera propuesta acerca de los mecanismos celulares del aprendizaje la propuso Santiago Ramón y Cajal, al sugerir que el aprendizaje debe surgir del fortalecimiento en la comunicación entre las neuronas ya existentes, ya que éstas (como él observó) han perdido su capacidad de multiplicarse.37 Con el interés generado por el surgimiento de las neurociencias cognitivas también se desarrollaron múltiples técnicas experimentales para el estudio neurobiológico de los procesos cognitivos. Por ejemplo, en 1970 Evarts y Mountcastle desarrollaron una técnica para el registro de la actividad eléctrica de neuronas únicas en animales vivos y en movimiento libre,38 estas técnicas y las desarrolladas más tarde por Robert Wurtz, Apostolos Georgopoulos y William Newsome, produjeron los primeros estudios que correlacionaban la actividad neuronal con procesos cognitivos como la percepción, la atención y la toma de decisiones.36,39 Hubo también un resurgimiento de los estudios anatómicos correlativos, con nuevas técnicas de visualización de tractos nerviosos, y el posterior desarrollo de las técnicas de resonancia magnética y tomografía por emisión de positrones, con las cuales se complementó el estudio anatómico de los procesos cognitivos en pacientes tanto enfermos como sanos e intactos.40 Sin embargo, uno de los estudios más importantes sobre los mecanismos moleculares de la memoria provino de investigaciones relacionadas con las formas más elementales de la memoria no declarativa: los procesos de habituación, deshabituación y sensibilización; éstos se estudiaron en animales invertebrados con un Sistema Nervioso muy simple, conformado sólo por entre 20 000 a 100 000 células. El ejemplo más representativo es la Aplysia, aunque también se usaron otros organismos como la Hermissenda, la Pleurobranchea, la Limax e incluso la Drosophila.41 Las aportaciones más destacadas de estos estudios pueden resumirse en la demostración de los ensambles celulares mínimos requeridos para el aprendizaje más elemental, así como los mecanismos moleculares y neuroquímicos responsables de la memoria de corto y largo plazo. Es decir, se demostró por primera vez que los cambios estructurales responsables de la memoria de corto plazo suelen ser modificaciones covalentes de proteínas ya existentes producidas por una u otra cinasa de segundos mensajeros; mientras que la memoria de largo plazo implica la síntesis de nuevas proteínas y que el <<switch molecular>> que media el cambio de memoria de corto a largo plazo está representado por segundos mensajeros como el cAMP, proteínas señalizadoras como la MAPK o PKA, y factores de trascripción génica como el CREB.36 Sin embargo, los estudios sobre los mecanismos moleculares responsables de la memoria continúan en marcha, y recientemente se ha demostrado que puede existir almacenamiento de información a largo plazo a pesar de una extensa inhibición en la síntesis de proteínas.42 Revisar los avances recientes en el terreno de los mecanismos moleculares de la memoria trasciende a los objetivos del presente artículo, pero se pueden encontrar en la bibliografía diversas actualizaciones del tema.43–45
TAXONOMÍA Y TÉRMINOS DE MEMORIA
]]> Desde luego, la evolución y transformación histórica de los sistemas de clasificación no ha terminado aún, los sistemas de memoria hasta aquí mencionados forman parte de los más aceptados y difundidos en su utilización hasta el momento actual; sin embargo, los psicólogos utilizan una gran diversidad de términos para referirse a distintas variedades, características o niveles de procesos mnésicos, por lo que en la literatura especializada podemos encontrar conceptos que pueden llegar a hacer confusa la conceptualización integral de la memoria y sus distintos componentes.46 Entre estos conceptos hay diversas dicotomías, como la mencionada respecto a la memoria declarativa y la no declarativa, pero existen muchas otras: procesos de memoria consciente vs. inconsciente; memoria de corto vs. largo plazo; memoria implícita vs. explícita; memoria retrospectiva vs. prospectiva; recuerdo voluntario vs. involuntario; aprendizaje intencional vs. incidental; así como diversos términos que hacen alusión a la memoria de corto plazo (de trabajo, activa) o a diversas variedades de memoria explícita de largo plazo: semántica, episódica, autobiográfica colectiva (para una revisión extensa véase: Roediger, 2008).46 Las definiciones y alcances de cada uno de estos términos en ocasiones son imprecisas y con frecuencia no son completamente excluyentes entre sí, pero aun así dejan ver que el estudio de los procesos mnésicos puede hacerse desde muy distintos puntos de vista.Hasta este momento, incluso la división inicial de los sistemas de memoria (declarativa y no declarativa) continúa siendo materia de debate debido a las distintas variedades de memoria que se han observado disociadas en estudios tanto en humanos como en animales que llegan a compartir características de ambos tipos de memoria. Esta dicotomía inicial de los sistemas es, sin embargo, muy simplista para lo que se ha demostrado en la actualidad.2 Conceptualizar estos sistemas como aislados resulta un grave error pues es claro que todos los sistemas de memoria trabajan en paralelo, ocasionalmente de forma independiente, a veces cooperando e incluso también de forma competitiva.46 Resulta evidente que la participación de los distintos sistemas de memoria no siempre es la misma, incluso cuando dos individuos distintos realizan exactamente la misma tarea de aprendizaje, lo cual pone de manifiesto que la codificación, motivación o abordaje inicial de la información puede determinar si ésta es procesada como una información procedimental, espacial, semántica o de otro tipo.2 La descripción reciente de las relaciones competitivas de los sistemas de memoria (declarativa vs. procedimental) resultó un hallazgo sorprendente; pero ahora existe evidencia clara (tanto en animales de experimentación como en humanos), de que la disminución de las funciones de un sistema de memoria procedimental, espacial o declarativo, puede favorecer el desempeño de otro sistema de memoria.47 En estudios iniciales, la inactivación transitoria de estructuras estriatales dorsales (concernidas en el desempeño de habilidades y tareas motoras) o del hipocampo (concernido en el desempeño de tareas espaciales), facilitó el desempeño en el aprendizaje del sistema que no fue inactivado farmacológicamente.47–52 Por otro lado, en estudios de imagen funcional se ha encontrado además que la participación de las distintas estructuras cerebrales (estriado–hipocampo) puede ofrecer un comportamiento diferencial en relación con el tiempo, demostrándose que en el aprendizaje inicial de las tareas perceptuales o motoras existe un aumento de la activación del hipocampo, con poca participación del estriado, en tanto que se observa el patrón de activación inverso conforme la tarea se repite y se aprende.47,51 Más aún, se ha comprobado que existe una importante modulación emocional para la utilización predominante de algunos sistemas mnésicos.53 El significado real de esta competencia entre algunos sistemas aún no se conoce con precisión, pero se ha propuesto que ambos sistemas han evolucionado por separado en respuesta a distintas necesidades, lo cual explicaría por qué eventualmente pueden encontrarse compitiendo por el manejo de la información. Estos y otros estudios han demostrado que las relaciones entre los distintos procesos de memoria son complejas, y no del todo comprendidas.47 A continuación se dará una breve explicación sobre los principales procesos componentes de los sistemas de memoria, según las clasificaciones antes comentadas.
SISTEMAS DE MEMORIA DE LARGO PLAZO
Memoria declarativa
Memoria semántica: El ámbito de la memoria semántica es la información almacenada sobre las características y atributos que definen los conceptos, así como los procesos que permiten su recuperación de forma eficiente para su utilización en el pensamiento y el lenguaje.54 El conocimiento del sitio anatómico en donde se producen las representaciones semánticas se ha revolucionado con el uso de los estudios de neuroimagen funcional. Estos estudios han demostrado que la información sobre las características de objetos específicos que es necesaria para la generación de conceptos es almacenada dentro de los mismos sistemas neuronales que están activos durante la percepción de esos mismos estímulos.55 Esto se ha comprobado en estudios con tareas de nominación de colores, nominación de acciones, objetos con formas similares, objetos manipulables hechos por el hombre (herramientas), etc.55 Pero ¿cómo es que se organiza la información sobre la forma de los objetos? los estudios de imagen han demostrado que se activan áreas específicas dentro de la corteza occípitotemporal cuando se utilizan ciertas categorías de objetos; por ejemplo, objetos vivos o animales (giro fusiforme lateral) vs. objetos inanimados (giro fusiforme medial).56 Incluso estudios de registro unitario con electrodos profundos en pacientes epilépticos han mostrado que las neuronas responden más a una categoría específica que a un estímulo especifico57, sin embargo, como se demostró posteriormente, este esquema de organización no siempre funciona así,58 lo cual ha sugerido que las representaciones de diferentes categorías de objetos están distribuidas ampliamente y pueden traslaparse entre sí, dando como resultado una gran diversidad de categorías potenciales.54
Es claro que el <<espacio neuronal cortical>> para almacenar todas las posibles categorías de objetos es muy limitado, por lo que debe existir alguna organización flexible (quizá basada en algunas características comunes) o priorizada para las categorías que resulten más importantes, pero si esto es así o no, aún es tema de debate.53 El surco temporal superior se ha asociado con la representación de movimiento de objetos y el área ventral de la corteza premotora izquierda con la representación de objetos que están asociados a movimiento.
Por otro lado, el área prefrontal inferior izquierda sería, según algunos investigadores, la responsable de la recuperación, mantenimiento, monitoreo y manipulación de las representaciones semánticas que están almacenadas en cualquier sitio; es decir, el equivalente a una memoria de trabajo semántica.59 De esta manera se han documentado asimetrías hemisféricas en el procesamiento semántico, encontrándose que el hemisferio izquierdo se centra en las palabras con un significado concreto, mientras que el derecho se asocia más con el procesamiento de palabras léxicamente ambiguas.60 En estudios recientes se ha demostrado que la activación de estructuras del lóbulo temporal (hipocampo, amígdala, corteza témporopolar) disminuye gradualmente conforme se recupera información semántica cada vez más antigua en su adquisición, observándose el patrón de activación inverso en los lóbulos parietales y frontales, lo cual sugiere que el lóbulo temporal tiene un papel limitado en el tiempo en la recuperación semántica.61 La relación entre memoria semántica y el lóbulo temporal también ha sido comprobada por estudios neuropatológicos de pacientes con demencia semántica, en los cuales se ha demostrado una atrofia significativa (59–65%) de estructuras temporales como el polo temporal y la corteza perirrinal, así como en otras áreas: corteza órbitofrontal, giro fusiforme, ganglios basales y sustancia blanca. En contraste, los estudios neuropatológicos no han demostrado asimetrías hemisféricas en los patrones de atrofia cerebral.62 También se ha demostrado recientemente, en estudios de resonancia magnética funcional, que en pacientes adultos con lesiones temporales izquierdas existe una verdadera reorganización de las áreas corticales involucradas en el procesamiento semántico normal, dicha reorganización depende de la categoría semántica especifica de objetos (vegetales, animales, etc.) que tenía asignada el área cortical dañada y de las condiciones de las áreas corticales que permanecieron indemnes para suplir la actividad de las áreas corticales dañadas.63 El estudio de la memoria semántica aún no esta completo, pero los estudios de imagen funcional han revolucionado el conocimiento de estos procesos en los pacientes sanos; todavía quedan por dilucidar los mecanismos precisos de categorización semántica, así como la participación del resto de las áreas corticales y de las estructuras subcorticales que se activan durante el proceso de la categorización semántica.
En la segunda parte del presente trabajo se profundizará acerca del resto de los sistemas de memoria declarativa y no declarativa, así como de la memoria de trabajo.
]]> REFERENCIAS
1. Rains, Dennis G. Sistemas de memoria. En: Principios de neuropsicología humana. Primera edición. México: Mc Graw Hill; 2004; pp: 241–286. [ Links ]
2. Squire LR. Memory systems of the brain: A brief history and current perspective. Neurobiol Learn Mem 2004;82:171–177. [ Links ]
3. Aristoteles. Metafísica de Aristoteles. Segunda edición. Madrid: Edit. Gredos; 1997. [ Links ]
4. Ebbinghaus H. Memory. A contribution to experimental psychology. New York: Columbia University; 1885. [ Links ]
5. James W. The principles of psychology. Tercera edición. United Kingdom: Courier Dover Publications; 1950. [ Links ]
6. Shacter DL. Implicit memory: History and current status. J Exp Psychol: Learn Mem Cog 1987;13:501–518. [ Links ]
7. Zubaran C, Fernandes JG, Rodnight R. Wernicke–Korsakoff Syndrome. Postgrad Med 1997;73:27–31. [ Links ]
8. Schacter DL. Forgotten ideas, neglected pioneers: Richard Semon and the history of memory. Philadelphia: Psychology Press; 2001. [ Links ]
9. von Bekhterev M. Demonstration eines gehirns mit zerstörung der vorderen und inneren theile der hirnrinde beider schläfenlappen. Neurologisches Zeitblatt 1900;19:990–1. [ Links ]
10. Milner B. The medial temporal–lobe amnesic syndrome. Psychiatr Clin N Am 2005;28:599–611. [ Links ]
11. Thorndike EL. Memory for paired associates. Psychological Rev 1909;15:122–138. [ Links ]
12. Pavlov IP. Conditioned reflex. London: Oxford University Press; 1927. [ Links ]
13. Skinner BF. Drive and reflex strength: I J Gen Psychol 1932;6:22–37. [ Links ]
14. Epstein R. Consciousness, art, and the brain: Lessons from Marcel Proust. Consciousness Cognition 2005;13:213–240. [ Links ]
15. Lashley KS. Brain mechanism and intelligence: A Quantitative study of injuries to the brain. Chicago: Univ of Chicago Press; 1929. [ Links ]
16. Victor M, Angevine JB, Mancall EL, Fisher CM. Memory loss with lesions of hippocampal formation. Arch Neurol 1961;5:244–263. [ Links ]
17. Rose FC. Persistent memory defect following encephalitis. Brain 1960;83:195–212. [ Links ]
18. Hebb DO. The organization of behavior. New York: Wiley; 1949. [ Links ]
19. Penfield W, Milner B. Memory deficits induced by bilateral lesions in the hippocampal zone. Arch Neurol Psychiatry 1958;79:475–97. [ Links ]
20. Scoville WB, Milner B. Loss of recent memory after bilateral hippocampal lesions. J Neurol Neurosurg Psychiatry 1957;20:11–21. [ Links ]
21. Sawa M, Ueki Y, Arita M, Harada T. Preliminary report on the amygdaloidectomy on the psychotic patients, with interpretation of oral–emotional manifestation in schizophrenics. Folia Psychiatrica Neurologica Japonica 1954;7:309–29. [ Links ]
22. Scoville WB. The limbic lobe in man. J Neurosurg 1954;11:64–6. [ Links ]
23. Milner B. The memory defect in bilateral hippocampal lesions. Psychiatric Research Report 1959;11:43–58. [ Links ]
24. Milner B. Disorders of learning and memory after temporal lobe lesions in man. Clin Neurosurg 1972;19:421–46. [ Links ]
25. Milner B, Corkin S, Teuber HL. Further analysis of the hippocampal amnesic syndrome 14–year follow–up study of HM. Neuropsychologia 1968;6:215–34. [ Links ]
26. Scoville WB, Correll RE. Memory and the temporal lobe. Acta Neurochirurgica 1973;28:251–258. [ Links ]
27. Drachman DA, Arbit S. Memory and the hippocampal complex II. Is the memory a multiple process? Arch Neurol 1966;15:52–61. [ Links ]
28. Corkin S. Acquisition of motor skills after bilateral temporal–lobe excision. Neuropsychologia 1968;6:255–65. [ Links ]
29. Warrington E, Weiskrantz L. New method for testing long–term retention with special reference to amnesic patients. Nature 1968;4:217:972. [ Links ]
30. Gollin ES. Developmental studies of visual recognition of incomplete objects. Percept Mot Skills 1960;11:289–98. [ Links ]
31. Mishkin M. Memory in monkeys severely impaired by combined but not by separate removal of amygdala and hippocampus. Nature 1978;273:297–298. [ Links ]
32. Zola–Morgan S, Squire LR. Preserved learning in monkeys with medial temporal lesions: Sparing of motor and cognitive skills. J Neurosci 1984;4:1072–1085. [ Links ]
33. Squire LR. Mechanisms of memory. Science 1986;232:1612–1619. [ Links ]
34. Tulving E. How many memory systems are there? American Psychologist 1985;40:385–398. [ Links ]
35. Tulving E, Schacter DL. Priming and human memory systems. Science 1990;247:301–306. [ Links ]
36. Milner B, Squire LR, Kandell ER. Cognitive neuroscience and the study of memory. Neuron 1998;20: 445–468. [ Links ]
37. Cajal SR. La fine structure des centres nerveux. Proc R Soc Lond 1894;55:444–468. [ Links ]
38. Mountcastle V. Modality and topographic properties of single neurons of cat somatosensory cortex. J Neurophysiol 1957;20:408–434. [ Links ]
39. Wurtz RH. Steady potential shifts during arousal and sleep in the cat. Electroencephalograp Clin Neurophysiol 1965;18:649–662. [ Links ]
40. Brownell GL, Sweet WH. Localization of brain tumors with positron emitters. Nucleonics 1953;11:40–45. [ Links ]
41. Kandell ER, Tauc L. Mechanism of prolonged heterosynaptic facilitation. Nature 1964;2002:145. [ Links ]
42. Routtenberg A, Rekart JL. Post–translational modification as the substrate for long–lasting memory. Trends Neurosci 2005;28:12–19. [ Links ]
43. Gold PE. Protein synthesis inhibition and memory: Formation vs. amnesia. Neurobiol Learn Mem 2008;89:201–211. [ Links ]
44. Hernandez PJ, Abel T. The role of protein synthesis in memory consolidation. Progress amid decades of debate. Neurobiol Learn Mem 2008;89:293–311. [ Links ]
45. Bailey CH, Barco A, Hawkins RD, Kandell ER. Molecular studies of learning and memory in Aplysia and the hippocampus: A comparative analysis of implicit and explicit memory storage. En: Byrne JH (ed.) Learning and memory: A comprehensive reference. EUA: Academic Press; 2008; vol. 2; pp.11–29. [ Links ]
46. Roediger III HL, Zaromb FM, Goode MK. A typology of memory terms. En: Byrne JH (ed.). Learning and memory: A comprehensive reference. EUA: Academic Press; 2008; vol. 1; pp.11–24. [ Links ]
47. Poldrack RA, Packard MG. Competition among multiple memory systems: converging evidence from animal and human brain studies. Neuropsychol 2003;1497:1–7. [ Links ]
48. Packard MG, Hirsh R, White NM. Differential effects of fornix and caudate nucleus lesions on two radial maze task: evidence for multiple memory systems. J Neurosci 1989;9:1465–72. [ Links ]
49. Packard MG, McGaugh JL. Double dissociation of fornix and caudate nucleus lesions on acquisition of two water maze tasks: further evidence for multiple memory systems. Behav Neurosci 1992;106:439–46. [ Links ]
50. Packard MG, McGaugh JL. Inactivation of hippocampus or caudate nucleus with lidocaine differentially affects expression of place and response learning. Neurobiol Learn Mem 1996;65:65–72. [ Links ]
51. Poldrack RA, Prabakharan V, Seger C, Gabrieli JDE. Striatal activation during cognitive skill learning. Neuropsychol 1999;13:564–74. [ Links ]
52. Packard MG. Exhumed from thought: Basal ganglia and response learning in the plus maze. Behav Brain Res 2009 (en prensa). [ Links ]
53. Wingard JC, Packard MG. The amygdala and emotional modulation of competition between cognitive and habit memory. Behav Brain Res 2008;193:126–131. [ Links ]
54. Martin A Chao LL. Semantic memory and the brain: Structure and processes. Curr Neurobiol 2001;11:194–201. [ Links ]
55. Martin A, Haxby JV, Lalonde FM, Wiggs CL, Ungerleider LG. Discrete cortical regions associated with knowledge of color and knowledge of action. Science 1995;379:649–652. [ Links ]
56. Caramazza A, Shelton JR. Domain specific knowledge systems in the brain: the animate–inanimate distinction. J Cog Neurosci 1998;10:1–34. [ Links ]
57. Kreiman G, Koch C, Fried I. Category–specific visual responses of single neurons in the human medial temporal lobe. Nat Neurosci 2000;3:946–953. [ Links ]
58. Ishai A, Ungerleider LG, Martin A, Schouter JL, Haxby JV. Distributed representation of objects in the ventral visual pathway. Proc Natl Acad Sci USA 1999;96:9379–9384. [ Links ]
59. Poldrack RA, Wagner DA, Prull MW, Desmond JE, Glover GH et al. Functional specialization for semantic and phonological processing in the left inferior frontal cortex. Neuroimage 1999;10:15–35. [ Links ]
60. Faust M, Ben–Artzi E, Harel I. Hemispheric asymmetries in semantic processing: Evidence from false memories for ambiguous words. Brain Lang 2008;105:220–228. [ Links ]
61. Smith CN, Squire L. Medial temporal lobe activity during retrieval of semantic memory is related to the age of memory. J Neurosci 2009;29:930–938. [ Links ]
62. Davies RR, Halliday GM, Xuereb JH, Kril JJ, Hodges JR. The neural basis of semantic memory: Evidence from semantic dementia. Neurobiol Aging 2008 (en prensa). [ Links ]
63.Leonard B, de Partz MP, Grandin C, Pillon A. Domain–specific reorganization of semantic processing after extensive damage to the left temporal lobe. NeuroImage 2009;45:572–586. [ Links ] ]]>