
1.5m, 0.9 m and 1.2 m respectively. It was possible to arrange different groups of soil Macrofauna according with their ranges of tolerance to environmental variables. Therefore, it is suggested that these patterns do reflect preferences of soil biota to microenvironments and do respond to soil degradation.]]>
Patrones de la macrofauna edáfica en un cultivo de Zea maiz durante la fase postcosecha en "La Mancha", Veracruz, México
Villalobos, F.J.,1,2 R. Ortiz-Pulido1, C. Moreno1, N.P. Pavón-Hernández1, H. Hernández-Trejo1, J. Bello1, y S. Montiel1
1 Instituto de Ecología, A.C., A.P. 63, Xalapa, Veracruz, C.P. 91000, México.
2 Facultad de Ciencias Agropecuarias. Universidad Autónoma del Estado de Morelos. Av. Universidad 1001, Col. Chamilpa, Morelos, C.P. 62210, México.
Recibido: 16 de abril 1998 ]]> Aceptado: 27 de octubre 1999
Resumen
Estudiamos la macrofauna del suelo de un cultivo de Zea maiz durante la fase postcosecha en el Centro de Investigaciones Costeras "La Mancha", Veracruz. Describimos patrones de densidad, distribución espacial y diversidad en relación con la temperatura, humedad y pH del suelo. Para esto conjuntamos cuatro estrategias: a) muestreamos la macrofauna en estratos de 10 cm hasta los 70 cm de profundidad; b) separamos la fauna a nivel de "morfoespecie"; c) estimamos el tamaño de las agregaciones de las "morfoespecies" mediante una técnica de cuadrante-varianza; y d) ordenamos, mediante un análisis de correspondencia canónica, morfoespecies y estratos en un marco de referencia ambiental. La densidad de la macrofauna del suelo estudiado es la más baja reportada hasta la fecha para agroecosistemas en el mundo (246 individuos/m2). Colectamos 46 morfoespecies, que se distribuyeron generalmente en el primer o segundo estrato del suelo y presentaron una distribución agregada. El diametro de las agregaciones de los Oligochaeta juveniles fue superior a 1.5 m y para las larvas de Tenebrionidae y los Diplopoda juveniles fue de 0.9 y 1.2 m, respectivamente. Debido a que es posible separar distintos grupos de acuerdo con sus rangos de tolerancia a la temperatura, pH, humedad y materia orgánica en el suelo, es factible que los patrones de distribución registrados sean un reflejo de las preferencias de la biota a diferentes microambientes y al estado de degradación del suelo.
Palabras clave: Macrofauna, densidad, distribución espacial, diversidad, CICOLMA, Veracruz, maíz.
Abstract
The soil macrofauna of a cornfield was studied during the fallow period in El Centro de Investigaciones Costeras "La Mancha", Veracruz. Patterns of soil macrofauna density, spatial distribution and diversity were described and their relationships with soil temperature, moisture, organic matter and pH were explored. Four strategies were combined to undertake this aim: a) sampling of soil macrofauna was carried out in seven ten cm soil layers from 0 to 70 cm depth; b) soil macrofauna was identified to morphospecies level; c) the size of morphospecies aggregations was determined following a the two-term local quadrat variance method (TTLQV); d) analysis of canonical correspondence was used to arrange morphospecies distribution in an spatial and environmental framework of reference. Density of soil macrofauna in the studied site seems to be the lowest value ever recorded in similar studies (246 individuals m-2). Forty-six morphospecies were collected that are mainly distributed in the top 20 cm soil layer and present an aggregated horizontal pattern of distribution. The diameter of aggregations of Oligochaeta juveniles, larvae of Tenebrionidae and Diplopoda juveniles was >1.5m, 0.9 m and 1.2 m respectively. It was possible to arrange different groups of soil Macrofauna according with their ranges of tolerance to environmental variables. Therefore, it is suggested that these patterns do reflect preferences of soil biota to microenvironments and do respond to soil degradation.
Key words: Soil macrofauna; density; spatial distribution patterns; diversity; CICLOMA; Veracruz; corn.
]]> Introducción
El estudio de patrones en agroecología permite formular preguntas sobre procesos ligados a la producción agrícola. Entre las múltiples implicaciones de tales patrones figura el de conocer bajo que condiciones la fauna del suelo produce efectos benéficos o nocivos en el cultivo y como estos pueden fomentarse o ser evitados mediante prácticas agronómicas. La hipótesis general del programa "Tropical Soil Biology and Fertility" (TSBF) sostiene que la manipulación de la macrofauna puede mejorar las características físicas del suelo y regular el proceso de la descomposición (Anderson e Ingram 1993, Lavelle et al. 1994). Sin embargo, todavía se desconocen aspectos fundamentales sobre la biología de esta fauna, careciendose en muchos casos de las bases para su manipulación.
La función ecológica de la macrofauna del suelo (invertebrados de talla >2 mm) en los agroecosistemas es múltiple. Dichas funciones (directas o indirectas) pueden clasificarse en dos tipos: a) actividades que resultan en una influencia importante sobre el rendimiento vegetal a corto plazo (e.g. herbivoría y dispersión de patógenos); y b) aquellas que tienen una influencia de menor intensidad pero de efectos acumulativos a largo plazo (e.g. humificación, mineralización de la materia orgánica, interacciones con microorganismos e impacto sobre la estructura y propiedades físico-químicas del suelo) (Ryszkowski 1985).
Diferentes estudios se han enfocado a la búsqueda de patrones en la macrofauna bajo condiciones edáficas óptimas (Lavelle 1983, González y Herrera 1983, Lavelle y Pashanasi 1989). La evidencia sugiere que la densidad y diversidad de la macrofauna del suelo es substancialmente menor en los agroecosistemas comparado con ecosistemas menos perturbados (Lavelle et al. 1994, Fraser 1994), y que la mayor actividad biológica tiene lugar en los primeros 20 cm de profundidad del suelo (Lavelle y Kohlman 1984, Villalobos y Lavelle 1990). En agroecosistemas tropicales, se conoce muy poco sobre la distribución de la macrofauna bajo condiciones extremas (e.g. temperatura elevada, humedad baja, acidez, escases de raíces vivas y contenido reducido de materia orgánica) (Amat y Soto 1988).
En este trabajo nos enfocamos a contestar las preguntas: ¿Cuál es la densidad, diversidad y distribución espacial de la macrofauna del suelo en la época seca (i.e., cuando hay una reducción notable en la biomasa de raíces vivas, durante la fase de descanso del cultivo de maíz)? y ¿Cómo influyen en estos patrones la temperatura, pH, humedad relativa y materia orgánica del suelo?
Materiales y métodos
El estudio lo realizamos en un cultivo de maíz localizado en el Centro de Investigaciones Costeras "La Mancha" (CICOLMA), Veracruz, México (96/22' O y 19/36' N). El clima en este lugar es cálido subhúmedo con lluvias en verano. La precipitación total anual varia entre 900 y 1800 mm, con una época marcada de secas de octubre a mayo y una de lluvias de junio a septiembre. La temperatura máxima extrema es de 34/C, la mínima extrema de 16/C y la media anual es de 23/C (Moreno-Casasola 1982, Ortiz-Pulido et al. 1995). Los tipos de vegetación primaria dominantes en la región son selva mediana sudcaducifolia y manglar (Rzedowski 1978). El suelo del sitio estudiado es de origen cuaternario con más del 90% de arena (Blain 1988), existiendo áreas cercanas dedicadas al cultivo de la caña de azúcar.
El sitio donde se realizó el trabajo de campo abarca tres ha y antes del estudio estuvo sometido a siete ciclos sucesivos de monocultivo de maíz. En esta área, antes de la siembra (marzo-mayo) es común que se realize un ablandamiento del suelo ("barbecho") con tractor que alcanza los 30 cm de profundidad, seguido de una cruza y una rastra para desmenuzar el suelo. Durante la fase de cultivo se aplica fertilizante (urea, 160 kg/ha), insecticida ("Nubacron", 0.33 litros/ha) y herbicida ("Hierbamina", 0.66 litros/ha). Raramente se ha observado daño causado por plagas del suelo. Comúnmente la siembra se realiza en junio y se cosecha de noviembre a diciembre, obteniendose una producción anual que varia entre dos y tres toneladas/ha de maíz desgranado. (López-Barradas, com. pers.).
El muestreo se realizó en la época seca (marzo de 1995) siguiendo el método propuesto por el programa TSBF (Anderson e Ingram 1993) con las siguientes modificaciones. Al interior del cultivo se delimitó una franja rectangular de 3 m de largo por 30 cm de ancho y 70 cm de profundidad. Alrededor de esta franja se cavó una zanja de 30 cm de ancho para permitir la separación del suelo por estratos de 10 cm. Los bloques de suelo (unidades muestrales) desprendidos de cada estrato midieron 30 x 30 cm y se tamizaron en una criba (malla de 4 mm2) inmediatamente después de su obtención. En el campo se realizó la separación manual directa de los organismos de cada bloque y se fijaron en alcohol al 70%. La fauna fue depositada en frascos etiquetados con el número de estrato y muestra. En total se revisaron en un día 10 unidades muestrales para cada uno de los siete estratos analizados.
]]> Al iniciar la revisión, en cada unidad muestral se registró temperatura y pH. De cada muestra se tomó una fracción de suelo (aproximadamente 150 gr) que fueron transportadas al laboratorio para determinar su humedad relativa por el método de diferencia de peso (24 h a 105/C). El porcentaje de materia orgánica fue determinada en el Laboratorio de suelos del Instituto de Ecología, A.C., usando la técnica de Walkley & Black.Utilizando un microscopio estereoscópico, los especímenes colectados fueron comparados y separados en morfoespecies. Consideramos una "morfoespecie" como aquella unidad taxonómica por abajo del nivel de Clase que presenta diferencias morfológicas conspicuas con respecto a las otras, o bien a aquellos estados de desarrollo diferentes de una misma especie. Este último criterio se basa en el supuesto de que cada estado de desarrollo de una misma especie tiene una función ecológica cualitativa y cuantitativamente distinta con relación a los otros. La función principal de cada morfoespecie en cada estrato fue determinada de acuerdo con Borror et al. (1981) en cuatro grupos tróficos: herbívoros, saprófagos, carnívoros y parásitos.
La densidad de cada morfoespecie fue cuantificada para cada estrato. Se compararon, mediante un ANDEVA de una vía (Zar 1984) con un nivel de significancia de P<0.05: a) las densidades por estrato para todas las especies y para cada una de las morfoespecies más abundantes, y b) temperatura, pH, humedad relativa y materia orgánica entre estratos. Tambien se obtuvieron los índices de diversidad de Hill para cada estrato (utilizando las morfoespecies registradas).
El tamaño horizontal de las agregaciones de las morfoespecies principales se estimó utilizando la técnica cuadrante-varianza de dos términos (TTLQ), usando el programa StatEcol (Ludwig y Reynolds 1988), que da como resultado el diametro de las agregaciones. Por último, para relacionar la abundancia de las morfoespecies con los factores ambientales se realizó un análisis de correspondencia canónica (Ter Braak, 1986; 1987). En este análisis, para evitar el exceso de 0 en los registros de morfoespecies, la matriz original fue modificada mediante la transformación de "beals smoothing". El análisis de correspondencia canónica se realizó utilizando el paquete estadístico PC-ORD (versión 2.05, McCune & Mefford, 1995) .
Resultados y discusión
Densidad. La densidad total de la macrofauna fue de 246 ind./m2 (Cuadro 1), que correspondieron a 46 morfoespecies (Cuadro 2). En relación a su abundancia, el 29.9% de los individuos registrados en este estudio correspondió a los Oligochaeta (lombrices de tierra) colectadas principalmente en sus formas juveniles (94.6%), donde el 6% de estos individuos fueron capullos. El segundo en mayor importancia fue el grupo de los Coleoptera (16.9%), representados principalmente por los Tenebrionidae (63% de ellos). Los Miriapoda ocuparon el tercer lugar (10.7%) y de estos, el 62.1% correspondió a los Diplopoda juveniles. Los Isoptera (Termitas) ocuparon solamente el 4.4% del total de la densidad. En total, los grupos restantes como los Hymenoptera, Hemiptera, Lepidoptera y Diptera, entre otros, representaron el 41.2% del total de la macrofauna (Cuadro 2).
La densidad registrada es inferior a la reportada en otros estudios realizados en agroecosistemas tropicales. Lavelle et al. (1981) registraron 1118-2722 ind/m2 en pastizales y selvas de la misma región, durante la época húmeda. En sabanas, pastizales o bosques tropicales, la macrofauna del suelo oscila entre 1830 y 3207 ind/m2 (Lavelle 1983), y en suelos de agroecositemas colombianos se han colectado entre 720 y 1358 ind/m2, durante la época seca (Amat y Soto 1988).
]]> Existen varios factores que pueden ser los causantes de la densidad reducida de la biota edáfica. Uno de estos factores es posiblemente la pérdida de materia orgánica del suelo (Pimentel et al. 1989, Elliot y Lynch 1994) por la exposición al ataque microbiano (Foth 1979) y por la erosión hídrica, acelerada por la pérdida de la cobertura vegetal (Mass y García-Oliva 1990). Lavelle et al. (1981) reportan que el contenido de materia orgánica de los suelos de esta región es particularmente bajo (0.8-2.1%), por lo que consideran que son poco aptos para la explotación agrícola. El contenido de materia orgánica de nuestro suelo (0.53-2.4%) no difiere de los resultados de estos autores. Otro factor que probablemente determine la baja densidad observada es el daño mecánico en la biota de este sitio por efecto de la labranza intensiva a que este suelo está sometido. La variación encontrada en Perú (730-3683 ind./m2) fue explicada con base en este factor. Además, es posible que las condiciones extremas imperantes en el cultivo, dadas por la estacionalidad climática y la textura arenosa del suelo, hallan tenido un efecto drástico sobre la densidad. En general, la densidad de la macrofauna es mayor en ecosistemas naturales que en agroecosistemas. Por ejemplo, en un suelo de Polonia se cree que la fauna de bosques caducifolios disminuyó desde 18 ó 45 gr (peso seco/m2) a 1.5 gr en un periódo de 2000 años debido a la labranza (Ryszkowski 1985).Es posible que debido a sus vínculos con organismos epígeos, la disminución de la fauna del suelo por causas antropogénicas también afecta a aquellos animales no presentes en el edafón (Ryszkowski 1985). En el suelo estudiado es notable la escasez de especies de Coleoptera con un ciclo de vida largo (de meses a varios años) como son los Elateridae y los Melolonthidae. Probablemente su ausencia se debe al efecto conjunto de todos los factores mencionados anteriormente.
Diversidad. Los números de diversidad de Hill (Fig. 1), para cada estrato, muestran que el estrato uno fue el que presentó la mayor riqueza específica (N0), el mayor número de especies abundantes (N1) y muy abundantes (N2).

El desfase existente entre la riqueza específica y el número de especies abundantes y muy abundantes, que es marcado en los primeros estratos, tiende a disminuir conforme se desciende en la profundidad (Fig. 1), y es prácticamente inexistente en los estratos seis y siete. La disminución en los valores de diversidad en los estratos profundos puede deberse a las condiciones extremas registradas en ellos (Cuadro 1), por lo que pocos grupos pueden existir en ellos, y los que lo hacen están en densidades bajas (Cuadro 1). A pesar de ello, se detectaron morfoespecies, como los Oligochaeta juveníles, que tienen una distribución euritópica, pero que tambien tiende a disminuir con la profundidad.
La riqueza específica de la macrofauna de este sitio es baja, lo que posiblemente refleja el estado de degradación del suelo. Este fenómeno se ha visto en lugares como Perú, donde especies presentes en selvas primarias han desaparecido como resultado de la tala y el cultivo (Lavelle y Pashanasi 1989). En este mismo país, un efecto menos dramático ha sido observado en sistemas agrícolas tradicionales o de bajos insumos (Lavelle y Pashanasi 1989).
Se ha argumentado que la riqueza específica contribuye poco a comprender aspectos funcionales del ecosistema y que es una medida redundante cuando especies diferentes comparten la misma función en el suelo (Di Castri y Younes 1990, en Beare et al. 1995). Aún cuando la función puede ser redundante entre organismos del suelo, el conjunto de funciones atribuibles a cualquier especie en particular es única en el funcionamiento del suelo (Beare et al. 1995). Organismos con una función similar a menudo presentan diferentes tolerancias a factores ambientales, y tienen además requerimientos fisiológicos y preferencias de microhabitat distintos (Beare et al. 1995), que dependen a veces de su estado de desarrollo. De esta manera cada estado de desarrollo en cada especie podría estar jugando un papel exclusivo en el sistema edáfico en un momento dado.
Algunos autores han tratado de resolver el problema de la diversidad funcional o taxonómica de la macrofauna del suelo. Por ejemplo, Lavelle y Pashanasi (1989) sugirieren el uso de la categoría "unidad taxonómica". El problema de esta aproximación es que muchos grupos, como los Coleoptera, son sumamente diversos en la fauna del suelo (Villalobos y Lavelle 1990) y la comprensión de sus patrones puede estar determinada por la experiencia taxonómica del investigador. Por otra parte, utilizar como medida de la biodiversidad a la riqueza de especies, en su sentido taxonómico, puede ocultar patrones de diversidad funcional (Villalobos y Lavelle 1990, Villalobos 1991).
Debido a que utilizamos el criterio de "morfoespecie", los resultados de diversidad obtenidos en el presente estudio son difícilmente comparables con los obtenidos en otros trabajos similares. Sin embargo, es necesario considerar que es prácticamente imposible llevar a cabo un trabajo donde se determinen taxonómicamente todas las especies colectadas, sobre todo cuando se toma en cuenta que a nivel mundial el grado de conocimiento taxonómico de muchos grupos de la fauna del suelo es incipiente. Así, el criterio de morfoespecie permitió obtener una medida de la heterogeneidad funcional de la macrofauna del suelo. Entre más fina sea la separación taxonómica de las diferentes entidades biológicas la información obtenida será más certera y dará lugar a mejores hipótesis sobre su ecología.
]]> Distribución espacial de la densidad en el plano vertical. La densidad total de la macrofauna del suelo también disminuye conforme aumenta la profundidad (Cuadro 1). Es notorio que la densidad total en el estrato uno difiere de la observada en los demás estratos. La densidad de los Oligochaeta juveniles se comporta de esta forma, sin embargo, la densidad registrada en el estrato dos fue superior a la registrada en los demás estratos. En los Chilopoda/Geophilomorpha no se detectaron diferencias significativas ya que se distribuyen indistintamente en todos los estratos en bajas densidades. Por su parte las larvas de Tenebrionidae e Isóptera se concentraron exclusivamente en los dos primeros estratos. La distribución vertical de los Diplopoda es semejante a la de las larvas de los Tenebrionidae, pero también se registraron en los estratos cinco y seis (Cuadro 1).A pesar de la mayor temperatura registrada en el primer estrato (Cuadro 1), la diversidad más alta de la macrofauna edáfica se presenta en esta zona del suelo (Fig. 1). Asímismo, es interesante notar que la diversidad en los estratos tres y cuatro fue menor que la observada en varios de los estratos inferiores (Fig. 1), aún cuando la temperatura y pH no variaron significativamente (Cuadro 1). Se observo que el suelo estudiado presenta una alta compactación en los estratos tres y cuatro. Tal vez por ello el porcentaje de materia orgánica contenida en el suelo es significativamente menor después del estrato tres (Cuadro 1). La compactación puede ser debida a que esta zona no es perturbada por la labranza y a que en ella se depositan minerales o materiales compactantes. Estas últimas partículas son posiblemente transportadas desde los estratos superficiales hasta los profundos por lixiviación (Dubroeucq et al. 1992) durante la época de lluvias. El transporte en la dirección opuesta también es factible (Foth 1979), cuando los minerales presentes en estratos profundos son solubilizados y transportados hacia la superficie, al elevarse el nivel del agua del manto fréatico. Este patrón puede explicar la distribución bimodal que se observa en la diversidad (Fig. 1). La compactación del suelo en esos estratos puede producir un efecto de frontera para muchos grupos de invertebrados. Por ejemplo, es posible que los Diplopoda juveniles, las larvas de Tenebrionidae y los Isóptera sean incapaces de atravesar esta barrera, como lo sugieren los datos del Cuadro 1. La forma de los invertebrados o su modo de desplazamiento puede jugar un papel importante en su capacidad de trasladarse entre diferentes estratos. Por ejemplo, los Geophilomorpha podrían usar para su desplazamiento, vertical y horizontal, las pequeñas galerías que forman los Oligochaeta. Estas galerias fueron observadas en estratos profundos. Esta situación probablemente repercute en la distribución uniforme de los Geophilomorpha en ambos planos.
De acuerdo con Ryszkowski (1985) la actividad agrícola afecta las condiciones ambientales del suelo y muchos grupos de la macrofauna son particularmente sensibles a estos cambios a largo plazo. Este autor señala que los Mollusca y Miriapoda, entre otros grupos, han sido prácticamente erradicados de la fauna del suelo en agroecosistemas de Polonia.
Creemos que es necesario un estudio más detallado sobre la estratificación de las variables ambientales en el perfil del suelo, así como de su influencia en la macrofauna, para detectar el efecto del ambiente en la distribución vertical de estos organismos.
Distribución espacial en el plano horizontal. Para medir el impacto de especies con incidencia directa en actividades benéficas o perjudiciales para los cultivos, es importante conocer el tamaño de las agregaciones de los invertebrados del suelo. La distribución horizontal y diametro de las agregaciones de las cinco morfoespecies más abundantes en este estudio se muestran en la Fig. 2. Como se puede ver, los Oligochaeta juveniles se distribuyeron de manera agregada y el tamaño de su agregación fue superior a 1.5 m. Por lo tanto, para este grupo no se logró detectar el tamaño de su agregación aún considerando el área máxima analizada en este estudio. Una situación similar se presentó con los Diplopoda pero no con los Chilopoda. Estos últimos mostraron una distribución uniforme. Los grupos en los que el tamaño de las agregaciones pudieron ser calculados fueron las larvas de Tenebrionidae, cuyas agregaciones (varianza máxima) fueron de 0.9 m, y los Isóptera en los que la agregación fue de 1.2 m (Fig. 2).

Los diferentes grupos de la macrofauna del suelo estudiado tienen estrategias distintas en cuanto a su disposición espacial. Factores como la disponibilidad de recursos alimenticios, condiciones microclimáticas, propiedades químicas y estructura del suelo pueden afectar significativamente el tamaño, la composición y la distribución de las comunidades bióticas existentes en el (Beare et al. 1995). Conocer a que factores responde la distribución espacial de cada una de las morfoespecies registradas es importante, ya que permitiría estimar más precisamente su densidad. Las repercusiones de estos patrones, sobre todo en sus efectos sobre las plantas, pueden ser analizadas a la luz de las transformaciones biogénicas promovidas por la macrofauna (Beare et al. 1995). Por ello, sugerimos estimar otros factores ambientales en todos los estratos del suelo. Conociendo la distribución de estos posiblemente se podría comprender mejor el patrón de morfoespecies particulares.
Es conveniente señalar que un muestreo al estilo Cuadrante-varianza es útil para detectar patrones de distribución horizontal de la fauna del suelo donde el patrón no es fácilmente observable y se requiere medirlo. Sugerimos que este tipo de muestreo debe usarse cuando se intente responder a preguntas específicas sobre la distribución espacial de la fauna edáfica y cuando se requiera de estimaciones más acertadas de la densidad de especies clave. Este método ha sido utilizado principalmente para la descripción de patrones de especies vegetales (Ludwig 1979, Heltshe y Ritchey 1984) pero no se conocen intentos previos aplicados al estudio de la fauna del suelo.
Pensamos que el uso de este tipo de muestreo no es conveniente cuando se pretende conocer los patrones de la fauna del suelo a una escala de paisaje. Además, es importante tomar en cuenta que su realización requiere de un esfuerzo de muestreo considerable.
]]> Variables ambientales y patrones de distribución de la macrofauna. Los suelos tropicales presentan, por lo general, temperaturas altas y regímenes hídricos contrastantes (acordes a la duración de la época seca, capacidad del suelo para retener agua y cobertura vegetal) (Lavelle 1983). Estas condiciones tienden a ser extremas en la medida en que el suelo queda expuesto por la falta de cobertura vegetal a las condiciones del medio.En el suelo estudiado la temperatura, el pH y el porcentaje de materia orgánica disminuyen al aumentar la profundidad, mientras que la humedad relativa se incrementa (Cuadro 1). El estrato uno presenta, para las cuatro variables, valores significativamente diferentes a los estratos dos al siete.
Los resultados obtenidos con relación al pH difieren con los reportados para el CICOLMA por Dubroeucq et al. (1992). Estos autores encuentran un pH más alcalino conforme se desciende en el suelo. Estas diferencias pueden ser atribuibles a las distintas técnicas utilizadas para estimar este parámetro.
En el sitio de estudio la humedad para el primer estrato es baja (9.5%) respecto a la registrada en un pastizal ubicado a 100 Km. al noroeste, donde, este estrato alcanza valores mínimos de 12%, pero superiores al punto de marchitamiento permanente (11.7%), durante la misma época (Villalobos 1991). Esto sugiere que en el suelo estudiado, a pesar de que la humedad puede ser una limitante para las plantas, algunas especies de la fauna del suelo han desarrollado mecanismos para soportar las condiciones extremas de sequia.
La agrupación de morfoespecies con preferencias hacia uno o más factores ambientales es una herramienta valiosa para la búsqueda de especies bioindicadoras y permite generar hipótesis sobre su biología. La Fig. 3 muestra la ordenación de las morfoespecies y su relación con los factores ambientales después de un análisis de correspondencia canónica. Las correlaciones entre las abundancia de las morfoespecies y las variables ambientales para los dos ejes fueron 0.98 y 0.88 respectivamente. Las correlaciones inter-set entre el eje 1 y los factores ambientales fueron: 0.9 con el pH, -0.85 con la humedad, 0.73 con la temperatura y 0.7 con la materia orgánica. La correlación ínter-set entre el eje 2 y los factores ambientales fue baja, siendo la temperatura la más alta (r = 0.56).
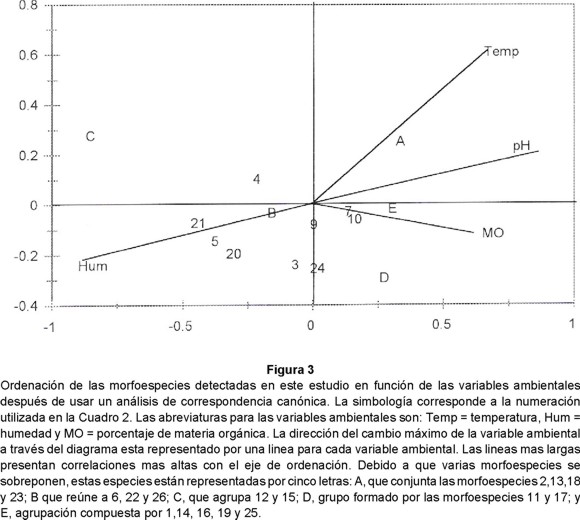
La abundancia de las morfoespecies detectadas en este estudio pueden ser utilizada para clasificarlas en cinco grupos (Fig. 3), dependiendo del o los factores ambientales que están más relacionados con su distribución de abundancias: A) aquellas relacionadas con el eje 1 y la materia orgánica, por ejemplo, Arachnidae y los juveniles de Diplopoda e Isoptera; B) las influenciadas por el eje 2, materia orgánica y humedad, como Hemiptera y las larvas de Aphodiinae y Scarabaeinae; C) las relacionadas por el eje 2 y la humedad, tales como los Geophilomorpha sp. 1, Chilopoda spp. 2 y 3 y Mermithidae; D) las determinadas por el eje 1 y la temperatura, como larvas de diptera y tenebrionidae; y E) las relacionadas con el eje 1 y la humedad, entre las que están las larvas y adultos de carabidae. En la Fig. 3 puede observarse la distribución de las morfoespecies considerando los factores ambientales y en el Cuadro 2 su actividad trófica predominante.
A pesar del conocimiento limitado de los hábitos alimenticios de la fauna del suelo, la clasificación utilizada (Cuadro 2) nos permite estimar los porcentajes de la fauna del suelo que participan en cada eslabón del flujo de materia y energía en el suelo (Ryszkowski 1985). Por ejemplo, de los grupos abundantes de la macrofauna colectados, sólo los Chilopoda/Geophilomorpha se consideran preponderantemente depredadores (Bueno, com. pers) y no participan en los primeros eslabones del reciclaje de la materia orgánica del suelo.
Por otra parte, la dominancia de los organismos saprófagos (Cuadro 2) coincide con lo observado en un campo de cultivo de trigo en Polonia (Ryszkowski 1985). Posiblemente en ambos sitios la dominancia de Oligochaeta y de larvas de insectos exopterigótos, determinen la estructura del suelo. Su dominancia probablemente también está determinada por la baja cantidad de raíces vivas observada, así como por una incorporación reducida de la materia vegetal proveniente del cultivo.
]]> En general, debido a la abundancia reducida de la macrofauna de este suelo y a las condiciones imperantes en el sustrato, es posible que estemos observando el resultado de efectos acumulados a largo plazo.
Conclusiones
La densidad de la macrofauna del suelo estudiado es notablemente baja en comparación a la observada en otros agroecosistemas del mundo. La mayor parte de las morfoespecies colectadas se distribuyen preferentemente en el primer o segundo estrato del suelo y tienen una distribución agregada. Se distinguen, sin embargo, grupos con una distribución uniforme tanto en el plano vertical como horizontal (el caso de los Chilopoda/Geophilomorpha).
A pesar de que el perfil del suelo estudiado presenta una notable variación en temperatura, pH, humedad y materia orgánica, pudieron distinguirse grupos de la macrofauna edáfica que parecen estar determinados por sus preferencias y rangos de tolerancia a estos factores. Consideramos que los patrones de distribución de la fauna del suelo son un reflejo de las preferencias de esta biota a diferentes microambientes.
Agradecimientos
Deseamos agradecer a M. Rodríguez, J. Galindo, C. Alvarez, A. Arango y A. Capistran por su ayuda en campo, y a C. Fragoso por sus valiosos comentarios a una versión inicial de este trabajo.
Literatura citada
]]>Amat, G. & L. Soto. 1988. Efectos del uso actual del suelo sobre la macrofauna edáfica en la región del Guejar, Reserva Natural Integral La Macarena. Meta 2(6-7):43-67. [ Links ]
Anderson, J.M. & J.S.I. Ingram, 1993. Tropical Soil Biology and Fertility. CAB International. [ Links ]
Beare, M.H., D.C. Coleman, D.A. Crossley, P.F. Hendrix & E.P. Odum. 1995. A hierarchical approach to evaluating the significance of soil biodiversity to biogeochemical cycling. Pp. 5-22. In: Collins, H.P., G.P. Robertson and M.J. Klug (Eds.). The Significance and Regulation of Soil Biodiversity. Kluwer Academic Publisher, Netherlands. [ Links ]
Blain, D. 1988. Factors affecting the early stages of regeneration of three tropical tree species in a seasonal forest, Veracruz, Mexico. Thesis MsC. York University, North York, Ontario. 155 pp. [ Links ]
Borror, D.J., D.M. De Long & C.A. Triplehorn. 1981. An introduction to the study of insects. 5th ed. Saunders College Publishing. 827 pp. [ Links ]
]]>Dubroeucq, D., D. Geissert, P. Moreno & G. Millot. 1992. Soil evolution and plant communities in coastal dunes near Veracruz, Mexico. Cah. Orstom, ser. P. dol. 27(2):237-250. [ Links ]
Elliot, L.F. & J.M. Lynch. 1994. Biodiversity and soil resilience. Pp. 352-363. In: Greenland, D.J. and Szabolcs, (Eds.) Soil Resilence and Sustainable Land Use. CAB International. [ Links ]
Foth, H.D. 1979. Fundamentals of Soil Science. 7th ed. John Wiley & Sons, NY, USA. 435 pp [ Links ]
Fraser, P.M. 1994. The impact of soil and crop management practices on soil macrofauna. Pp. 125-132. In: Pankhust, C.E., B. M. Doube, V.V.S.R. Gupta and P.R. Grace, (Eds.) Soil Biota. Management in Sustainable Farming Systems. C.S.I.R.O, Australia. [ Links ]
Gonzalez, O.R. & M.A. Herrera. 1983. La macrofauna del suelo de una plantación de majagua (Hibiscus elatus SW) en Cuba: Resultados preliminares. Poeyana 258:1-16. [ Links ]
Heltsche, J.F. & T.A. Ritchey 1984. Spatial pattern detection using quadrat samples. Biometrics 40:877-885. [ Links ]
Lavelle, P. 1983. The soil of tropical savannas. I The community structure. Pp. 477-484. In: Goodall, D.W. and F. Bourliere, (Eds.) Tropical Savannas. (Ecosystems of the world, 13.) Elsevier, Amsterdam. [ Links ]
Lavelle, P., M. Dangerfield, C. Fragoso, V. Eschenbrenner, D. López-Hernández, B. Pashanasi & L. Brussaard, 1994. The relationship between soil macrofauna and tropical soil fertility. Pp. 137-169. In: Woomer, P.L. and M.J. Swift, (Eds.) The Biological Management of Tropical Soil Fertility. TSBF. A Wiley-Sacey Publication. [ Links ]
Lavelle, P. & B. Kohlmann. 1984. Etude quantitative de la macrofaune du sol dans une forét tropicale humide du Mexique (Bonampak, Chiapas). Pedobiologia 27:377-393. [ Links ]
Lavelle, P., M.E. Maury & V. Serrano. 1981. Estudio cuantitativo de la fauna del suelo de la región de Laguna Verde, Ver. Epoca de lluvias. Pp. 65-100. In: Reyes-Castillo, P.(Ed.) Estudios ecológicos en el trópico mexicano. Instituto de Ecología, México, D.F. [ Links ]
Lavelle, P. & B. Pashanasi. 1989. Soil macrofauna and land management in Peruvian Amazonia (Yurimaguas, Loreto). Pedobiologia 33:283-291. [ Links ]
Ludwig, J.A. 1979. A test of different quadrant variance methods for the analysis of spatial patterns, Pp. 289-303. In: Cormack, R.M. and J.K. Ork, (Eds.) Spatial and temporal Analysis in Ecology, International Co-operative Publishing House, Fairland, Maryland. [ Links ]
Ludwig, J.A. & J.F. Reynolds. 1988. Statistical Ecology. John Wiley & Sons, New York. 329 pp. [ Links ]
Mass, J.M. & F. García-Oliva. 1990. La conservación de suelos en zonas tropicales: el caso de México. Ciencia y Desarrollo. 15(90):21-36. [ Links ]
McCune, B. & M.J. Mefford. 1995. PC-ORD, multivariate analysis of ecological data. Version 2.05. MjM sofware, Gleneden Beach, Oregon, USA. [ Links ]
Moreno-Casasola, P. 1982. Ecología de la vegetación de dunas costeras. Biotica 7(4):577-602. [ Links ]
Ortiz-Pulido, R., H. Goméz, F. González-García & A. Alvarez. 1995. Avifauna del Centro de Investigaciones Costeras La Mancha, Veracruz, México. Acta Zool. Mex. (n.s.) 66:87-118. [ Links ]
Pimentel, D., T.W. Cullinery, I.W. Buttler, D.J. Reinemann & K.B. Beckman. 1989. Low-input sustainable agriculture using ecological management practices. Agr. Ecosyst. Env. 27:3-24. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Ed. Limusa. 432 pp. [ Links ]
Ryszkowski, L.1985. Impoverishment of soil fauna due to agriculture. Intecol Bulletin 12:7-17. [ Links ]
Ter Braak, C. 1986. Canonical correspondence analisys: a new eigenvector technique for multivariate direct gradient analysis. Ecology 67: 1167-1179. [ Links ]
––––––––––. 1987. The analisys of vegetation-environment relationships by canonical correspondence analysis. Vegetatio 68: 69-77. [ Links ]
Villalobos, F.J. 1991. The community structure of soil Coleoptera (Melolonthidae) from a tropical grassland in Veracruz, Mexico. Pedobiologia 35:225-238. [ Links ]
Villalobos, F.J, & P. Lavelle. 1990. The soil coleoptera community of a tropical grassland from Laguna Verde, Veracruz, (Mexico). Rev. Ecol. Biol. Sol. 27(1):73-93. [ Links ]
Zar, J.H. 1984. Biostatistical Analysis. Prentice-Hall, New Jersey. 929 pp. [ Links ]
]]>