
Combining ability effects in summer squash hybrids type Grey Zucchini
César Sánchez–Hernández1*; Clemente Villanueva–Verduzco2; Jaime Sahagún–Castellanos2; Juan Martínez–Solís2; Juan Porfirio Legaria–Solano2; Miguel Ángel Sánchez–Hernández3
1 Instituto Tecnológico Superior de Cosamaloapan. Av. Tecnológico s/n Col. Los Ángeles C. P. 95400 Cosamaloapan, Ver. MÉXICO. Tel: 01(288) 8823100. Correo–e: cesarsh79@hotmail.com (*Autor para correspondencia).
2 Departamento de Fitotecnia. Universidad Autónoma Chapingo, km 38.5 Carretera México–Texcoco. C. P. 56230. Chapingo, Estado de México. Tel. y Fax: 01 (595) 952–1500.
3 Universidad del Papaloapan. Av. Ferrocarril Hidalgo s/n. Ciudad Universitaria Loma Bonita, Oaxaca, México. C. P. 68400 Tel. y Fax: 01(281) 872–2239.
]]> Recibido: 11 de noviembre, 2010.
Resumen
Se evaluaron ocho híbridos comerciales de calabacita (Cucúrbita pepo L.) y sus 56 cruzas posibles con el Método I Modelo II de Griffing (1956) en Chapingo, México durante 2007 y 2008, con el propósito de identificar las mejores combinaciones entre los híbridos comerciales con base en sus efectos de aptitud combinatoria general (ACG) y aptitud combinatoria específica (ACE), para iniciar a corto plazo un programa de mejoramiento y la obtención de variedades sintéticas de calabacita. El diseño experimental utilizado fue bloques completos al azar con tres repeticiones; la unidad experimental consistió en dos surcos de 5 m, separados a 0.8 m y 0.2 m entre plantas (62,500 plantas·ha–1). Los resultados del análisis combinado indicaron que en la población teórica obtenida de la combinación de las cruzas F1 entre híbridos comerciales de calabacita, fueron más importantes los efectos de dominancia para los principales componentes del rendimiento: frutos por planta, rendimiento por planta y por hectárea, y los aditivos para número de días a floración femenina. Es factible mejorar la población por selección hacia precocidad. De las seis mejores cruzas de calabacita para rendimiento de verdura, tres podrían utilizarse sólo como híbridos (ACG < ACE): Hurakán x Lolita (52.16 t·ha–1); Lolita x Tala (50.14 t·ha–1) y Tala x Terminator (47.72 t·ha–1); dos como variedades sintéticas (ACG > ACE): WA9041 x Lolita (51.45 t·ha–1) y Grey Zucchini M+M x Terminator (50.00 t·ha–1) y una como población base para mejoramiento por selección recurrente, ya que tanto los efectos aditivos como los de dominancia son importantes (ACG = ACE) en la cruza: Lolita x Terminator (44.09 t·ha–1).
Palabras clave: Cucurbita pepo L., aptitud combinatoria general, aptitud combinatoria específica, efectos maternos, efectos recíprocos.
Abstract
Eight commercial hybrids of summer squahs type 'Grey Zucchini' (Cucúrbita pepo L.) and their 56 possible crosses of a diallel using the Method I Model II of Griffing (1956) were evaluated in Chapingo Mexico, during 2007 and 2008; to identify the best combinations among the commercial hybrids according to the general combining ability (ACG) and specific combining ability (ACE) effects; and to begin a genetic improvement program to get squash synthetic varieties. The experimental design used was a randomized complete block with three replications. The experimental plot consisted in two rows of 5 meters, with row separation of 0.80 m and 0.2 m between plants (62,500 plants–ha–1). The combined analysis showed that dominance effects for the main components of the yield: fruit per plant, yield per plant and hectare were more important, and number of days to female flowering additive effects were important and make it feasible to improve precocity. From the best six crosses for squash fruit yield, three of them could only be used as hybrids (ACG < ACE): Hurakan x Lolita (52.16 t·ha–1); Lolita x Tala (50.14 t·ha–1); Tala x Terminator (47.72 t·ha–1); two as synthetic varieties (ACG > ACE): WA9041 x Lolita (51.45 t·ha–1); Grey Zucchini M+M x Terminator (50.00 t·ha–1) and one as a base population to be improved by recurrent selection, since the additive and of dominance effects are important (ACG = ACE): Lolita x Terminator (44.09 t·ha–1).
Key words : Cucurbita pepo L., general combining ability, specific combining ability, maternal effects, reciprocal effects.
]]>INTRODUCCIÓN
La caracterización de los progenitores por su aptitud combinatoria general (ACG) y aptitud combinatoria específica (ACE), permite establecer las estrategias y técnicas adecuadas para estimar parámetros genéticos y el método de mejoramiento genético más adecuado (Hallauer y Miranda, 1981), tanto en especies autógamas como alógamas.
Se denominan cruzas dialélicas a las cruzas simples que pueden lograrse entre los elementos de un conjunto básico de p líneas progenitoras (Griffing 1956a, b). Los diseños uno y tres de Griffing (1956a, b) sirven para estimar aptitud combinatoria general (ACG), aptitud combinatoria específica (ACE), efectos maternos (EM), efectos recíprocos (ER) y, en el caso en que se considere que los híbridos son una muestra aleatoria, componentes de varianza. Todo esto es importante para la toma de decisiones en programas de mejoramiento genético vegetal.
La aptitud combinatoria es la capacidad que tienen un individuo o una población de combinarse con otros, medida por medio de su progenie (Márquez, 1988). Sin embargo, la aptitud combinatoria debe determinarse no sólo en un individuo de la población sino en varios, a fin de poder seleccionar los que exhiban la más alta aptitud combinatoria. Existen varios diseños de análisis dialélico para estimar ACG y ACE, pero el más utilizado es el de Griffing (1956 b), en sus cuatro métodos: 1) progenitores y sus cruzas F1 directas y recíprocas; 2) progenitores y cruzas F1 directas; 3) cruzas F1 directas y recíprocas; y 4) cruzas F1 directas.
La progenie de las cruzas (F1) produce información sobre los progenitores al medir sobre ella los caracteres de interés. Los resultados se interpretan mediante un modelo lineal que incorpora ACG, ACE, EM y ER, según el método de Griffing empleado; lo cual permite la interpretación en términos de parámetros genéticos.
La reducción de los costos de inversión por compra de semilla es una estrategia sustentable que impactaría a la producción de calabacita e incrementaría los márgenes de ganancia para el productor, puesto que algunos híbridos comerciales podrían ser usados exitosa y ventajosamente en programas de mejoramiento genético vegetal para la formación de variedades sintéticas cuyos progenitores sean híbridos de cruza simple sobresaliente. Estos progenitores, idealmente deberían ser tales que: 1) no contribuyan con niveles importantes de endogamia; 2) la endogamia generada no esté fuertemente asociada con una fuerte depresión endogámica, y 3) contribuyan a que el rendimiento se mantenga dentro de niveles aceptables (Sahagún y Villanueva, 1997); por tal razón, los objetivos del presente estudio fueron: a) identificar híbridos comerciales con alto valor genético con base en los efectos de ACG y ACE, para producir sintéticos y variedades de alto rendimiento y b) evaluar el potencial productivo de las cruzas entre híbridos comerciales.
MATERIALES Y MÉTODOS
Progenitores
]]> Se emplearon ocho híbridos de calabacita tipo 'Grey Zucchini' de diferentes casas comerciales (Cuadro 1). Con ellos se obtuvieron 56 cruzas (28 directas y 28 recíprocas) posibles en la primavera de 2006. De cada cruza se aseguró al menos la polinización de 10 plantas del progenitor femenino.
Evaluación experimental
Las cruzas y los progenitores se evaluaron en dos ciclos en 2007 y 2008 en el Campo Agrícola Experimental de la Universidad Autónoma Chapingo. En 2007 la siembra se realizó el 15 de abril, y en 2008 el 20 de abril de forma directa; se depositaron dos semillas por mata para posteriormente ralear a una planta, bajo condiciones de riego. Se usó un diseño experimental de bloques completos al azar con tres repeticiones, donde la unidad experimental fue una parcela de dos surcos de 5 m de largo separados a 0.8 m con una separación entre plantas de 0.2 m (62,500 plantas·ha–1). La fertilización total fue de 120–80–00, aplicando 60–80–00 al momento de la siembra, y el resto de la fertilización nitrogenada se aplicó en el aporque. Las malezas se controlaron manualmente. Los caracteres estudiados fueron: peso de fruto por corte (PFR, g) con base en todos los frutos por corte de cada unidad experimental con una balanza granataria; número de frutos por corte (NFR) de un total de 17; largo y ancho de fruto (LFR, AFR, cm) de una muestra aleatoria de cinco frutos medidos con un vernier; días a floración masculina y femenina (DFM y DFF), registrados cuando existía un 50% de plantas con flores abiertas; rendimiento por planta por corte (g·planta–1), el cual se obtuvo como el cociente del peso de frutos por corte de un total de 17 entre el número de plantas por unidad experimental y rendimiento por hectárea (t·ha–1).
Diseño genético
Se empleó el Método I Modelo II de Griffing (1956 b), que incluye los progenitores y las cruzas directas y recíprocas, cuyo modelo lineal de análisis combinado de localidades, de acuerdo con Montesinos et al. (2007), es: Yijkl = μ+ πl+ τij + δk(l)+(πτ)ijl+ eijkl donde Yijkl es el valor fenotipico observado de la cruza (i, j) dentro del bloque k en el año l; m es la media general; πl es el efecto del año l; (πτ)ijl es el efecto del bloque k en el año l; δk(l) es el efecto de la interacción de la cruza (πτ)ijl con el año l; τij es el efecto de la cruza (i, j) y es igual a: τij = gi + gj + sij + mi – mj + rij; donde g, es el efecto de la ACG del progenitor i; gj es el efecto de la ACG del progenitor j; sij es el efecto de la ACE de la cruza (i, j); mi es el efecto materno del progenitor i; mj es el efecto materno del progenitor j; rij es el efecto recíproco de la cruza (i, j); e es el efecto aleatorio del error correspondiente a la observación (i, j, k, l).
De acuerdo con el criterio de agrupamiento propuesto por Serrano y Mendoza (1990), los mejores progenitores para formar una población base para mejoramiento por selección, serán aquellos con mayores efectos de ACG. El criterio de comparación de las cruzas F1 se basó en sus valores genotípicos, de modo que aparecen tres situaciones: 1) en cruzas con ACG = gi + gj > ACE se espera que no presenten depresión endogámica significativa en generaciones avanzadas (Fn, n>1), pudiendo utilizarse como variedades sintéticas cuando éstas sean de alto rendimiento; 2) con ACG < ACE, las cruzas mostrarían cambios aleatorios en su rendimiento en Fn, ya que los efectos específicos son de mayor magnitud que los aditivos en la expresión de rendimiento; así, de ser una cruza rendidora sólo deberá utilizarse como cruza y no como sintético, y 3) cuando ACG = ACE, ambos tipos de acción génica (aditividad y dominancia) son importantes por lo que la depresión endogámica en Fn se espera que sea reducida, pudiendo usarse la cruza como variedad sintética y como población base para selección recurrente.
Análisis estadístico
El análisis estadístico se realizó con el programa SAS versión 9.0 para microcomputadora. Se utilizó el algoritmo propuesto por Montesinos et al. (2007) para el análisis de varianza combinado por ambientes (Cuadro 2) (Martínez, 1983) del diseño I de Griffing. La prueba de significancia de los efectos principales (ACG, ACE, EM, ER) e interacciones (AmbxACG, AmbxACE, AmbxEM, AmbxER) se realizó mediante las pruebas de F que presenta Martínez (1983) en el análisis de varianza combinado del diseño I de Griffing.
]]> Componentes de varianza genéticaEn la estimación de los componentes de varianza con el Método I Modelo II de Griffing (efectos aleatorios en el caso en que se considere que los híbridos son una muestra de una población de híbridos), se asumió que los progenitores son híbridos de cruza simple formados por líneas no emparentadas y con coeficientes de endogamia igual a la unidad, por lo que los componentes de varianza de aptitud combinatoria general (σ2g) y de aptitud combinatoria específica (σ2s) de la población de referencia como factores de efectos aleatorios, tienen las siguientes equivalencias en términos de covarianza [Cov] de familias de medios hermanos (MH) y hermanos completos (HC) (Cockerham, 1954 y Kempthorne, 1957):
σ2ACG= Cov MH; σ2ACE= Cov HC – 2 Cov MH.
Lo anterior tiene, a su vez, las siguientes equivalencias en términos de varianza genética aditiva (σ2A) y varianza genética de dominancia (σ2D):
Cov MH = ((1 + F)/4) σ2A ; Cov HC = ((1 + F)/2) σ2A+ ((1 + F)/2) σ2D; asumiendo (en nuestro caso) que F=1 la Cov MH = 1/2 σ2A y Cov HC = σ2A+ σ2D.
Finalmente, los componentes de varianza en términos de varianzas genéticas son: σ2g = 1/2 σ2A por lo tanto la σ2A= 2σ2ACG; la σ2S = Cov HC – 2 Cov MH y σ2S= σ2A+ σ2D – 2(1/2σ2A) de esta manera la σ2D= σ2ACE.
Con base en el análisis de varianza combinado y a partir de las esperanzas de los cuadrados medios (Cuadro 2), se demuestra que:
 donde:
donde:

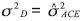





A partir de la varianza genética aditiva ( 2A = 22ACG) y la varianza genética de dominancia (2D = 22ACE), se calcularon los valores de heredabilidad en sentido estrecho.
2A = 22ACG) y la varianza genética de dominancia (2D = 22ACE), se calcularon los valores de heredabilidad en sentido estrecho.

para los siete caracteres, donde (2F) es la varianza fenotípica, la cual se obtuvo a partir de los componentes de varianza estimadas (σ2ACG,σ2ACE,σ2EM, σ1ER,σ1AMBxACG, σ2AMBxACE, σ2AMBxEM, σ2AMBxER, σ2EC) por el modelo del análisis combinado de Montesinos et al. (2007) de la siguiente forma:

y el grado promedio de dominancia
]]> (Comstock y Robinson, 1952).
(Comstock y Robinson, 1952). Se realizó el cálculo de la proporción de los efectos de ACG , ACE, EM y ER. Éstos se calcularon con base en las sumas de cuadrados, con respecto a la proporción que ocupan cuando las cruzas se dividen en estos efectos.
RESULTADOS Y DISCUSIÓN
Análisis de varianza
Los cuadrados medios combinados por ambientes y los niveles de significancia para los caracteres: rendimiento por planta (RPP, g), número de frutos por planta (FPP), largo de fruto (LFR, cm), ancho de fruto (AFR, cm), días a floración masculina (DFM), días a floración femenina (DFF) y rendimiento de calabacita para verdura por hectárea (REN, t·ha–1) con el método I modelo II de Griffing, que se presentan en el Cuadro 3, indican que hubo diferencias altamente significativas (P < 0.01) entre cruzas para todos los caracteres evaluados, lo cual se puede atribuir a la divergencia genética que presentan los progenitores por su diversidad de origen. Esto hace posible la identificación de cruzas precoces y tardías, con frutos más grandes (largos y anchos) y sobre todo con rendimiento contrastante. La contribución a la varianza del rendimiento por planta atribuible a las cruzas, estuvo constituida por 42% para los efectos de ACG, 45% no aditivos (ACE), 5% efectos maternos y 8% efectos recíprocos. Los resultados con respecto a los efectos de ACG y ACE, indican que tanto los efectos aditivos como los no aditivos contribuyen de manera equilibrada en las cruzas para rendimiento. Al respecto, Gutiérrez et al. (2002) encontró en maíz que a medida que la divergencia genética de los materiales se incrementa, aumenta también la diferencia entre los valores de Aptitud Combinatoria, ya sea para ACG o para ACE; o bien para los dos tipos de acción génica.
Se encontraron mayores efectos del tipo aditivo (61.59% para efectos de ACG y 26.95% para ACE) en largo de fruto; 72.67 y 14.91% para ACG y ACE, respectivamente, en ancho de fruto; 88.08 y 7.58% de ACG y ACE, respectivamente; en días a floración masculina y en días a floración femenina los efectos de ACG fueron 87.37% y de ACE 7.83%. Por otra parte, la mayor contribución a la varianza del número de frutos por planta atribuible a las cruzas se debió principalmente a los efectos de dominancia (ACE) = 52.26%, superando a los efectos aditivos (ACG) = 29.16%; lo anterior probablemente se debió a la heterosis resultante de las combinaciones híbridas entre los progenitores.
Existieron diferencias altamente significativas (P<0.01) de ACG entre híbridos progenitores para LFR, AFR, DFM y DFF, no así para RPP y FPP. En aptitud combinatoria específica existieron diferencias altamente significativas entre híbridos progenitores en RPP, FPP, LFR y REN, y no se encontraron diferencias en las variables AFR, DFM y DFF. Este comportamiento se atribuye a una baja acumulación aditiva de genes para rendimiento en los cruzamientos. No obstante, en algunas variables existieron efectos aditivos.
Los efectos maternos (EM) no fueron significativos (Cuadro 3), es decir, los caracteres evaluados están influidos por genes de cromosomas localizados en el núcleo, y no en el citoplasma. Sin embargo, en los efectos recíprocos (ER) se encontraron diferencias significativas en RPP y REN, lo cual es indicativo de que hay cruzas en que los progenitores no se comportan de igual manera en la cruza directa que en la cruza recíproca, aunque estas diferencias no se pueden atribuir a los efectos maternos de cada progenitor, sino a efectos de interacción entre genes del núcleo y del citoplasma.
A pesar de la significancia de algunas interacciones (AmbxACE, AmbxEM y AmbxER), su contribución a la varianza fenotípica observada para las diferentes características resultó pequeña en comparación con la contribución de los efectos de ACG y ACE (Cuadro 3).
]]> Componentes de varianza genéticaEn la población teórica obtenida de la recombinación de las cruzas F1 entre los híbridos comerciales progenitores, se encontró que existe varianza genética aditiva y de dominancia. La varianza genética aditiva resultó importante en los caracteres largo y ancho de fruto, y días a floración masculina y femenina, por lo que es factible mejorarlos por selección recurrente, aprovechando que presentan valores medios de heredabilidad (42.11, 49.78, 53.88 y 54.45%, respectivamente); mientras que en número de frutos por planta, rendimiento por planta y por hectárea fueron bajos (25.10, 10.95 y 25.48%, respectivamente) (Cuadro 4). Los resultados anteriores se reafirman al analizar los coeficientes de variación genética aditiva (CVA) y de dominancia (CVD). Se observó que para largo y ancho de fruto, así como para días a floración masculina y femenina, el CVA fue mayor que el CVD, lo que confirma la importancia de los efectos aditivos para estos caracteres. Sin embargo, para frutos por planta, rendimiento por planta y por hectárea que son los principales componentes del rendimiento, el CVD fue mayor que el CVA (Cuadro 4). Un aspecto importante de estos dos coeficientes es que son comparables entre caracteres, y con ellos se puede predecir el éxito esperado en la selección. Así, los caracteres frutos por planta, rendimiento por planta y por hectárea mostraron alta variabilidad genético–aditiva; días a floración masculina y femenina mostraron variabilidad intermedia y largo y ancho de fruto mostraron la mínima variabilidad (Cuadro 4).
El grado promedio de dominancia ( ) indicó sobredominancia en los caracteres RPP (1.6733), FPP (2.4600) y REN (1.6761), mismos que pueden mejorarse por hibridación, ya que teóricamente la F1 superaría significativamente al genotipo homocigoto dominante. Al respecto Hallauer y Miranda (1981) indican que los valores mayores a la unidad en la estimación del grado promedio de dominancia, proporcionan información para explotar la heterosis, y que el mayor vigor híbrido se debe a la presencia en el heterocigote (F1) de un número mayor de genes dominantes que en los progenitores. El largo y ancho de fruto, días a floración masculina y femenina presentan dominancia parcial.
) indicó sobredominancia en los caracteres RPP (1.6733), FPP (2.4600) y REN (1.6761), mismos que pueden mejorarse por hibridación, ya que teóricamente la F1 superaría significativamente al genotipo homocigoto dominante. Al respecto Hallauer y Miranda (1981) indican que los valores mayores a la unidad en la estimación del grado promedio de dominancia, proporcionan información para explotar la heterosis, y que el mayor vigor híbrido se debe a la presencia en el heterocigote (F1) de un número mayor de genes dominantes que en los progenitores. El largo y ancho de fruto, días a floración masculina y femenina presentan dominancia parcial.
Análisis de progenitores
Los efectos de ACG de los híbridos comerciales utilizados como progenitores y la varianza de los efectos de ACE a través de sus cruzas en promedio de ambientes, se presentan en el Cuadro 5.
Con base en los efectos de ACG para rendimiento por planta y por hectárea, los mejores progenitores fueron en orden decreciente: Terminator (RPP = 0.121 g·planta–1) y REN = 3.788 t·ha–1), Lolita (RPP = 0.113 g·planta–1 y REN = 3.545 t·ha–1) y WA9041 (RPP = 0.056 g·planta–1 y REN = 1.765 t·ha–1). Es importante resaltar el hecho de que los progenitores Lolita y WA9041 tuvieron valores negativos de ACG para DFF (–1.812 y –2.250 días, respectivamente), lo que indica que sus cruzas son más precoces; por su parte, Terminator presentó valores positivos de ACG para DFF (3.843 días). La existencia de efectos positivos de ACG en días a floración, es una situación desfavorable si se deseara mejorar dicho carácter hacia precocidad, porque tales efectos implican ciclos más tardíos. Los peores progenitores fueron: Grey Zucchini de seminis (RPP = –0.102 g·planta–1 y REN = –3.211 t·ha–1) y Dolarzini (RPP = –0.068 g·planta–1 y REN = –2.150 t·ha–1), según los efectos de ACG.
Lolita y WA9041 presentaron efectos positivos de ACG para largo de fruto (0.285 y 0.324 cm, respectivamente) y negativos para ancho de fruto (–0.023 y –0.001 cm, respectivamente) (Cuadro 5); dichos resultados indican que estos dos progenitores producen frutos más largos que anchos.
Terminator ocupó el primer lugar en orden decreciente en σ2ACE = 0.0697 en rendimiento por planta y por hectárea (σ2ACE = 68.0826), pudiendo interpretarse como de baja estabilidad genética a través de sus cruzas; por su parte, Lolita ocupó el cuarto lugar (σ2ACE = 0.0269 en RPP y 26.2875 en REN), considerándose como de estabilidad genética media a través de sus cruzas al igual que WA9041 que se ubicó en el quinto lugar (σ2ACE = 0.0246 en RPP y 24.0711 en REN) (Cuadro 5).
Los híbridos que resultaron ser los mejores progenitores con base en la magnitud de sus efectos positivos de ACG fueron, en orden decreciente, Terminator, Lolita y WA9041. De éstos, el más consistente a través de sus cruzas fue WA9041; mientras que los más inconsistentes fueron Tala y Hurakán, por su varianza de ACE a través de las cruzas en que participan (Cuadro 5).
Análisis de cruzas
]]> Se presentan las cruzas de mayor interés por la magnitud de los efectos genotípicos (ACG y ACE) para rendimiento de calabacita para verdura en promedio de ambientes (Cuadro 6). Se discuten las seis primeras en orden decreciente para REN y que son estadísticamente similares:1) (Hurakán x Lolita) i = 5, j = 4 con ACG = 2.6505 < ACE = 11.2145 y con un REN = 52.16 t·ha–1 en la cruza directa y 48.87 t·ha–1 en la cruza recíproca, por lo que la cruza se puede utilizar empleando como progenitor femenino a cualquiera de los dos híbridos en cuestión. Predomina la ACE, por lo que sólo es utilizable como cruza, ya que se espera depresión endogámica en Fn.
2) (WA9041 x Lolita) i = 6, j = 4 con ACG = 5.3113 > ACE = 2.1637 y REN = 51.45 t·ha–1, por lo que puede utilizarse directamente como variedad de polinización libre sin detrimento del rendimiento en generaciones avanzadas (ACG > ACE).
3) (Lolita x Tala) i = 4, j = 1 con ACG = 2.3218 < ACE = 7.3529 y REN = 50.14 t·ha–1, utilizable solamente como cruza, ya que ACE > ACG.
4) (Grey zucchini M+M x Terminator) i = 3, j = 7 con ACG = 2.1693 > ACE = –6.5370 y REN = 50.00 t·ha–1; esta cruza se puede utilizar directamente como variedad de polinización libre sin detrimento del rendimiento en generaciones avanzadas (ACG > ACE) y como población base para selección recurrente.
5) (Tala x Terminator) i = 1, j = 7 con ACG = 2.5647 < ACE = 8.4335 y REN = 47.72 t·ha–1. Se recomienda su uso únicamente como cruza.
6) (Lolita x Terminator) i=4, j=7 con ACG = ACE (7.33 t·ha–1) y REN = 44.09 t·ha–1 por lo que en esta cruza son igualmente importantes tanto los efectos aditivos como los de dominancia, pudiéndose utilizar como variedad sintética y como población base para selección recurrente.
Nótese que en las seis cruzas con rendimiento numéricamente más alto se encuentra presente en cuatro de ellas el híbrido Lolita y en tres el híbrido Terminator, que fueron los progenitores con mayores efectos de ACG (3.545 y 3.788 t·ha–1, respectivamente) (Cuadro 5).
De estos resultados resalta el hecho de que tres de las seis mejores cruzas evaluadas resultaron entre híbridos de casas comerciales diferentes, superando a los híbridos comerciales progenitores en seis toneladas por hectárea, pudiéndose interpretar que se debe a combinación de germoplasma genéticamente contrastante (divergencia genética); y las otras tres cruzas más rendidoras se obtuvieron entre híbridos progenitores de la misma compañía semillera, tal vez porque tienen diferente información genética. El alto rendimiento de una cruza puede deberse a la suma de efectos aditivos de los genes de ambos progenitores, o bien a los efectos de interacción de los alelos dominantes de un progenitor con los alelos recesivos del otro progenitor (Falconer, 1985).
Efectos maternos y recíprocos
]]> Los efectos maternos positivos para rendimiento por planta y por hectárea de los híbridos Lolita, Hurakán y WA9041, indican que tales progenitores tienen mayor rendimiento en sus cruzas cuando son usados como progenitor femenino, que en sus respectivas cruzas recíprocas. Los valores negativos de efectos maternos indican lo contrario (Cuadro 7). WA9041 resultó ser un buen progenitor femenino por tener efectos maternos positivos en frutos por planta (0.3958 frutos), rendimiento por planta (0.075 g·planta–1) y por hectárea (2.2658 t·ha–1); mostró efectos maternos negativos en días a floración femenina y masculina, lo que indica que sus cruzas directas son más precoces. Los híbridos Lolita y Hurakán tuvieron un comportamiento similar a WA9041 en efectos maternos positivos para frutos por planta (0.313 y 0.5938 frutos, respectivamente) y por hectárea (0.4453 y 1.3953 t·ha–1, respectivamente); sin embargo, Lolita presentó valores negativos para días a floración femenina (–0.2187 d) y Hurakán positivos (0.5521 d), resultados que indican que las cruzas directas del progenitor Lolita son más precoces y las de Hurakán más tardías.En los caracteres largo y ancho de fruto, los efectos maternos fueron positivos en los progenitores Grey Zucchini de Seminis (0.0198 y 0.0667 cm, respectivamente) y Dolarzini (0.1042 y 0.0042 cm, respectivamente), lo que significa que las cruzas directas producen frutos más grandes (largos y anchos) que en sus respectivas cruzas recíprocas. Los progenitores Grey Zucchini M+M y Terminator mostraron efectos maternos positivos para largo de fruto (0.1458 y 0.1365 cm, respectivamente), por lo que sus cruzas directas producen frutos más largos que sus respectivas cruzas recíprocas.
Las cruzas WA9041 x Grey zucchini M+M i=6, j=3; Hurakán x Dolarzini i=5, j=8; Lolita x Tala i=4, j=1 y WA9041 x Lolita presentaron efectos recíprocos para rendimiento, es decir, las interacciones entre el ADN nuclear y el citoplásmico no tienen el mismo efecto fenotípico en la cruza directa que en la recíproca. Por los efectos maternos, en la cruza directa tuvieron una media de 44.85, 42.64, 50.1 y 51.45 t·ha–1, respectivamente, mientras que en la cruza recíproca las medias fueron 28.34, 31.56, 41.50 y 43.53 t·ha–1 (Cuadro 6). Cabe indicar que las cruzas WA9041 x Lolita y Lolita x Tala i=4, j=1 fueron la segunda y tercera cruzas más rendidoras por planta y por hectárea cuando se realizó la cruza directa, por lo que se debe tener cuidado al realizar la cruza para mantener altos rendimientos por planta y por hectárea.
CONCLUSIONES
En la población teórica obtenida de la recombinación de las cruzas F1 entre híbridos comerciales de calabacita, es más importante la varianza de dominancia para los principales componentes del rendimiento (frutos por planta, largo de fruto y rendimiento por planta); sin embargo, en largo y ancho de fruto, así como en días a floración masculina y femenina, es más importante la varianza aditiva. Por lo tanto, el método de mejoramiento apropiado en la población de referencia sería por hibridación con altas posibilidades de obtener cruzas superiores a los progenitores.
Los híbridos que resultaron ser los mejores progenitores, a juzgar por la magnitud de sus efectos positivos de ACG, fueron, en orden decreciente: Terminator, Lolita y WA9041. De ellos, el más consistente a través de sus cruzas fue WA9041.
Terminator y Lolita fueron los mejores híbridos comerciales progenitores para rendimiento por hectárea de calabacita (44.23 t·ha–1 y 42.20 t·ha–1, respectivamente), y el peor fue Tala (21.92 t·ha–1).
De las seis mejores cruzas de rendimiento de calabacita para verdura, tres podrían utilizarse sólo como híbridos (ACG < ACE): Hurakán x Lolita (52.16 t·ha–1); Lolita x Tala (50.14 t·ha–1); Tala x Terminator (47.72 t·ha–1); dos directamente como variedades de polinización libre sin detrimento del rendimiento en generaciones avanzadas (ACG > ACE): WA9041 x Lolita (51.45 t·ha–1); Grey Zucchini M+M x Terminator (50.00 t·ha–1) y Lolita x Terminator (44.09 t·ha–1) como población base para mejoramiento por selección recurrente, ya que los efectos aditivos y de dominancia son importantes en la cruza (ACG = ACE).
]]> LITERATURA CITADA
COCKERHAM, C. C. 1954. An Extension of the concept of partitioning hereditary variance for analysis of cavariance among relatives when epistasis is present. Genetics. 39: 859–882. [ Links ]
COMSTOCK, R. E.; ROBINSON, F. H. 1952. Estimation of Average of Dominance of Genes. Heterosis.. Ames, Iowa State College Press. pp 494–516. [ Links ]
FALCONER, D. S. 1985. Introducción a la Genética Cuantitativa. Editorial CECSA, México. 135 p. [ Links ]
GRIFFING V. 1956a. A generalized treatment of the use of diallel crosses in quantitative inheritance. Heredity 10: 31–50. [ Links ]
GRIFFING V. 1956b. Concept of general and specific combining ability in relation to diallel crossing system. Austr. J. Biol. Sci. 9: 463–493. [ Links ]
GUTIÉRREZ DEL RÍO, E.; PALOMO G. A.; ESPINOZA, B. A.; DE LA CRUZ L. E. 2002. Aptitud combinatoria y heterosis para rendimiento de líneas de maíz en la comarca lagunera, México. Revista Fitotecnia Mexicana Vol 25(3): 271–277. [ Links ]
HALLAUER, A. R.; MIRANDA, J. B. 1981. Quantitative Genetic in Maize Breeding. Iowa State University Press. Ames, Iowa. pp: 268–368. [ Links ]
KEMPTHORNE, O. 1957. An Introduction to Genetics Statistics. John Wiley And Sons. New York. 545 p. [ Links ]
MÁRQUEZ S., F. 1988. Genotecnia Vegetal. Tomo II. Métodos, Teoría, Resultados. AGT Editor. México. 665 p. [ Links ]
MARTÍNEZ G., A. 1983. Diseño y Análisis de los Experimentos de Cruzas Dialélicas. Centro de Estadística y Cálculo, Colegio de Postgraduados, Chapingo, México. 249 p. [ Links ]
MONTESINOS L., O. A.; MASTACHE L., A. A.; LUNA E. I.; HIDALGO C., J. V. 2007. Mejor predictor lineal e insesgado combinado de los diseños uno y tres de Griffing. Téc. Pecu. Méx. Vol 45(2): 131–146. [ Links ]
SAHAGÚN C., J.; VILLANUEVA V., C. 1997. Teoría de las variedades sintéticas formadas con híbridos de cruza simple. Revista Fitotecnia Mexicana. Vol 20:69–79. [ Links ]
SERRANO C., L.; MENDOZA O., L.1990. Formación de híbridos de sorgo para grano III. Proposición de un modelo de selección de progenitores con base en sus parámetros genéticos. Revista Fitotecnia Mexicana Vol 13: 44–55. [ Links ]
]]>