
El mundo feliz de las hormigas
The happy world of the ants
Germán Octavio López-Riquelme y Fidel Ramón*
Lab. de Socioneurobiología, División de Investigación, Facultad de Medicina, UNAM. Edificio de Investigación, 6° piso. Avenida Universidad No. 3000, Col. Copilco, C.P. 04510, México, D.F. *E-mail: fidelrr@unam.mx
]]>Artículo recibido el 19 de abril de 2010.
Aceptado el 24 de mayo de 2010.
Resumen
La exitosa vida de las hormigas ha evolucionado a partir de la división del trabajo reproductivo entre individuos fértiles (reinas) y estériles (obreras) y del trabajo de manutención de diferentes castas de obreras. Este éxito representa un problema al tratar de explicar su enorme desarrollo social y sobrevivencia a los desastres naturales. La división del trabajo y sus interacciones a nivel colonial hace a las hormigas estupendos modelos para estudios anatómicos, embriológicos, genéticos, fisiológicos y moleculares. Debido a la división del trabajo, su comportamiento, instinto e inteligencia, representan, además, un problema difícil de entender. Así, por sus adaptaciones conductuales especie-específicas, las hormigas constituyen excelentes modelos neurobiológicos para estudiar tanto su sistema sensorial sofisticado y especializado en la detección de señales comunicativas de naturaleza química, como las causas y la estructura del comportamiento debido a su organización social. Uno de los principales problemas para los investigadores es establecer los mecanismos mediante los cuales los individuos colaboran en el complejo comportamiento de la colonia. Estas propiedades tienen como sustrato físico el cerebro y es en este diminuto órgano donde el comportamiento complejo es generado y controlado. No obstante, no es el comportamiento de sus individuos lo que hace a las hormigas seres dominantes en la biosfera, sino el comportamiento general de la colonia. Las capacidades individuales han evolucionado dentro de un contexto social y es bajo esas condiciones que debe ser evaluado para entender cómo la expresión génica y la función cerebral son sensibles a ese contexto. Ningún otro grupo de animales presenta tal cantidad de problemas fascinantes para los biólogos, psicólogos, sociólogos, físicos y matemáticos. En este artículo haremos una breve revisión de la organización social de las hormigas y las bases de cómo el sistema nervioso genera y regula los complejos patrones de comportamiento social.
Palabras Clave: Cerebros de hormigas, división del trabajo, eusociedades, evolución social, hormigas, olfato, organización social, selección por parentesco, sistema nervioso, sociogenómica, socioneurobiología.
Abstract
The successful life of ants has evolved due to the division of reproductive labor among fertile (queens) and sterile individuals (workers), and to the support labor by different kinds of workers. Because of this and their age on Earth, ants represent an evolutionary issue in relation to their ability for social development and to survive natural catastrophes. The division of labor and colonial level phenomena make ants excellent neurobiological models to study the structure and the basis of behavior of their social organization and division of labor; also their species-specific behavioral adaptations and their sophisticated sensorial system specialized in the detection of chemical signals. One of the main issues for researchers is to elucidate the mechanisms by which individuals cooperate to the whole colony behavior. These properties have as physical substrate the brain, and it is in this minute organ that complex behaviors are generated and controlled. However, individual behavior is not what make ants dominant creatures in the biosphere, instead, the systemic behavior of the colony as a whole. Individual abilities have evolved in a social context, and no other animal group generates so many fascinating issues for biologists, psychologists, sociologists, physicists and mathematicians. In this paper, we briefly review what it is known about the social organization of ants and how the nervous system generate the complex patterns of social behavior.
]]> Key Words: Ant brain, division of labor, eusocieties, social evolution, ants, olfaction, social organization, kin selection, nervous system, sociogenomics, socioneurobiology.
Introducción
Como todos aquellos interesados en las hormigas, apoyados sobre las manos y las rodillas hemos observado, hurgado con ramitas y excavado nidos en el campo, en los jardines de la ciudad e incluso en las áreas naturales de Ciudad Universitaria. En repetidas ocasiones hemos terminado aguijoneados o con decenas de mordidas, las cuales, durante días nos provocan comezón en todo el cuerpo, por lo que no es raro vernos rascándonos por los pasillos del edificio en donde realizamos nuestra investigación.
¿Por qué admiramos tanto a las hormigas?, ¿por qué tomarse todas esas molestias y armar un laboratorio para estudiar estos minúsculos insectos? Cuando vemos una hormiga frente a nosotros, nos preguntamos cosas como: ¿a dónde va?, ¿cómo sabe a donde ir?, ¿cómo sabe regresar a su casa?, ¿cómo sabe qué, cuándo y dónde hacer lo que hace?, ¿cómo sabe quiénes son los miembros de su colonia y quiénes sus enemigos? y un largo etcétera. Evidentemente es mucho más fácil no preguntarse nada y aplastar la hormiga con el dedo, pero nosotros hemos preferido el difícil camino de tratar de entender cómo es su mundo.
Pero no se nos malinterprete, el estudio de las hormigas no es un asunto trivial o meramente de puro placer. El estudio de las hormigas, así como el de otros insectos sociales, es fundamentalmente el estudio de los niveles de organización y de selección, por lo que es importante para comprender el funcionamiento y evolución de sistemas biológicos complejos en los que están involucrados desde los genes hasta la sociedad. Los genes producen proteínas que construyen sistemas neuroendocrinos y musculares que interactúan regulando el comportamiento de los individuos. Después, las complejas interacciones de los individuos producen el comportamiento social y para lograr entender cómo funciona y evoluciona esa organización, es necesario entender cómo interactúan sus diferentes niveles; es decir, cómo están vinculados los genes con el desarrollo y funcionamiento del sistema nervioso individual y éste con la organización social1.
En este artículo queremos mostrar un poco de ese mundo haciendo una corta revisión de lo que actualmente sabemos sobre la organización social de las hormigas, la relevancia de ese conocimiento y la importancia de nuestro trabajo en la enorme empresa de entender el funcionamiento de esos complejos sistemas que son las sociedades de hormigas.
La vida en sociedad: una historia de éxito ecológico
Las hormigas son excepcionales entre los insectos por su dominancia como depredadores, excavadores y herbívoros, y su historia evolutiva quizá sea la de mayor éxito entre los metazoarios terrestres. Aunque las más de 12,000 especies de hormigas constituyen aproximadamente el 2% de todas las especies de insectos, forman al menos un tercio de su biomasa, y en términos globales representan un tercio de la biomasa animal total. Se ha estimado que en el mundo existen diez mil billones de hormigas, las que pesarían aproximadamente lo mismo que todos los seres humanos2.
]]> Las hormigas se encuentran en todos lados, incluso en los ambientes más perturbados y sólo están ausentes de las regiones con glaciares permanentes y los cuerpos de agua. Debido a su abundancia ejercen una influencia importante en muchos hábitats, controlando el crecimiento poblacional de otros artrópodos, removiendo y aireando grandes cantidades de suelo en bosques y praderas y haciendo circular nutrientes esenciales para otras formas de vida.
¿A qué se debe que las hormigas sean tan abundantes, tan exitosas y que dominen sobre la Tierra? No hay lugar a dudas, su éxito se origina a partir de la cooperación y organización social basada en la división del trabajo. Así, mientras uno o varios individuos se encargan de la reproducción, los otros cuidan a las crías, consiguen alimento, protegen al nido y construyen el hogar4.
¿Quién habita la colonia?
Todas las hormigas viven en enormes familias constituidas por una o varias madres (llamadas reinas) y su progenie de hembras estériles (llamadas obreras). Entre ellas no hay egoísmo, todo lo que hacen es por la colonia, de manera que cada individuo lleva sus presas a casa para compartirlas con las demás. La reina es una enorme hormiga que vive enclaustrada en la parte menos accesible y más segura del nido, y sólo se dedica a poner huevos que puede fecundar o no, ya que tiene control sobre la abertura y cierre del órgano donde almacena los espermatozoides. Si los huevos son fecundados resultan en hembras diploides que pueden desarrollarse en obreras estériles o en nuevas reinas aladas según las condiciones ambientales y la alimentación que reciban. Por el contrario, un huevo no fecundado produce un macho alado haploide que se desarrolla por partenogénesis.

Vida y muerte de una colonia
El nacimiento de una colonia comienza con el vuelo nupcial. Durante la época reproductora, generalmente cuando el clima es más favorable, las colonias maduras liberan miles de reinas vírgenes y machos que salen volando en busca de parejas de otras colonias para aparearse. Las reinas pueden aparearse con uno o varios machos y almacenar desde 7 hasta 300 millones de espermatozoides en una bolsa llamada espermateca. Ahí, los espermatozoides se mezclan y permanecen en estado suspendido toda la vida de la reina, que en algunas especies puede llegar ¡hasta los 20 años!
Después del apareamiento, las reinas continúan brevemente con su primer y único vuelo para, posteriormente, aterrizar en sitios cuidadosamente seleccionados. Una vez en el suelo, se arrancan las alas y excavan un refugio para iniciar su propia colonia y no salir jamás. Con las reservas grasas en su cuerpo y las proteicas de los músculos alares que no volverán a usar, las reinas ponen huevos, alimentan a las larvas y crían a su primera generación de obreras. De cada huevo eclosiona una pequeña larva en forma de gusano que es cuidadosamente alimentada y limpiada durante algunas semanas hasta que inicia la metamorfosis, para la cual, se encierra en un capullo que teje con la seda de sus glándulas labiales (aunque hay especies que no tejen capullo). Cuando ha terminado la pupación, el capullo es retirado, ya sea por la reina o por sus hermanas mayores y la joven hormiga comienza su vida de trabajo en la colonia.
]]> Una vez que la segunda generación de obreras ha sido criada, las colonias entran a un período de rápido crecimiento en el que sólo se producen obreras. Los recursos son reinvertidos en obreras del tipo más eficiente y en infraestructura y no se produce ninguna forma reproductiva. La reina interrumpe cualquier actividad y se consagra a la oviposición, mientras que las obreras realizan todas las tareas. La sociedad se cierra y, con pocas excepciones, nunca se adoptan individuos del exterior.Al llegar a cierto tamaño crítico, que depende de la especie, las colonias comienzan a producir descendencia sexual, lo que implica el desvío de recursos de la producción de obreras hacia la crianza de machos y reinas. El tiempo y la cantidad de descendencia sexuada producida resultan críticos para el futuro de la colonia. Una producción que ocurre pronto puede representar la ventaja de que la descendencia funde nuevas colonias rápidamente. Sin embargo, esto también puede reducir el número total de descendientes que una colonia puede producir durante toda su vida.
¿Qué pasa cuando la reina muere por vejez o por algún accidente? Algunas colonias tienen múltiples reinas y continuamente producen nuevas que se aparean dentro del nido con parientes cercanos. En estas colonias las reinas que mueren son sustituidas por las nuevas, de manera que estas sociedades son casi inmortales. Sin embargo, en la mayoría de los casos, cuando la reina muere, la colonia perece lentamente al dejar de producir obreras. Sin embargo, las obreras, aunque con ovarios reducidos, pueden poner huevos sin fecundar de los que se desarrollan machos. Gracias a estas obreras, la colonia puede contribuir genéticamente a las siguientes generaciones, aún cuando la reina haya perecido.
Sociedades y eusociedades
Pero, ¿qué es una sociedad? Los biólogos definen una sociedad como un grupo de individuos de la misma especie, organizados de manera cooperativa mediante un sistema eficiente de comunicación recíproca que buscan un objetivo común5. Muchos animales viven en sociedades que pueden ser simples o complejas, la mayoría de las cuales ha evolucionado a partir de unidades familiares y el cuidado de la descendencia.
Las hormigas por supuesto que también forman sociedades, pero éstas, como las de los otros insectos sociales, tienen características particulares y extremas que han hecho que los biólogos definan un tipo muy específico de sociedades: las eusociedades. Literalmente la palabra significa sociedad verdadera, pero los biólogos usan el término para describir una "sociedad avanzada", la cual está caracterizada porque sus miembros sacrifican su capacidad reproductiva en favor de la de otros. Una eusociedad tiene cuatro características fundamentales: 1) división del trabajo reproductivo; 2) cooperación en el cuidado de las crías (la descendencia ayuda a los progenitores durante su vida); 3) varias generaciones de adultos que ayudan al cuidado de la descendencia y 4) altruismo.
Además de la división del trabajo reproductivo entre reinas y obreras, las tareas de mantenimiento se reparten entre grupos de obreras más o menos especializadas. La especialización principal se encuentra entre obreras que se dedican a actividades dentro del nido y aquéllas que las realizan en el exterior, pero aún dentro de estas categorías puede haber especialización. Estas obreras realizan las tareas necesarias actuando coordinadamente a través de un complejo sistema de comunicación quimiotáctil.
Actualmente sabemos que muy pocos grupos animales forman eusociedades: 1) los insectos sociales, que incluyen todas las hormigas, muchas especies de abejas y avispas y todas las termitas6; 2) los camarones sociales del género Synalpheus que habitan esponjas en arrecifes de coral7 y 3) los únicos vertebrados eusociales son la rata-topo desnuda (Heterocephalus glaber) y la rata-topo Damara (Cryptomys damarensis), que viven en colonias subterráneas en la parte oriental de África8.
]]> Los insectos sociales, ¿un reto para la teoría de la evolución?
Cuando Darwin escribió El origen de las especies, tenía muy clara la idea que deseaba transmitir. Darwin deseaba convencer al lector y su argumentación resulta muy persuasiva. Además, empleó una estrategia deseable en todo científico: la autocrítica. Se planteó objeciones que suponía el lector podría tener y las resolvió con destreza. La principal de dichas objeciones y que él mismo llamó "una dificultad especial que al principio me pareció insuperable y fatal para toda la teoría", fueron los insectos sociales y la presencia de individuos estériles en sus colonias. ¿Cómo -se preguntaba Darwin- podrían propagar sus caracteres estos individuos si no se reproducen?
Aunque Darwin ya había intuido que el parentesco jugaba un papel importante en la evolución de castas estériles en los insectos sociales, no fue hasta 1955 que J. B. S. Haldane9 propuso una solución que parecía explicar la evolución del altruismo y que expresaba en su famosa frase: "...me arrojaría voluntariamente al agua para evitar que se ahogaran tres de mis hermanos o nueve de mis primos hermanos...". La idea general es muy simple: el altruismo puede evolucionar si por medio de un acto de sacrificio el altruista incrementa el éxito reproductivo de sus parientes. Sin embargo, no fue sino hasta 1964 que W. D. Hamilton10 desarrolló de manera completa la teoría de la selección por parentesco que explicaba la evolución del altruismo en los himenópteros sociales. La explicación estaba basada en la selección a nivel de genes. Hamilton reconoció que existen dos formas en las que los alelos pueden pasar a generaciones futuras. La primera es a través de la reproducción individual produciendo hijos e hijas y es conocida como aptitud. La segunda es colateral, promoviendo el bienestar de hermanos, hermanas y otros parientes y el de su descendencia, la cual presenta un porcentaje de los mismos alelos debido a la ascendencia común. Esta medida la llamó aptitud inclusiva e incorpora tanto la reproducción individual como su influencia en la reproducción de parientes colaterales: Aptitud inclusiva = Aptitud individual + Aptitud de parientes cercanos. Por lo tanto, cuando un animal realiza un acto altruista hacia un hermano, la aptitud inclusiva es la del animal (que ha disminuido a raíz del acto) más el aumento en aptitud por aquella porción de genes en común con el hermano.
Para Hamilton, la determinación haplodiploide del sexo de los himenópteros era fundamental para la evolución de sociedades en las que sus miembros sacrifican su reproducción en beneficio de la de otros. Las hembras (reinas y obreras) de las hormigas se desarrollan de huevos fecundados (diploides) con un juego de cromosomas de la madre y uno del padre. Los machos se desarrollan de huevos no fecundados (son haploides), con sólo un juego de genes provenientes de la madre. No hay reducción del número de cromosomas durante la espermatogénesis11. Como resultado de este tipo de determinación del sexo todas las hermanas hijas de la misma madre y del mismo padre tienen en común la mitad de los genes de la madre y todos los genes de su padre. Puesto que la mitad de su componente genético viene de cada padre, la fracción promedio de genes que las hermanas tendrán en común está dada por: (1 x 1/2) + (1/2 x 1/2) = 3/4. Así, las hembras comparten más genes entre ellas (75%) que los que comparten con sus hermanos (25%) o con su madre (50%), o que los que compartirían con sus hijas (50%) por lo que, según este razonamiento, la aptitud de las obreras depende mayormente de la fecundidad de sus hermanas fértiles que de la suya propia, lo cual, según Hamilton (1964)10, ha favorecido la evolución del altruismo.
No obstante que la teoría de la selección por parentesco parece explicar sencillamente la evolución de las sociedades de insectos, sólo es válida si la reina es fecundada por un solo macho. Pero hay algo más, también depende de la proporción de sexos.
No existe sociedad perfecta: todos tienen intereses propios
Robert L. Trivers12,13 advirtió que el argumento de Hamilton tiene una dificultad relacionada con la proporción de sexos. Para las reinas, la proporción de sexos óptima es 1:1, ya que cada sexo tiene la misma cantidad de genes en común con ella, es decir, 50%. Si ésta fuera la proporción de sexos producida en una colonia, las obreras no obtendrían beneficios de su sacrificio reproductivo. La razón es la siguiente relación aritmética: si la proporción de sexos es 1:1, entonces el 50% de la producción corresponde a machos y el otro 50% a hembras, así, una mitad de los machos con 1/4 de genes en común con sus hermanas, más una mitad de hembras con 3/4 de genes en común con sus hermanas, resulta en un medio de genes en común con los sexuados producidos por la colonia completa, es decir, (3/4 x 1/2) + (1/4 x 1/2) = 1/2, la misma proporción de genes que si las obreras se reprodujeran solas, por lo que el sacrificio reproductivo no tendría sentido. La única manera en que las obreras tengan beneficios a partir de la selección por parentesco, es que aumenten la proporción de hermanas hembras fértiles hasta 3:1y eso precisamente es lo que ocurre. Aunque aún no sabemos cómo hacen las obreras para lograr producir más hermanas que hermanos, los datos sugieren que pueden reconocer el sexo de la cría14.
No obstante, la reina, a través del apareamiento con varios machos, puede manipular y minimizar el conflicto con sus obreras, pues reduce el parentesco entre ellas, por lo que logra que las obreras críen sus huevos machos. Otra forma de reducir el conflicto entre reinas y obreras es incrementar el número de reinas en una colonia (fenómeno llamado poliginia), con lo que aumenta la heterogeneidad genética intracolonial. Sin embargo, se ha encontrado que en la mayoría de los casos de poliginia, la sociedad es fundada por una asociación de reinas emparentadas14. Por si fuera poco, la reina puede actuar químicamente sobre sus obreras y evitar que ovipositen al inhibir la función de sus ovarios.
Tomando en cuenta todo lo anterior, actualmente se considera que, si bien es cierto que el parentesco podría ser una vía para la evolución de eusociedades y que el parentesco puede determinar numerosos detalles de la vida social15, la teoría de Hamilton podría estar incompleta. Primero, porque Hamilton consideró solamente a los himenópteros eusociales con sistema haplodiploide de determinación del sexo. Sin embargo, no consideró a los escarabajos de ambrosia que también presentan haplodiploidía pero no forman eusociedades. Segundo, tampoco consideró que las termitas no son haplodiploides pero sí forman eusociedades. Ahora conocemos otros animales eusociales que no son haplodiploides como los camarones sociales y las ratas-topo. Podemos decir, entonces, que la selección por parentesco podría, pero no necesariamente, favorecer la evolución de sociedades altruistas. Una fuerza más importante sería la selección natural actuando sobre la colonia de acuerdo con las presiones y las oportunidades que ofrece el ambiente. Las colonias así formadas, estarían favorecidas por su capacidad para crear y defender sus nidos16.
]]>División del trabajo
La división del trabajo es un tópico fundamental en la biología de los insectos sociales, pero también en la biología en general, ya que muchos de los principios involucrados se aplican a todos los sistemas biológicos17. La división del trabajo está caracterizada porque grupos de individuos especializados realizan diferentes actividades. Es la única forma de cooperar y funcionar como unidad, en lugar de la acción descoordinada de miembros no especializados que sólo se estorbarían durante la realización de una actividad común18.
La división del trabajo optimiza el consumo de energía en relación con la producción debido, primero, a la repartición de tareas19; segundo, a que los especialistas enfocan su actividad en las partes del nido donde es más probable que encuentren estímulos relacionados con su tarea; tercero, a que la ejecución de tareas relacionadas requiere de habilidades sensoriales y motoras similares; y, cuarto, porque la realización de una tarea mejora con la experiencia6. La determinación de las castas en los insectos sociales es principalmente mediada por el ambiente, de donde provienen señales endocrinas y nutricionales que actúan como factores próximos que "conducen" el desarrollo de los individuos. Existen diferentes sistemas de división del trabajo: el polietismo temporal y el polimorfismo.
Polietismo temporal: repartición de las tareas según la edad
El polietismo temporal está basado en la transición de tareas conforme las obreras envejecen, empezando con tareas de servicio interno como el cuidado de la cría, y avanzando progresivamente hacia el mantenimiento del nido hasta alcanzar las actividades externas, como el forrajeo. A lo largo de la ontogenia las obreras sufren cambios fisiológicos internos que provocan que su respuesta a los estímulos externos varíe19,20.
Aunque en las hormigas todavía no se han obtenido datos suficientes para demostrar la regulación hormonal de la división del trabajo por polietismo temporal, en las abejas este problema se ha estudiado con detenimiento. El polietismo temporal de las abejas está regulado por los niveles de dos hormonas, la hormona juvenil (HJ) y la vitelogenina (Vg). La HJ es producida en un pequeño órgano glandular llamado corpora allata ubicado detrás del cerebro. Su función es orquestar la muda y la metamorfosis manteniendo el estado larvario e inhibiendo la metamorfosis que los convertiría en adultos. La Vg, por su parte, es una proteína precursora del vitelo en desarrollo de los oocitos en los insectos. En los insectos sociales, estas hormonas han trascendido su papel en la maduración y reproducción y ahora juegan un papel fundamental en la organización social y división del trabajo regulándose mutuamente: altos niveles de Vg suprimen la HJ, y altos niveles de HJ suprimen la Vg21,22. Cuando las obreras recién eclosionan de la pupación como abejas adultas, comienzan a sintetizar Vg elevando sus niveles en la sangre, lo cual suprime la HJ. Niveles bajos de HJ y altos de Vg mantienen a las obreras en la fase de nodrizas realizando tareas internas. Después de las primeras tres semanas, la actividad del corpora allata aumenta y los niveles de HJ también aumentan, disminuyendo la Vg y comienza la vida forrajera de las abejas. Aunque no se sabe cómo interactúan estas hormonas ni exactamente qué cambios producen en el sistema nervioso, las variaciones hormonales tienen efectos dramáticos en la estructura y función del cerebro, ya que regiones específicas de él incrementan su volumen23. Estos cambios modifican el comportamiento y la fisiología sensorial provocando que las abejas comiencen a forrajear.
Es posible que en aquellas especies de hormigas en las que existe división del trabajo por polietismo temporal, también se presenten cambios fisiológicos y estructurales similares.
]]> Polimorfismo: cada forma una tarea
Entre los insectos sociales, el polimorfismo se refiere a la presencia de varias castas del mismo sexo morfológica y funcionalmente diferentes6, y en todos los casos conocidos el crecimiento alométrico es el responsable del desarrollo de castas físicamente distintas. El crecimiento alométrico ocurre cuando dos partes diferentes del cuerpo crecen a tasas diferentes, de manera que cuando el crecimiento de un grupo de individuos termina en diferentes tiempos, el resultado es que los adultos tendrán no sólo diferentes tamaños, sino también diferentes formas; esto es, diferentes proporciones corporales. De esta manera, las obreras más grandes, típicamente presentan cabezas que son más grandes en relación con el resto del cuerpo, comparadas con obreras minoro media118. En el polimorfismo, generalmente las obreras de un tamaño se especializan en aquella tarea para la cual su tamaño es el más adecuado; e.g., las obreras pequeñas cuidan a la cría y forrajean, las obreras grandes defienden el nido, etc. Ya que la alometría de las castas físicas se encuentra relacionada con la distribución de patrones de comportamiento entre las castas, es muy probable que en el sistema nervioso ocurran cambios alométricos similares.

La forma más representativa de polimorfismo es la presencia, en algunas especies, de enormes obreras llamadas soldados que se dedican principalmente a la defensa. Aun cuando exista el potencial genético para producir estas obreras gigantes, eso no ocurre de manera indiscriminada, ya que existe un patrón del desarrollo que determina que las obreras grandes sólo sean producidas en etapas particulares del desarrollo colonial19.
Aunque el polimorfismo se presenta en hormigas y en termitas, en las abejas y avispas sólo existe un tamaño de obreras, por lo que el tamaño no es definitivo en la determinación de la tarea que realiza un individuo y el polimorfismo por sí solo no explica la división del trabajo. Así, aun cuando en una colonia existan individuos de diferente tamaño, la casta está definida en términos conductuales, por lo que la morfología es una especialización relacionada con la conducta.
La especialización morfológica generalmente produce individuos que difícilmente pueden realizar tareas diferentes a las que están especializados, por lo que si la demanda de obreras de este tipo se reduce, éstas no pueden realizar otras tareas tan eficientemente o no las realizan en absoluto. Debido a que la morfología de los insectos holometábolos está determinada por el tiempo de desarrollo larval, las sociedades polimórficas son poco capaces de responder rápidamente a los cambios ambientales que requieren de ajustes en el nivel social. Por esto, el polietismo físico es poco común entre las sociedades de insectos, encontrándose sólo en cerca del 20% de los géneros de hormigas. Las restricciones para la evolución del polimorfismo son económicas y embriológicas: los especialistas son costosos de producir y la modificación del desarrollo es limitada24, por lo que es más común que las sociedades de hormigas basen su sistema de división del trabajo en el polietismo temporal. Incluso las sociedades polimórficas pueden presentar polietismo temporal, de manera que el polimorfismo y el polietismo temporal producen una distribución poblacional con individuos de diferentes tamaños y edades característica para cada especie: la demografía adaptativa25.
Demografía adaptativa y sociogénesis
El comportamiento de la sociedad está definido por su demografía. Las proporciones de obreras son determinadas principalmente a través de la demografía adaptativa, que consiste en la programación de la producción de individuos de diferentes castas, que resulta en distribuciones de frecuencia de edad y tamaño en la casta obrera que se mantienen constantes18.
Cada especie tiene un programa de diferenciación morfológica y conductual particular y para construir la sociedad, un proceso llamado sociogénesis que consiste en los pasos mediante los cuales los individuos sufren cambios en casta, comportamiento y fisiología. Lo podemos comparar con el proceso análogo de morfogénesis de cualquier organismo multicelular. Las señales ambientales inician una secuencia de restricciones fisiológicas progresivas en los individuos que produce una demografía de la población obrera que incrementa la sobrevivencia y reproducción de la colonia18.
]]>Pocos genes para una gran sociedad
En muchas especies de insectos sociales se ha encontrado que existen factores genéticos que influencian la tendencia de los individuos a realizar una tarea26. Es común que las reinas de los insectos sociales se apareen con varios machos, fenómeno conocido como poliandría. Además de reducir el conflicto reproductivo con las obreras, es posible que la poliandría esté vinculada con la necesidad de heterogeneidad genética dentro de la colonia para producir las diferencias interindividuales necesarias que conduzcan a la compleja división del trabajo11. Las abejas, por ejemplo, se aparean con distintos machos, por lo que la descendencia es genéticamente heterogénea. Las abejas descendientes de diferentes padres difieren en su tendencia a realizar ciertas tareas. Además, forrajeras y nodrizas de abejas tienen diferencias en la expresión de hasta un 40% de genes en el cerebro27.
En algunas especies de hormigas se han reportado casos en los que las castas difieren en sus genotipos26,28-30. En la hormiga cortahojas Acromyrmex echinatior, por ejemplo, se ha encontrado que existe un componente genético del polimorfismo, característica que siempre se supuso como un fenómeno controlado por el ambiente. Los individuos de diferentes líneas paternas tienen diferentes predisposiciones para desarrollarse como obreras minor o mayor. De esta manera la poliandría tiene un papel fundamental en la división del trabajo y el desarrollo del polimorfismo31.
Se han identificado diversos genes involucrados con la división del trabajo. El comportamiento de forrajeo, por ejemplo, en diversas especies de insectos está asociado con la expresión de un gen llamado for (de forrajeo), el cual codifica para una proteína cinasa dependiente de GMPc que se expresa en las neuronas del cerebro32. En las abejas, este gen está implicado en la especialización de la casta de forrajeras pues éstas presentan una expresión más alta de este gen que las nodrizas33. En las hormigas ocurre lo opuesto: mientras que las obreras jóvenes presentan altos niveles de expresión de este gen, las obreras forrajeras presentan bajos niveles34. Los efectos de este gen pueden ser de largo plazo en la organización del sistema nervioso, y de corto plazo sobre la función de las neuronas del sistema sensorial y del SNC.
Otro gen involucrado es el gen period, un componente del reloj biológico. Conforme las obreras envejecen y cambian de tareas, comienzan a realizar sus tareas preferentemente de día y se manifiesta un ritmo circadiano de actividad-reposo, así como oscilaciones del gen period. Este ritmo es fundamental durante las actividades diarias de forrajeo35.
Muchos otros genes podrían estar relacionados con el comportamiento social. Lo interesante es que no necesariamente son nuevos genes. Muchos genes que presentan los animales solitarios, como el period, el for, el malvolio, etc., han sido aprovechados por la evolución social y ahora son usados para el comportamiento social. El punto crucial de transición es la sensibilidad de esos genes para ser regulados por el ambiente social y modificar el sistema nervioso y el comportamiento de los individuos de acuerdo con el contexto social27.
Pero, ¿quién hace qué y cuándo?
La decisión de una obrera individual para realizar una tarea es el fundamento de la división del trabajo y apenas estamos respondiendo a preguntas como: ¿cómo funciona la distribución de las tareas y qué estímulos, condiciones, factores, etc., determinan, en un momento dado, qué tarea realiza una obrera y cómo ajusta una colonia sus esfuerzos a las demandas del ambiente?, ¿cuáles son los mecanismos por los cuales los miembros de la colonia se diferencian en castas y se reparten el trabajo?
]]> Actualmente sabemos que existen factores internos basados en atributos de los individuos y factores externos basados en estímulos ambientales transitorios, que interactúan y determinan la ejecución de una tarea individual por una obrera36. La probabilidad de que una obrera realice una tarea depende de: 1) la magnitud de la tarea (factores externos), es decir, qué tan importante o urgente es la tarea y cuántas obreras son necesarias para llevarla a cabo y 2) la sensibilidad individual a los estímulos relacionados con dicha tarea (factores internos). Aunque se ha sugerido en repetidas ocasiones que las obreras podrían presentar diferencias específicas de la casta en la percepción sensorial y/o en las respuestas conductuales a los estímulos relacionados con una tarea20, esto no ha sido debidamente probado y sólo existen algunos reportes en los que se han realizado comparaciones entre castas (por ejemplo López-Riquelme et al., 200637).
El justo medio: ni tan especializados, ni tan flexibles
Aunque hemos hecho hincapié en que la división del trabajo implica una especialización en mayor o menor grado, generalmente esta división es plástica y las obreras pueden cambiar de tareas según se requiera. Esto se logra gracias a la flexibilidad conductual de los individuos, e incluso las castas físicas más especializadas pueden ser inducidas a realizar tareas fuera de su repertorio normal. Por ejemplo, cuando la proporción de obreras minor:mayor de la hormiga Pheidole baja del normal 3:1 ó 20:1y llega a1:1, las mayor comienzan a realizar tareas que normalmente sólo realizan las minor19. La plasticidad en el comportamiento de las castas es un factor importante en la organización de una colonia, pero requiere de complejos cambios fisiológicos en los individuos, por lo que también existen límites: las mayor de Pheidole pueden cambiar su repertorio para realizar las tareas de las minor, pero las minor no pueden realizar las tareas de las mayor20.
Como los miembros de la colonia no pueden obtener información acerca del estado de la colonia completa, la información puede ser adquirida a través de las interacciones con otras obreras o a partir del estado del nido. La decisión de las tareas a realizar podría estar basada en la integración adquirida a partir de las interacciones sociales acopladas con las predisposiciones conductuales asociadas con la casta o el estado fisiológico, lo cual puede inducir cambios en la actividad endocrina y genética de las obreras, induciendo cambios en el comportamiento20. Esta regulación sistémica es muy compleja y se parece mucho a la que ocurre en los tejidos y órganos de los organismos multicelulares. Por esto, es más apropiado tratar a la colonia como una unidad funcional, un superorganismo en vez de como a un grupo de individuos.
El superorganismo
Las hormigas tienen repertorios conductuales relativamente simples, sin embargo, vista desde lejos y en su ambiente, la colonia de hormigas puede considerarse como un superorganismo; esto es, una entidad grande y difusa cuyo aparato de ingestión, compuesto por las obreras forrajeras, se mueve hacia adelante y hacia atrás sobre el territorio circundante en un ritmo circadiano5,19. Es la colonia, no los individuos, la que compite con otras entidades semejantes por los recursos y territorio.
La existencia de similitudes entre los organismos y las sociedades ha sido reconocida desde hace tiempo. No obstante, para la mayoría de los biólogos evolucionistas el concepto de superorganismo invoca el de selección de grupo el cual ha sido rechazado en múltiples ocasiones38. Sin embargo, para los sistemistas existen suficientes características que permiten considerar a la colonia como un organismo5:
1) Se comporta como una unidad a partir de la función cooperativa de muchos miembros.
]]> 2) Muestra características sistémicas especie-específicas.3) La colonia pasa por un ciclo de crecimiento y reproducción que es adaptativo.
4) La colonia está diferenciada en un "germoplasma" (machos y reinas) y un "soma" (obreras).
5) La reproducción de un superorganismo produce descendencia superorganísmica.
6) Los superorganismos generalmente son sésiles (pero no sus miembros).
7) Los superorganismos mantienen homeostasis intraorganísmica.
8) Los superorganismos se encuentran bien armados contra los enemigos o son crípticos
Un organismo y un superorganismo son sistemas complejos, organizados, coordinados e individualizados de actividades, las cuales son primariamente dirigidas a la obtención y asimilación de sustancias del medio, tanto para producir otros sistemas similares conocidos como descendencia, como para proteger al sistema mismo y a dicha descendencia de las perturbaciones ambientales19. Ambos sistemas se encuentran arreglados en al menos dos tipos diferenciados de subsistemas: el soma y la línea germinal39.
Ejemplos de comportamiento de superorganismo son: la distribución de información basada en la emisión de feromonas y contactos físicos; comparables a la organización del sistema nervioso y emisión de hormonas en los animales, la distribución del alimento boca a boca (trofalaxia), mecanismo comparable a la transferencia de nutrientes por el flujo sanguíneo; la demografía adaptativa, que se refiere a la formación de nuevos individuos en respuesta a las pérdidas ocasionadas por diversos factores, mecanismo comparable a la regeneración tisular en respuesta a un daño en los organismos multicelulares. La homeostasis social también involucra el control ambiental, como por ejemplo, el mantenimiento de una temperatura más o menos constante dentro del nido.
No obstante lo anterior, la diferencia entre organismos multicelulares y organismos sociales es clara. Todas las células de un organismo son genéticamente idénticas, y entre los individuos de una colonia de hormigas existe una considerable heterogeneidad genética, la cual podría generar conflictos por la diversidad de intereses reproductivos. Pero, ¿a qué nivel actúa la selección?
]]>Niveles de selección
Resulta evidente que los seres vivos pueden encontrarse jerárquicamente organizados en lo que se ha denominado niveles de organización. Sin embargo, a pesar de lo común de estos niveles de organización y de las unidades autónomas resultantes, la discusión acerca de en qué nivel actúa la selección natural ha sido polémica17.
La teoría de los niveles de selección ha influenciado de manera importante nuestro entendimiento de la selección natural y los niveles en donde actúa. Existen dos soluciones al problema, que las unidades de selección sean los genes o individuos, o que sean los grupos de individuos40, y hay varios aspectos en relación a esto. Primero, el reconocimiento de la jerarquía biológica. Segundo, que la unidad de selección legítima pueden ser tanto los genes como los grupos de individuos. Tercero, que exista una correspondencia entre la operación de la selección en diferentes niveles. Finalmente, que la selección pueda actuar simultáneamente en diferentes niveles. Cuando la selección actúa en direcciones opuestas en niveles adyacentes, el resultado será un compromiso entre las fuerzas de selección en cada nivel17.
La polémica surgió debido a que se reconoció que los organismos no se replican en el proceso de reproducción, ya que todos los organismos mueren y sólo sus genes pasan a las siguientes generaciones. Eso condujo a Dawkins (1993)38 a proponer que la unidad fundamental de selección es el gen, una unidad de herencia llamada replicador. Esta perspectiva reduccionista dejó de lado que en los individuos los genes se encuentran agrupados en sociedades y no actúan aislados. Los genes cooperan unos con otros en forma coordinada, en las células solitarias, en los organismos multicelulares y en las sociedades, y la selección natural ha favorecido esta coordinación a tal grado, que esas sociedades de genes funcionan como entidades individuales. Es a través del comportamiento de la sociedad que los genes, incapaces de una vida solitaria, se separan al paso de las generaciones. Esto significa que los genes aislados no son visibles para la selección natural, pues ésta no puede seleccionar directamente entre genes individuales o paquetes, sino entre fenotipos. Así, puede hacerse una diferencia entre la unidad de replicación y el vehículo, el cual puede ser un organismo o un grupo de organismos. La unidad de replicación es una pieza de información genética, mientras que el vehículo es la entidad sobre la cual actúa directamente la selección y contiene la información genética. De esta manera, los individuos, así como otras unidades de nivel superior, pueden ser vehículos y, por tanto, sujetos de selección40.
La individualidad se mantiene mediante la comunicación química
Las hormigas viven en sociedades complejas coordinadas por un sofisticado sistema de comunicación química a través de feromonas. Todos los niveles de la socialidad de las hormigas tienen una base química: la regulación, integración y cohesividad dependen de ella. Las respuestas a las feromonas dependen de la concentración y del contexto, por lo que una sola feromona puede tener diferentes funciones. Hay dos grupos importantes de feromonas en las hormigas. El primero es el olor de la colonia, el cual reside en la cutícula y participa en la discriminación entre compañeros y extraños. El segundo grupo incluye feromonas producidas en glándulas y que liberan respuestas especie-específicas24.

Una de las principales ventajas de la comunicación es que puede permitir que una sociedad se mantenga más o menos cerrada reconociendo lo que no le pertenece, de manera que sus beneficios permanezcan confinados a sus miembros. Por lo tanto, muchas señales están involucradas en el reconocimiento de la membresía y el parentesco19.
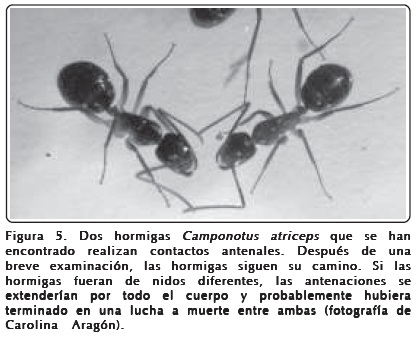
]]> Todos los miembros de una colonia tienen "olores pasaporte". Este olor de la colonia se extiende sobre la superficie del cuerpo de cada hormiga y el reconocimiento de los compañeros de nido es aprendido en un corto tiempo después de la eclosión. Se ha sugerido que este aprendizaje es una forma de impronta que es detectada mediante quimiorrecepción por contactos antenales. Estas señales de reconocimiento están constituidas principalmente por una mezcla particular de hidrocarburos cuticulares que podría formar una experiencia sensorial analizada por la totalidad que forman sus componentes. Los hidrocarburos que componen este olor de reconocimiento provienen de diferentes fuentes, principalmente un órgano glandular ubicado en la cabeza, llamado glándula postfaríngea. Cada hormiga produce una mezcla de feromonas cuya composición depende de la constitución genética y fisiológica e incluso de la dieta. Con los cotidianos contactos físicos entre hormigas, con el aseo mutuo, el intercambio de alimento, etc., los olores son pasados de uno a otro individuo, mezclándose y formando el olor de la colonia.Cuando las hormigas van en fila en busca de alimento o se encuentran en las afueras de su nido, las hormigas frecuentemente se detienen por un momento y se tocan unas a otras con sus antenas. Qué es lo que hacen, es difícil de saber, pero es claro que se tocan por muy breves momentos (antenear lo llaman los especialistas) y luego continúan su marcha. Como los receptores olfativos de las hormigas se encuentran en las antenas, es fácil imaginar que durante esos breves contactos se identifican por medio de los compuestos que se encuentran en la cutícula. Sabemos que los receptores en las antenas son sensibles a esas moléculas, por lo que imaginamos que en esa forma las hormigas de un nido pueden identificar a otras del mismo nido, así como a las de otros nidos.
Olfato: el principal sentido de las hormigas
Las hormigas viven en un mundo de olores, donde la visión tiene un papel muy restringido y nos es imposible imaginar ese mundo lleno de mensajes que viajan en el aire por medio de moléculas y que son detectadas por las antenas y procesadas en sus pequeños cerebros. La percepción de señales químicas juega un papel esencial en la sobrevivencia y organización de las sociedades de hormigas. Por lo tanto, su sistema olfativo debe tener elaborados mecanismos capaces de detectar diversas señales y liberar el comportamiento adecuado. Debido a esto, las adaptaciones neurofisiológicas para la vida social están basadas principalmente en especializaciones de las vías olfativas y su entendimiento es fundamental para la comprensión del comportamiento social. La descripción estructural de la vía olfativa en el sistema nervioso central en las hormigas ha revelado una organización altamente conservada y especializada para la vida social41,42.
La detección de moléculas en el aire se hace por medio de receptores que se encuentran en órganos especializados sobre las antenas, llamados sensilas, y sabemos que existen muchos tipos de receptores. Sin embargo, como el número de olores depende del número de moléculas emitidas y éstas parecerían ser una enorme cantidad, nos preguntamos si también es necesario tener una gran cantidad de receptores, quizá tantos como moléculas, pero eso no parece ser así. Por ejemplo, sabemos que aproximadamente el 3% de nuestros genes se usan para codificar receptores a diferentes moléculas odorantes y que en cada célula receptora se expresa sólo un tipo de receptor. Así, los ratones tienen aproximadamente 900 genes que codifican para 1,200 receptores a odorantes, mientras los humanos tenemos entre 322 y 900 genes de este tipo. Entonces, ¿cómo es posible que con tan pocos receptores seamos capaces de identificar aproximadamente 10,000 olores diferentes?
La respuesta no la sabe nadie, todavía, pero hay quienes piensan que cada receptor es como la letra de un alfabeto, de manera que una molécula odorífica puede ser captada por varios receptores cuyas letras formarían una palabra. Así, si el idioma español tiene 31 letras y podemos escribir tantos libros, es fácil imaginar que con unos 500 tipos de receptores el número de palabras que podrían formarse es enorme. Para tener una idea de las posibilidades de un sistema como éste, el número de bases en el código genético es de sólo 4 y, sin embargo, con ellas se puede crear un número casi infinito de secuencias genéticas. Otros en cambio piensan que los receptores olfativos actúan como espectroscopios biológicos, donde lo que activa al receptor son las vibraciones de la molécula de odorante, y cómo aún la más pequeña diferencia en estructura molecular produce un espectrograma vibracional diferente, una serie de receptores pueden cubrir todo el espectro olfativo y, por tanto, todos los olores posibles.
El cerebro de las hormigas: un órgano pequeño pero complejo
]]> Aunque el cerebro de las hormigas es pequeño, tiene una alta sofisticación cognitiva. Es en este minicerebro de menos de 0.06 milímetros cúbicos donde el comportamiento individual complejo es generado y controlado. Pero el volumen absoluto del cerebro no dice gran cosa, sino el volumen relativo. Consideremos aquellas especies polimórficas en las que existen enormes soldados con cabezas gigantes y grandes mandíbulas. En estos individuos, la relación volumen cerebral: tamaño-corporal es menor que en las obreras pequeñas, ya que gran parte del espacio cefálico es ocupado por los músculos mandibulares. Por ejemplo, aunque el tamaño de la cabeza de obreras mayor y minor de Camponotus floridanus presenta grandes diferencias, los cerebros de las mayor sólo son cerca de un 8% más grandes que los de las minor. Entre las castas mayor y minor de Atta ocurre algo parecido: en las obreras minor el cerebro ocupa el 60% del volumen total de la cabeza, mientras que en las obreras mayor apenas el 5%43.Si comparamos el cerebro de las hormigas con el de sus parientes sociales, como las abejas y las avispas, podemos observar que se parecen mucho. Sin embargo, en las hormigas, los ojos y los lóbulos ópticos (regiones que procesan la información visual), se encuentran muy reducidos, lo cual refleja la limitada importancia que la visión juega en la vida de las hormigas. Esto tiene una explicación. Las abejas y avispas son insectos voladores que localizan fuentes de alimento y se orientan por medio de la visión. Por el contrario, las hormigas viven en nidos bajo el suelo o rocas y no son voladoras, excepto por los sexuados durante el apareamiento (quienes sí presentan enormes ojos y lóbulos ópticos asociados). Aunque las hormigas pueden orientarse por medio de la visión, en general dependen del olfato para ello, ya que siguen rastros olorosos que otras hormigas han dejado sobre el suelo. Además, todos los insectos sociales basan su organización social en la comunicación química.

Contrario a lo que ocurre con el sistema visual, el sistema olfativo de las hormigas está muy desarrollado. Aunque las hormigas no tienen las antenas más grandes ni complejas entre los insectos, pueden presentar varios miles de sensilas, minúsculos órganos olfativos compuestos cada uno por varias neuronas receptoras a los olores. Además, las regiones cerebrales que procesan las señales provenientes de las antenas, los lóbulos antenales (LA), se encuentran muy desarrolladas, ya que son complejas y muy grandes: pueden representar hasta el 12% del volumen total del cerebro. Otra región cerebral muy desarrollada en las hormigas son los cuerpos fungiformes (CF), una región que procesa e integra información visual, olfativa y mecanosensorial y que controla el comportamiento complejo. Es tal la importancia de los CF, que puede constituir hasta el 55% del volumen total del cerebro y las diferencias en su tamaño entre las castas están relacionadas con la función de cada una. Sin embargo, el volumen relativo de los LA y de los CF es constante en muchas especies de hormigas, por lo que se ha sugerido que existe un cerebro hormiga "estándar", con una composición evolutivamente muy estable: con los CF constituyendo el doble del volumen de los LA43. Esto se debe principalmente a que los cerebros de las hormigas están especializados en el procesamiento de información olfativa.
Consideremos la organización estructural de las principales regiones olfativas del cerebro de las hormigas. Las neuronas que forman las sensilas de las antenas tienen sus dendritas inmersas en un líquido que está en contacto con el ambiente a través de poros en la cutícula. Las moléculas del aire difunden a través de estos poros cruzando el fluido para después entrar en contacto con receptores específicos de la membrana de la neurona, lo cual resulta en la apertura de canales catiónicos a través de segundos mensajeros. El efecto es la alteración de las propiedades eléctricas de la neurona receptora, generando un potencial receptor y potenciales de acción hacia los axones. Esta información olfativa es llevada hacia el cerebro a través de cientos de axones que corren por toda la antena hasta la cabeza en forma de un nervio. Una vez que llegan al cerebro olfativo llamado lóbulo antenal (LA), los axones que componen el nervio se separan en diferentes tractos que inervan grupos de estructuras esferoidales llamadas glomérulos41,42. Otras neuronas ubicadas dentro del LA conectan hasta los 400 glomérulos que presentan las hormigas ya sea entre ellos o con regiones exquisitamente compartamentalizadas en el cerebro superior, áreas comparables a la corteza cerebral de los vertebrados y que han sido llamadas cuerpos fungiformes (CF), debido a su forma de hongo. Los CF están compuestos de neuronas llamadas células Kenyon, cuyas dendritas forman el neuropilo de los CF, una región aferente (que recibe información) en forma de copa llamada el cáliz. La parte eferente (que manda información) está formada por los axones que salen de cada árbol dendritico de dichas células dando origen primero a un pedúnculo y, después de ramificarse, a dos lóbulos (los lóbulos α y β). El cáliz está dividido en tres compartimientos discretos: 1) el labio, que recibe información olfativa del LA; 2) el collar, que recibe información visual y 3) el anillo basal, que recibe información tanto olfativa como visual44. Estos compartimientos están subdivididos, a su vez, en varias capas de acuerdo con las aferencias y la organización del neuropilo sensorial primario (ya sean los LA o los lóbulos ópticos). Esta separación de la información aferente representa diferentes propiedades visuales y olfativas formando un mapa abstracto de la organización de los neuropilos sensoriales en el cáliz41-44. De esta manera, los CF reciben información multisensorial en muchas capas segregadas de manera que puede ser combinada para formar una imagen compleja del ambiente y del resto del animal.
Como hemos visto, la base de las sociedades de las hormigas es la comunicación química y el comportamiento de las hormigas muestra esto en la forma más clara, pero también lo muestra la organización de su cerebro, el cual básicamente es un cerebro social que principalmente procesa información olfativa. Como sea que funcionen los receptores, ya sea formando espectrogramas o palabras, sabemos que las hormigas son capaces de detectar una plétora de estímulos de naturaleza química provenientes del ambiente o de la sociedad, de manera que fácilmente pueden distinguir una hormiga del mismo nido o de otro nido diferente.
Nosotros hemos podido registrar la actividad eléctrica en las diferentes regiones de la vía olfativa, desde la antena hasta el cerebro, en respuesta a pequeños soplos que contienen moléculas interesantes para ellas. El registro que se realiza en la antena, se llama "electroantenograma" y a nosotros nos ha permitido estudiar no sólo qué moléculas detectan las hormigas, sino también, por medio de otro registro llamado electroencefalograma, qué información mandan las antenas al cerebro y qué hacen las neuronas del cerebro cuando llega a ellas, es decir, cómo se codifican las propiedades de los estímulos en las diferentes partes de la vía olfativa. Con estos estudios esperamos poder asociar eventos eléctricos particulares en las antenas y el cerebro con algunos comportamientos aparentemente azarosos de las hormigas, así como aquellos a los que nosotros les podemos dar una intención clara.
Conclusiones: cerebros sociales, genomas sociales
]]> El sistema nervioso y sensorial de las hormigas parece estar bien adaptado a la vida social, ya que gran parte de su maquinaria neuronal ha sido liberada de los compromisos de la vida en solitario; por ejemplo, la búsqueda de pareja, la construcción del nido, la evasión de depredadores, la oviposición, el cuidado de la cría, etc. Con la liberación de esas tareas debido a la división del trabajo y a la cooperación, el sistema nervioso de las hormigas fue sometido a las presiones de la vida en sociedad. Ya que el éxito y sobrevivencia de cada hormiga depende de su capacidad de trabajar coordinadamente para lograr la reproducción de la reina es fundamental que su susceptibilidad a responder a estímulos sociales esté bien desarrollada.Es probable que la evolución del sistema nervioso en insectos sociales haya conducido hacia la complejidad y especialización de los centros cerebrales que participan en la comunicación: 1) sistemas sensorioperceptuales que involucran a los órganos sensoriales periféricos y/o a las áreas involucradas en el procesamiento de las señales comunicativas (LA) y 2) áreas de asociación y procesamiento de información multimodal y de generación, coordinación y control de patrones motores y conductuales complejos (CF). Esto se debe principalmente a la dependencia de la vida social de las hormigas en la comunicación química, por lo que es posible que las estructuras neuronales implicadas en estos procesos sociales se hayan visto sujetas a fuertes presiones de selección dentro de la sociedad.
La idea fundamental es que tanto la estructura y función cerebral, como la expresión génica, son sensibles al contexto social y se han propuesto vías de comunicación entre el sistema nervioso y el genoma que contribuirían al control del comportamiento social. Así, la información adquirida por el sistema nervioso bajo ciertas condiciones sociales puede inducir cambios en la función genómica que, a su vez, modificarían la estructura y función del sistema nervioso. Por lo tanto, uno de los aspectos más importantes y trascendentes del estudio del cerebro hormiga, un cerebro social, puede mostrarnos los mecanismos neurofisiológicos del comportamiento social y cómo la evolución social afecta la evolución cerebral.
A final de cuentas, los insectos sociales pueden enseñarnos mucho acerca del funcionamiento y de la evolución de los sistemas sociales, y, por qué no, como lo desearon muchos mirmecólogos del pasado, también podrían enseñarnos mucho sobre nosotros mismos.
Agradecimientos
Facultad de Medicina, UNAM y DGAPA, UNAM.
Referencias
1. Erber, J. & Page, R.E. The evolutionary dynamics of social organization in insect societies: from behavior to genes and back. Santa Fe Institute Bulletin (Verano 2001). [ Links ]
2. Hölldobler, B. & Wilson, E.O. Journey to the Ants: A story of scientific exploration (Harvard University Press, 1995). [ Links ]
3. Tshinkel, W.R. The nest architecture of the Florida harvester ant, Pogonomyrmex badius. Journal of Insect Science4, 21 (2004). [ Links ]
4. Wilson, E.O. The social biology of ants. Annu. Rev. Entomol. 8, 345-368 (1963). [ Links ]
5. Hölldobler, B. & Wilson, E.O. The superorganism (Norton, 2009). [ Links ]
6. Wilson, E.O. The insect societies (Harvard University Press, 1971). [ Links ]
7. Duffy, J.E. The ecology and evolution of eusociality in sponge-dwellingshrimp. In Genes, Behavior, and Evolution in Social Insects (ed. Kikuchi, T.) 217-254 (University of Hokkaido Press, Sapporo, Japan, 2003). [ Links ]
8. Sherman, P.W. The biology of naked-mole rat (Princeton, 1991). [ Links ]
9. Haldane, J.B.S. "Population Genetics". New Biology 18, 34-51 (1955). [ Links ]
10. Hamilton, W.D. The genetical evolution of social behavior, I y II. J. Theor. Biol. 7, 1-52 (1964). [ Links ]
11. Croizer, R.H. & Pamilo, P. Evolution of social insect colonies. Sex allocation and kin selection (Oxford University Press, 1996). [ Links ]
12. Trivers, R.L. & Willard, D.E. Natural selection of paternal ability to vary the sex-ratio of offspring. Science 179, 90-92 (1973). [ Links ]
13. Trivers, R.L. & Hare, H. Haplodiploidy and the evolution of the social insects. Science 191, 249-263 (1976). [ Links ]
14. Jaisson, P. La hormiga y el sociobiólogo (FCE, México, 2000). [ Links ]
15. Korb, J. & Heinze, J. Multilevel selection and social evolution of insect societies. Naturwissenschaften 91, 291-304 (2004). [ Links ]
16. Wilson, E.O. Kin selection as the key of altruism: its rise and fall. Social Research 72(1), 159-166 (2005). [ Links ]
17. Bourke, A.F.G. & Franks, N.R. Social evolution in ants (Princeton University Press, 1995). [ Links ]
18. Wilson, E.O. The sociogenesis of insect colonies. Science 228 (4707), 1489-1495 (1985). [ Links ]
19. Hölldobler, B. & Wilson, E.O. The Ants (Harvard University Press, 1990). [ Links ]
20. Robinson, G.E. Regulation of division of labor in insect societies. Ann. Rev. Entomol. 37, 637-665 (1992). [ Links ]
21. Hartfelder, K. Insect juvenile hormone: from status quo to high society. Brazilian Journal of Medical and Biological Research 33, 157-177 (2000). [ Links ]
22. Denison, R. & Raymond-Delpech, V. Insights into the molecular basis of social behavior from studies on the honey bee, Apis mellifera. Invert. Neurosci. 8, 1-9 (2008). [ Links ]
23. Fahrbach, S.E. & Robinson, G.E. Behavioral development in the honey bee: toward the study of learning under natural conditions. Learning & Memory 2, 199-224 (1995). [ Links ]
24. Sudd, J.H. & Franks, N.R. The behavioural ecology of ants (Chapman and Hall, 1987). [ Links ]
25. Tofts, C. & Franks, N.R. Doing the right thing: ants, honeybees and naked mole-rats. TREE 7(10), 346-349 (1992). [ Links ]
26. Parker, J. D. A major evolutionary transition to more than two sexes? Trends Ecol. Evol. 2, 83-86 (2004). [ Links ]
27. Robinson, G.E. Sociogenomics: social life in molecular terms. Nature Reviews Genetics 6, 257-271 (2005). [ Links ]
28. Snyder, L.E. Non-random behavioral interactions among genetic subgroups in a polygynous ants. Anim. Behav. 46, 431-439 (1993). [ Links ]
29. Fraser, V.S., Kaufmann, B., Oldroyd, B.P. & Croizer, R.H. Genetic influence on caste in the ant Camponotus consobrinus. Behav. Ecol. Sociobio. 47, 188-194 (2000). [ Links ]
30. Stuart, R.J. & Page, R.E. Genetic component to division of labor among workers of a Leptothoracine ant. Naturwissenschaften 78, 375-377 (1991). [ Links ]
31. Hughes, W.O.H., Summer, S., Van Borm, S. & Boomsma, J.J. Worker caste polymorphism has a genetic basis in Acromyrmex leaf-cutting ants. PNAS 100(16), 9394-9397 (2003). [ Links ]
32. Ben-Shahar, Y. The foraging gene, behavioral plasticity, and the honey bee division of labor. J. Comp. Physiol. A 19, 987-994 (2005). [ Links ]
33. Ben-Shahar, Y., Leung, H.T., Pak, W.L., Sokolowski, M.B. & Robinson, G.E. cGMP-dependent changes in phototaxis: a possible role for the foraging gene in honey bee division of labor. J. Exp. Biol. 206, 2507-2515 (2003). [ Links ]
34. Ingram, K., Oefner, P. & Gordon, D.H. Task-specific expression of the foraging gene in harvester ants. Molecular Ecology 14, 813-818 (2005). [ Links ]
35. Bloch, G., Sullivan, J.P. & Robinson, G.E. Juvenile hormone and circadian locomotor activity in the honey bee Apis mellifera. J. Insect. Physiol. 48, 1123-1131 (2002). [ Links ]
36. Beshers, S.N. & Fewell, J.H. Models of division of labor in social insects. Annu. Rev. Entomol. 46, 413-440 (2001). [ Links ]
37. López-Riquelme, G.O., Malo, E.A., Cruz-López, L. & Fanjul-Moles, M.L. Antennal olfactory sensitivity in response to task-related odours of three castes of the ant Atta mexicana (Hymenoptera: Formicidae). Physiological Entomology 31, 353-360 (2006). [ Links ]
38. Dawkins, R. El gen egoísta (Salvat Editores, España, 1993). [ Links ]
39. Wilson, D.S. & Sober, E. Reviving the superorganism. J. Theor. Biol. 136, 337-356 (1989). [ Links ]
40. Reeve, H.K. & Keller, L. Levels of selection: burying the units of selection debate and unearthing the crucial new issues. In Levels of selection in evolution (ed. Keller, L.) 3-14 (Princeton University Press, 1999). [ Links ]
41. Gronenberg, W. & López-Riquelme, G.O. Multisensory convergence in the mushroom bodies of ants and bees. Acta Biologica Hungarica 55(1-4), 31-37 (2004). [ Links ]
42. López-Riquelme, G.O. Hormigas como sistemas modelo para el comportamiento complejo. Bases neurobiológicas de la comunicación química y la división del trabajo en las hormigas. Tesis de Doctorado (Facultad de Ciencias, UNAM, 2009). 215 págs. [ Links ]
43. Gronenberg, W. & Hölldobler, B. Morphologic representation of visual and antennal information in the ant brain. J. Comp. Neurol. 412, 229-240 (1999). [ Links ]
44. Gronenberg, W. Subdivisions of Hymenopteran mushroom body calyces by their afferent supply. J. Comp. Neurol. 430, 474-489 (2001). [ Links ]
Información sobre los autores
Germán Octavio López-Riquelme
]]> Germán Octavio López-Riquelme es biólogo egresado de la Facultad de Ciencias de la UNAM. Realizó el Doctorado en Ciencias Biológicas con especialización en Neuroetología en el laboratorio de Neurofisiología Comparada de la Facultad de Ciencias de la UNAM. Realizó una estancia de investigación en Arizona Research Laboratories. Division of Neurobiology de la Universidad de Arizona en Tucson, Az, USA y posteriormente dos estancias más de investigación en el Laboratorio de Ecología Química de Insectos en El Colegio de la Frontera Sur (ECOSUR), en Tapachula, Chiapas. Es apicultor aficionado con un gran interés en la cría de reinas y el mejoramiento genético apícola. Ha sido pionero en el estudio de las bases neurobiológicas del comportamiento social en hormigas y otros insectos sociales en México. Ha publicado los resultados de su investigación sobre hormigas en revistas indexadas y se ha presentado en congresos nacionales e internacionales. Fue invitado por la Universidad de Illinois para impartir una conferencia sobre la estructura de las conexiones del sistema nervioso en los insectos sociales. Además de sus actividades de investigación, ha sido docente, impartiendo cursos y seminarios a estudiantes de licenciatura y posgrado. En el ámbito de la divulgación ha publicado 17 artículos en diferentes revistas como: Ciencias. ¿Cómo ves? y México desconocido, entre otras. Además, ha participado como asesor científico del Museo de las Ciencias Universum e impartido conferencias de divulgación en la UNAM y en otras instituciones educativas privadas. También ha sido entrevistado para programas de radio y prensa escrita. Actualmente realiza una estancia posdoctoral en el Laboratorio de Socioneurobiología de la Facultad de Medicina de la UNAM en donde hace una investigación sobre la codificación olfativa en el cerebro de las hormigas.Fidel Ramón
Fidel Ramón estudió la licenciatura en la Facultad de Medicina de la UN AM y en 1968 viajó a los Estados Unidos de Norteamérica para hacer un doctorado en Fisiología y Biofísica en la Universidad de Duke, en Carolina del Norte. Al término del doctorado fue contratado por la misma Universidad, durante 15 años y en ese tiempo trabajó con axones gigantes de calamar en el Laboratorio de Biología Marina de Woods Hole, Mass, EUA, sobre biofísica de canales iónicos. En 1983 regresó a México, al Departamento de Fisiología y Biofísica del CINVESTAV del IPN. Donde permaneció hasta 1993 y posteriormente se incorporó a la-Facultad de Medicina de la UNAM. El Dr. Ramón pertenece al SNI, nivel II, y a varias academias científicas, ha publicado numerosos trabajos de investigación en revistas internacionales que son citados frecuentemente. Actualmente investiga además del problema del sueño utilizando como animal modelo el acocil, la neurofisiología del comportamiento en las hormigas.
]]>