
Hábitos alimenticios de Floridichthys polyommus Hubbs, 1936 (Pisces: Cyprinodontidae) en dos sistemas lagunares costeros
Feeding habits of Floridichthys polyommus Hubbs,1936 (Pisces: Cyprinodontidae) in two coastal lagoon systems
Alicia Virginia Poot Salazar*, Walter Gabriel Canto Maza, María Eugenia Vega-Cendejas
Laboratorio de Taxonomía y Ecología de Peces, CINVESTAV-IPN, Unidad Mérida, km 6 antigua carretera a Progreso. AP 73 "Cordemex". 97310 Mérida, Yucatán. E. mail: apoot@mda.cinvestav.mx
]]> Recibido: 4 de octubre 2004.
Resumen
Se determinaron y compararon espacial y estacionalmente los hábitos alimenticios de Floridichthys polyommus (Pisces: Cyprinodontidae) en dos sistemas lagunares de la península de Yucatán. Se analizaron los contenidos estomacales de 445 y 561 ejemplares correspondientes a las lagunas de Puerto Viejo (Quintana Roo) y Chelem (Yucatán), respectivamente. Esta especie presenta una amplia generalización trófica, incluyendo en su dieta 126 componentes alimenticios en Puerto Viejo y 80 en Chelem. Consume principalmente microcrustáceos, fitoplancton y macrófitas. Sin embargo, las preferencias alimenticias fueron diferentes entre ambos sistemas, tanto espacial como estacionalmente. En Puerto Viejo F. polyommus es un consumidor de microcrustáceos, mientras que en Chelem es fitoplanctófago. Considerando su variación trófica ontogenética, los ejemplares de menor talla se alimentaron principalmente de microcrustáceos y los más grandes de fitoplancton o macrófitas. En Puerto Viejo, esta especie consume principalmente microcrustáceos (35.1%) durante la época de secas, mientras que en lluvias y nortes, el fitoplancton es el más representativo. Este último grupo trófico fue en Chelem el de mayor abundancia durante las tres épocas del año y su valor se incrementó en la época de secas (51.6%). La variación trófica espacial y estacional entre ambas lagunas puede relacionarse con la disponibilidad de los recursos alimenticios en cada sistema, así como con los cambios hidrológicos en cada época climática.
Palabras clave: Floridichthys polyommus, hábitos alimenticios, variación trófica estacional, Laguna de Puerto Viejo, Laguna de Chelem.
Abstract
The feeding habits of Floridichthys polyommus (Pisces: Cyprinodontidae) were determined and compared spatially and seasonally between two coastal lagoon systems in Yucatan Peninsula. A total of 445 and 561 stomach contents were analyzed from the lagoons of Puerto Viejo (Quintana Roo) and Chelem (Yucatan), respectively. This specie presents a wide trophic generalization, including 126 alimentary items in Puerto Viejo and 80 in Chelem. It consumes basically microcrustaceans, phytoplankton, and macrophytes. However, trophic preferences were different spatially and seasonally between both systems. In Puerto Viejo F. polyommus feeds on microcrustaceans, whereas in Chelem it consumes phytoplankton. Considering its trophic ontogenetic variation, smaller individuals preyed on microcrustaceans and the larger ones on phytoplankton or macrophytes. In Puerto Viejo, microcrustaceans were the principal and more abundant food during the dry season (35.1%), while during the rainy and cold seasons phytoplankton was the most representative item. Phytoplankton was always the most abundant food in Chelem all year long, with an increase in the dry season (51.6%). Spatial and seasonal trophic variation between both lagoons can be related to the food components availability in each system and to hydrological variability for each season of the year.
Key words: Floridichthys polyommus, feeding habits, seasonal trophic variation, Puerto Viejo lagoon, Chelem lagoon.
]]>Introducción
La información sobre dietas, disponibilidad del alimento y conducta alimenticia de los peces es fundamental para entender la estructura de sus comunidades, patrones de distribución y estrategias de sus historias de vida (Blaber, 1997).
Los peces, en contraste con la mayoría de otros grupos faunísticos, presentan gran plasticidad trófica y muestran diferentes hábitos alimenticios en relación con la disponibilidad del alimento y ontogénicamente como respuesta a sus requerimientos fisiológicos y nutricionales (Wootton, 1990). Los hábitos alimenticios pueden cambiar aún en una misma especie de acuerdo a la localidad, disponibilidad del alimento, la estabilidad, la edad y/o el sexo (Prejs & Colomine, 1981; Blaber, 1997).
No obstante que las especies de la familia Cyprinodontidae no presentan una importancia comercial, su papel ecológico es fundamental en la transformación y transporte de energía al interior del ecosistema, al constituir recursos alimenticios para especies de niveles tróficos superiores (Vega-Cendejas, 1998). Las especies pertenecientes a esta familia habitan principalmente aguas templadas, someras y de altas salinidades. El conocido "bolin yucateco" (Floridichthys polyommus), se encuentra incluso en aguas casi anóxicas, temperaturas de hasta 38ºC y salinidades mayores de 100 ppm (Vega-Cendejas et al., 1994).
Estudios realizados por Vega-Cendejas et al. (2000) y Vega-Cendejas y Hernández (2004) sobre las comunidades de peces de la laguna Puerto Viejo del Parque Nacional Isla Contoy y del sistema lagunar de la Reserva de Río Lagartos reportan a F. polyommus como una de las especies de mayor abundancia en ambos sistemas costeros. Considerando la abundancia de esta especie y la carencia de información sobre su biología, el presente trabajo tiene como objetivo caracterizar sus hábitos alimenticios y determinar sus diferencias tróficas entre ambos sistemas lagunares y a nivel estacional.
Materiales y métodos
La laguna Puerto Viejo se encuentra en la parte central de Isla Contoy, en el Estado de Quintana Roo (21º30'N, 86º48'W). Alberga tres islotes en su interior y se caracteriza por presentar aguas cálidas de alta salinidad (39) con altos niveles de oxígeno disuelto y silicatos. Presenta una profundidad de 1.3 m durante secas y 1.1 m en lluvias y nortes (Fig. 1).
]]>
La laguna de Chelem se ubica al noreste de las costas del Estado de Yucatán, a 30 km. al norte de la ciudad de Mérida (21º15-17'N, 89º39-48' W). Tiene una superficie aproximada de 15 km2, y una longitud y anchura de 14.7 y 1.8 km, respectivamente. Presenta una boca artificial con 225 m de ancho y una profundidad que varía entre 0.5 y 1.5 m, excepto en la zona de la dársena, que alcanza los 3.0 m (Fig. 1).
En ambos sistemas lagunares las colectas de los peces se realizaron bimensualmente, de abril de 1999 a enero de 2000 en la laguna Puerto Viejo y de marzo de 2002 a enero de 2003 en la laguna de Chelem, utilizando un chinchorro playero de 15 m de largo por 1.5 m de caída y apertura de malla de 1 pulgada. Los ejemplares colectados se preservaron en formaldehído al 30% y fueron transportados al laboratorio para su análisis.
Se registró el peso y longitud estándar de cada uno de los ejemplares. Posteriormente se extrajo el contenido estomacal y se observó al microscopio estereoscópico y óptico. Los componentes obtenidos se identificaron hasta el nivel taxonómico más bajo posible según el grado de digestión, con base en bibliografía especializada.
Los componentes en la dieta se cuantificaron según las tallas y las épocas del año, utilizando los métodos de Porcentaje por Área (PA) y de Frecuencia de Ocurrencia (FO) (Tresierra & Culquichicom, 1995). Para estimar la variación trófica ontogénica se establecieron clases de longitud de 0.9 cm, obteniendo en total 8 intervalos o clases de talla. El método de PA es una modificación del método volumétrico, utilizado cuando el contenido estomacal está constituido por presas muy pequeñas (diatomeas, copépodos, ostrácodos) y cuyo peso no es posible registrar en una balanza analítica (Vega-Cendejas, 1990). Consiste en distribuir uniformemente el contenido estomacal sobre una superficie milimétrica y estimar el área ocupada por éste como el 100%; el área total de cada componente alimenticio se representa como un porcentaje del área total de todos los contenidos estomacales. Este método proporciona una medida representativa de la biomasa y puede aplicarse a todos los componentes alimenticios, incluyendo los más pequeños (Hyslop, 1980).
El método de FO consiste en contar el número de veces que aparece una presa determinada en el contenido estomacal de un grupo de peces y expresarla como porcentaje del número de estómagos analizados (Tresierra & Culquichicom, 1995):
% FO = (n/NE)(100)
En donde:
FO= frecuencia de ocurrencia,
n= número de estómagos con cierto componente alimenticio,
]]> NE= número total de estómagos analizados.Este método fue utilizado como complemento del PA para precisar si un componente dado es accidental o es el alimento preferencial en la dieta del depredador (Hyslop, 1980). Según la FO, los componentes se clasificaron en tres categorías: a) preferenciales, que caracterizan al régimen alimenticio, si FO>50%, b) secundarios, aquellos que son consumidos en cierta etapa de la ontogenia y que son considerados como los que sustituyen a los anteriores cuando estos faltan, cuando su FO >10% y <50%. Finalmente fueron considerados como c) accidentales aquellos componentes alimenticios cuyo consumo fue muy escaso o raro (<10%) (Yáñez-Arancibia, 1975).
Resultados
Se analizaron 445 ejemplares de la laguna Puerto Viejo (0.9 a 7.0 cm de LE) y 561 provenientes de la laguna de Chelem (1.1 a 9.0 cm de LE). El número de individuos analizados por época climática varió entre ambos sitios (Tabla 1); sin embargo, siempre fue mayor a 85 individuos, de los cuales para ambos sitios el 100% se encontraban con el estómago medio lleno o lleno.
Los resultados obtenidos muestran que esta especie presenta un amplio grado de generalización trófica, incluyendo en su dieta 126 componentes alimenticios en la laguna Puerto Viejo y 80 en la laguna de Chelem. Los taxa identificados en los estómagos se agruparon en grupos tróficos para un mejor manejo. Como fitoplancton se consideraron a las bacilariofitas, clorofitas, cianofitas, rodofitas y microalgas que no pudieron identificarse. Dentro de los microcrustáceos quedaron comprendidos los anfípodos, copépodos, isópodos, ostrácodos, tanaidaceos, misidáceos y crustáceos no identificados. En los protozoarios se incluyeron los dinoflagelados, foraminíferos, oligotríquidos, trofozoítos y protozoarios no identificados. En los moluscos se agruparon los bivalvos, gasterópodos y poliplacóforos. Como macrófitas, se incluyeron algas y semillas de algas observables en el microscopio estereoscópico y pastos marinos. En los Porifera se incluyeron restos de esponjas y sus espículas. Se consideró como detritus al material alimenticio que debido a su consistencia, grado de digestión y presencia de arena sugiere que es materia orgánica en descomposición. Dentro de los Annelida se consideraron a los poliquetos. En Echinodermata se incluyó a la familia Ophiuroidea. En la categoría "otros" se incluyeron componentes cuya representatividad fue baja, tales como ácaros, huevecillos, larvas de insectos o insectos adultos, restos de peces o escamas, MANI (material animal no identificado) y MONI (material orgánico no identificado).
En la laguna Puerto Viejo los grupos tróficos más abundantes en la dieta fueron microcrustáceos (32.1%), fitoplancton (26.4%), protozoarios (15.3%) y detritus (10.5%). En relación a su variación trófica ontogénica, los individuos con intervalos de tallas de 1.1-2.0 cm de LE consumieron más del 50% de microcrustáceos y protozoarios, y más del 30% de fitoplancton y detritus. Conforme los ejemplares incrementaron su talla (2.1-3.0 a 6.1-7.0 cm de LE) consumieron preferentemente microcrustáceos y fitoplancton (>45%). En el intervalo de tallas de 6.1-7.0 cm aumenta el consumo de fitoplancton a más de 40%. Con respecto a los demás grupos tróficos cabe resaltar que hubo un incremento en el consumo de macrófitas, así como una disminución de detritus con el incremento de talla.
En la laguna de Chelem, los grupos tróficos más abundantes en la dieta fueron fitoplancton (39.4%), macrófitas (21.1%), microcrustáceos (10.3%) y protozoarios (10.4%). A través de su crecimiento, los individuos con tallas de 1.1-2.0 cm de LE consumieron más del 50% de fitoplancton y el 30% de microcrustáceos y protozoarios, mientras que los ejemplares de 2.1-3.0 y 3.1-4.0 cm de LE consumieron más del 45% de fitoplancton y más del 12% de protozoarios. Conforme incrementaron su talla (4.1-5.0 a 7.1-8.0 cm) consumieron preferentemente fitoplancton y macrófitas (>60%). En el intervalo de tallas de 8.1-9.0 cm el consumo de fitoplancton aumentó a más de 70%. Con respecto a los demás grupos tróficos, hubo una disminución en el consumo de detritus y un incremento en el de moluscos con el crecimiento en talla.
]]> Secas. En la laguna Puerto Viejo se analizaron 273 ejemplares correspondientes a los meses de abril y junio, con tallas de 1.0 a 7.0 cm de LE. Los grupos tróficos más abundantes en la dieta durante esta época del año fueron los microcrustáceos (35.1%), fitoplancton (22.5%), protozoarios (15.0%) y detritus (8.6%) (Tabla 2). Los componentes preferenciales (FO>50%) fueron 4 y los secundarios 27 con FO de 10.3% a 48.4% (Tabla 3). El resto de los componentes alimenticios con FO por debajo del 10% se consideraron como accidentales (80 componentes). Durante el crecimiento de esta especie en la época de secas, los ejemplares más pequeños (1.1-2.0 cm de LE) consumen principalmente protozoarios (33.7%) y microcrustáceos (17.4%), conforme aumentan de talla (2.1-3.0 cm a 5.1-6.0 cm de LE), el espectro trófico cambia hacia un mayor consumo de fitoplancton (32.7%) y microcrustáceos (37.8%). En los individuos con intervalo de tallas de 6.1-7.0 cm, el consumo de macrófitas en la dieta fue casi del 60%, lo cual indica una tendencia de esta especie a ser especialista en esta etapa de su ontogenia durante esta época del año (Fig. 2A).
En la laguna de Chelem, se analizaron 91 ejemplares de esta especie que corresponden al mes de marzo, con tallas de entre 2.4 a 7.0 cm de LE. Los grupos tróficos más abundantes durante esta época del año fueron el fitoplancton (51.6%), macrófitas (14.3%), protozoarios (13.1%), detritus (10.8%) y microcrustáceos (7.1%) (Tabla 2). Los componentes preferenciales fueron 12, las presas secundarias 20 con FO desde 11.0% a 48.4% (Tabla 3). El resto de los componentes alimenticios se consideraron como accidentales (25 componentes). En ejemplares juveniles de 2.1 a 3.0 cm de LE esta especie consumió principalmente fitoplancton (47%), protozoarios (20%), microcrustáceos (15%) y detritus (11%). En los intervalos de tallas de 3.1 a 4.0 cm se observa un aumento en el consumo de detritus y macrófitas con el 14 y 10% respectivamente, disminuyendo el consumo de microcrustáceos (6%). Conforme esta especie incrementa de talla de 5.1 a 7.0 cm su preferencia alimenticia se vuelve más especializada hacia el consumo de fitoplancton (54%) y macrófitas (18%), manteniéndose constante el consumo de microcrustáceos, detritus y protozoarios, contribuyendo con el 21% del contenido estomacal (Fig. 2B).
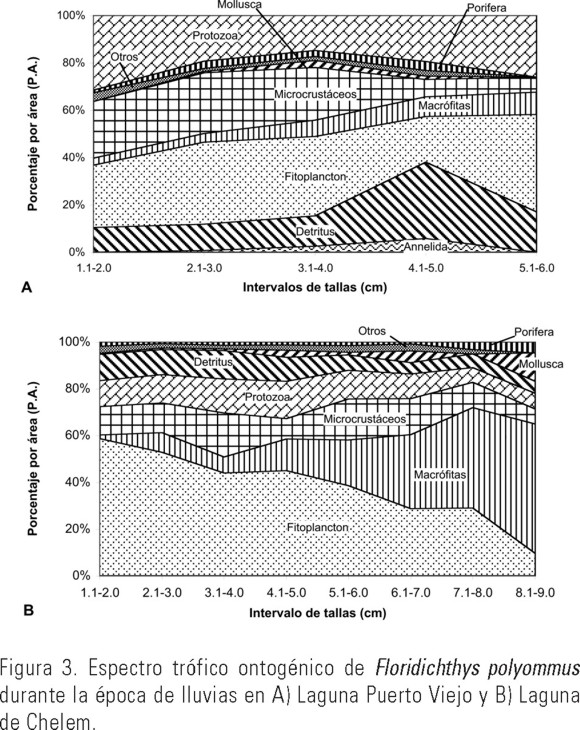
Lluvias. En la laguna Puerto Viejo se analizaron 85 ejemplares correspondientes a los meses de julio y septiembre (1.3 a 5.8 cm de LE). Los grupos tróficos más abundantes durante esta época fueron el fitoplancton (32.7%) microcrustáceos (23.2%), protozoarios (18.8%) y detritus (12.6 %) (Tabla 2). Los componentes preferenciales fueron 9 y los secundarios 21 con FO desde 11.8% a 48.2% (Tabla 4). El resto de los componentes alimenticios (41) fueron accidentales. A través de los diversos intervalos de talla analizados para esta especie durante esta época del año, se registró en los ejemplares mas pequeños y los de mayor talla un consumo preferencial de protozoarios (31.5%) y fitoplancton (26.3%), mientras que en los intervalos de tallas de 2.1 a 4.0 cm se apreció un incremento en el consumo de fitoplancton (34.7%) y microcrustáceos (25.6%), disminuyendo a su vez el de protozoarios (19.3%). Los ejemplares de tallas entre 4.1-5.0 cm consumieron principalmente detritus (32.2%) y protozoarios (19.5%). Se observa una tendencia hacia la generalización trófica en los intervalos de tallas intermedios (Fig. 3A).
En la laguna de Chelem se analizaron 219 ejemplares correspondientes a los meses de julio y septiembre, con tallas de entre 1.1 a 9.0 cm de LE. Los grupos tróficos más abundantes durante esta época al igual que en la época de secas fueron el fitoplancton (44.0%), macrófitas (14.6%), microcrustáceos (14.3%), detritus (9.4%) y protozoarios (12.6%) (Tabla 2). Los componentes preferenciales fueron ocho y las presas secundarias 26, con FO de 10.5% a 49.3% (Tabla 4). El resto de los componentes alimenticios (26) se consideraron accidentales. Los organismos de tallas pequeñas (1.1 a 2.0 cm) se alimentan principalmente de fitoplancton (58%), pero también forman parte importante en su alimentación los microcrustáceos, detritus y protozoarios (12%, 11% y 11% respectivamente). Con el incremento de talla de 2.1 a 6.0 cm de LE se mantiene el fitoplancton como el alimento principal con el 45%, aumenta el consumo de microcrustáceos (14%), protozoarios (14%), macrófitas (12%) y detritus (10%). En los individuos de mayor talla (6.1 a 9.0 cm de LE) el grupo alimenticio dominante fueron las macrófitas (43%), disminuyendo su preferencia por el fitoplancton (22%) y microcrustáceos (11%) (Fig. 3B).
Nortes. En la laguna Puerto Viejo se analizaron 86 ejemplares correspondientes a los meses de noviembre y enero, con intervalo de tallas de 1.2 a 6.0 cm de LE. Fitoplancton (41.8%), microcrustáceos (14.2%), protozoarios (14.0%), detritus (11.6 %) y macrófitas (10.6%) fueron los grupos tróficos más abundantes (Tabla 2). Los componentes preferenciales fueron nueve y las presas secundarias 33, con FO de 10.5% a 48.8% (Tabla 5). El resto de los componentes alimenticios (34) se consideraron accidentales. Durante esta época del año el crecimiento de esta especie se caracterizó por presentar una gran generalización trófica con el mayor consumo de detritus (17.8%), protozoarios (28.6%) y fitoplancton (45%) en los individuos con tallas entre 1.1 y 3.0 cm de LE. En los intervalos de tallas de 3.1 a 5.0 cm de LE, el consumo de detritus disminuyó (8.9%), aumentando el de microcrustáceos (32.2%) y fitoplancton (49.1%), los cuales alcanzaron en estos intervalos sus máximos valores de consumo. Los individuos con intervalo de tallas de 5.1-6.0 cm presentaron una mayor generalización trófica incluyendo en su dieta mayores cantidades de los otros grupos tales como anélidos (11.5%), moluscos (2.9%) y porífera (3.9%) además de fitoplancton (25.4%) y microcrustáceos (15.9%) (Fig. 4A).
]]>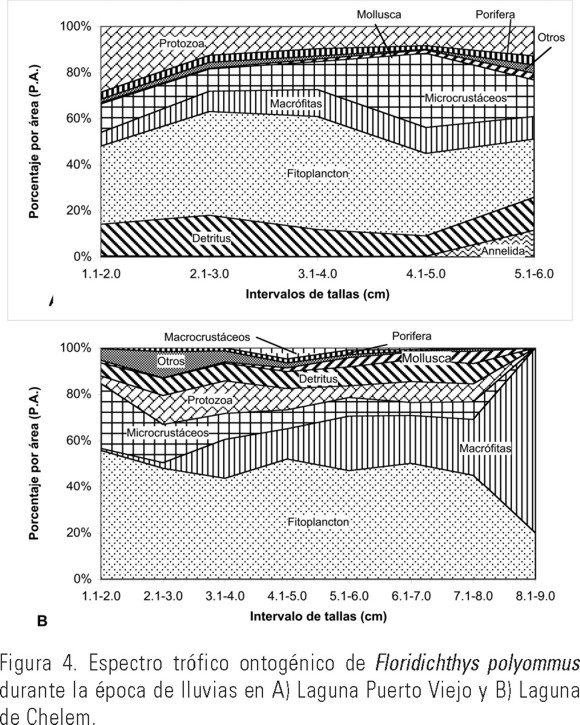
En la laguna de Chelem se analizaron 251 ejemplares correspondientes a noviembre y enero, con tallas de 1.5 a 9.0 cm de LE. Los grupos tróficos más abundantes durante esta época al igual que en la época de secas y lluvias fueron el fitoplancton (47.9%), macrófitas (16.4%), microcrustáceos (10.3%), protozoarios (9.8%) y detritus (7.8%) (Tabla 2). Los componentes preferenciales fueron nueve, las presas secundarias 21, con FO de 12.8% a 48.6% (Tabla 5). El resto de los componentes alimenticios (42) se consideraron accidentales. En esta época no se observó variación en cuanto a los componentes alimenticios con respecto a los meses de secas y lluvias. Los organismos de tallas pequeñas (1.1 a 2.0 cm) consumieron principalmente fitoplancton (56%) y microcrustáceos (28%). Con el incremento de talla de 2.1 a 4.0 cm, el fitoplancton (46%) y los microcrustáceos (14%) siguieron siendo los grupos preferenciales, incrementando el consumo de macrófitas (10%), protozoarios (13%) y detritus (8%). Conforme esta especie aumentó de talla (4.1 a 8.0 cm) se mantuvieron estables sus preferencias hacia el fitoplancton (47%), macrófitas (23%), detritus (8%) y protozoarios (7%). En ejemplares adultos (8.1 a 9.0 cm) el componente alimenticio dominante fueron las macrófitas, con el 80% del contenido estomacal (Fig. 4B).
Discusión
Se determinó que F. polyommus pertenece a la categoría de consumidores de microcrustáceos en la laguna Puerto Viejo y de fitoplancton en la laguna de Chelem. La categoría trófica referida en la primera laguna ha sido descrita por Vega-Cendejas et al. (2000). Por su parte Schmitter-Soto (1998) incluye a esta especie dentro de las consumidoras principalmente de detritus, que consumen secundariamente en su dieta anfípodos, copépodos, poliquetos y algas.
En relación a la ontogenia, se observó que la dieta cambió con el crecimiento. De acuerdo con Tresierra y Culquichicom (1995), la mayoría de los peces en sus edades tempranas, son omnívoros, pero conforme el pez va creciendo, sus preferencias alimenticias cambian y algunos se vuelven altamente selectivos. En el caso de F. polyommus, en la laguna Puerto Viejo se determinó una tendencia a la selectividad en los intervalos de tallas menores y mayores, ocurriendo el mayor aumento del espectro trófico en los intervalos de tallas intermedios. Los más pequeños presentaron una alimentación basada principalmente en microcrustáceos y los más grandes en el consumo de fitoplancton. En la laguna de Chelem se determinó una tendencia con el crecimiento hacia la especialización en el consumo de macrófitas. Los juveniles ingirieron mayores proporciones de fitoplancton y microcrustáceos, mientras que los ejemplares adultos consumieron principalmente macrófitas (ocupando hasta un 80% del peso total del contenido estomacal). Los resultados obtenidos en la laguna de Chelem coinciden con los reportados por Vega-Cendejas (1998), que considera a esta especie como consumidora de microcrustáceos en sus etapas juveniles y de macrófitas en sus etapas adultas.
En relación a las diferencias tróficas ontogénicas, se ha observado que reflejan las capacidades de los peces de ciertas tallas para capturar alguna presa (Sumton & Greenwood, 1990). Por tanto, a pesar de que el patrón de alimentación de los ejemplares juveniles basado en microcrustáceos (fuente rica en proteínas y con alto contenido energético) podría estar en función de los requerimientos energéticos de acuerdo al crecimiento, dicha preferencia puede requerir un mayor gasto de energía por ser los microcrustáceos considerablemente más móviles (Deudero, 2001). Además se ha observado que dicho grupo trófico aporta siete veces menos transferencia de biomasa a través de la predación en comparación con los grupos fitoplancton y macrófitas (Vega-Cendejas, 1998), por lo cual es posible que el mayor consumo de microcrustáceos en las etapas tempranas del desarrollo de esta especie se deba a su disponibilidad en el medio. Otro aspecto importante de estos cambios ontogénicos en la dieta es que permiten una repartición de los recursos con otras especies, generando de esta forma mayores tasas de crecimiento entre ellas y reduciendo la competencia interespecífica (García-Berthou & Moreno-Amich, 2000). Esto puede estar sucediendo, ya que resultados obtenidos por Vega-Cendejas et al. (2000) demuestran que de las 28 especies que conforman la comunidad en la laguna Puerto Viejo, siete se encuentran en la categoría de consumidores de microcrustáceos. En el caso de la laguna de Chelem se han registrado 53 especies de peces (Vargas, 2004), de las cuales dos también incluyen en su alimentación a los microcrustáceos, fitoplancton y macrófitas, entre otros grupos tróficos (Canto, 2004).
En cuanto a la variación trófica temporal registrada en Puerto Viejo, se obtuvo que el grupo trófico preferencial y más abundante (35.1%) durante la época de secas fue el de los microcrustáceos, mientras que en lluvias y nortes lo fue el fitoplancton. En la laguna de Chelem el grupo trófico preferencial y más abundante durante secas y lluvias siempre fue el fitoplancton con el mayor consumo en la primera época. En nortes el fitoplancton siguió siendo el principal grupo consumido; sin embargo, el consumo de macrófitas fue mayor en comparación con las otras dos épocas del año.
Las diferencias de consumo de los grupos tróficos a nivel estacional y entre los intervalos de tallas podrían explicar la coexistencia de los individuos de diferentes etapas ontogénicas en el área de estudio durante todo el año. Estas diferencias tróficas, al estar en sincronía con los patrones de productividad estacional, dan como resultado una repartición de los recursos (Vega-Cendejas, 1998). En el caso particular de la laguna de Chelem, el mayor consumo de fitoplancton durante la época de secas se debe a que en esta época del año los valores de productividad se incrementan por las altas temperaturas y elevada salinidad, lo cual da lugar a procesos de descomposición de materia orgánica y mineralización, proporcionando un aporte de nutrientes a la columna de agua que es aprovechado por el fitoplancton (Collí, 2001). Durante esta misma época, en la laguna Puerto Viejo, el mayor consumo de microcrustáceos en los estómagos se explica por el desarrollo de macrófitas, debido al incremento en las concentraciones de oxígeno por la alta incidencia lumínica que favorece el desarrollo de este grupo trófico (González, 1995), sobre el cual habitan gran cantidad de copépodos, anfípodos y otros microcrustáceos.
Una característica que forma parte del éxito de sobrevivencia de esta especie es que hace uso de otros recursos disponibles cuando los preferenciales son escasos debido al aumento de temperatura y salinidad, condiciones que pueden ser toleradas por muy pocos de sus competidores potenciales, como Atherinomorus stipes (Müller & Troschel) y Lucania parva (Baird & Girard) que también forman parte de las especies dominantes en la laguna Puerto Viejo, así como Opsanus phobetron (Walters & Robins) y Lagodon rhomboides (Linnaeus) en la laguna de Chelem. Como resultado, sus poblaciones llegan a ser abundantes, dominando en aguas con altas temperaturas y salinidades. Resultados obtenidos por Vega-Cendejas et al. (2000) demuestran que la época de secas se caracteriza por presentar los valores más bajos de densidad y biomasa de peces en la laguna Puerto Viejo. Sin embargo, F. polyommus tuvo los mayores valores de densidad promedio (62.1 g x 100 m−2) en esta época del año, cuando las salinidades y temperaturas son extremas (>40 y 30ºC). En la laguna de Chelem las temperaturas son de 35ºC, con salinidades hasta de 48 ppm; aunque no se tiene un registro de las especies dominantes en esta época, se sabe que en lluvias (31ºC y 40 ppm) se registran menor densidad y biomasa de peces en comparación con nortes (21ºC y 29 ppm) y la especie dominante es F. polyommus (Vargas, 2004). Gerking (1994) menciona que los requerimientos energéticos de los ciprinodóntidos son mayores en aguas calientes, por lo que forrajean intensamente por el recurso alimenticio principal, los pequeños invertebrados, los cuales, al disminuir su disponibilidad, son cambiados por detritus, algas verdes o diatomeas.
]]> En la época de lluvias, el aumento en la concentración de nutrientes, principalmente nitratos y sílice soluble reactivo, así como la disminución de la salinidad, favorece la presencia de bacilariofitas (Herrera-Silveira et al., 1999), lo cual explica los altos valores de FO de diatomeas en los estómagos durante esta época del año en ambas lagunas. En general, la mayor abundancia de fitoplancton consumido también puede ser una respuesta a la disminución del recurso energético principal (microcrustáceos) debido a una mayor demanda por parte de las especies que compiten por él, lo cual a su vez permite que otros recursos disponibles en el ecosistema sean aprovechados por las especies más aptas. Es decir, existe en realidad baja disponibilidad de la presa animal preferencial como resultado de una competencia interespecífica que induce un cambio de dieta (Declerck et al., 2002). Esto también explica la alta frecuencia en los estómagos de alimentos bajos en energía, como Porifera.La presencia de fitoplancton en los estómagos de F. polyommus durante la época de nortes se debe a que durante esta época del año los fuertes vientos ocasionan una mayor oxigenación, lo cual favorece la producción de este recurso (Herrera-Silveira et al., 1998). En el caso de la laguna de Chelem se han registrado 101 especies de fitoplancton, variando sus densidades de 2.7x107 cél/l en la época de nortes a 5.3x107 cél/l en la época de secas (Herrera-Silveira et al., 1999).
Cabe resaltar que la capacidad de adaptación de las especies de ciprinodóntidos de vivir en ambientes caracterizados por altas temperaturas y salinidades se debe en parte a su alta generalización trófica; si bien ésta es una característica presente en la mayoría de los peces, el éxito de sobrevivencia en dichas condiciones de estrés va a depender no solo de la ecofisiología de la especie, sino también de la amplitud del espectro alimentario de sus competidores en el área, de tal forma que, al disminuir el número de ciertos grupos tróficos, puedan reemplazarlos por otros existentes en el ecosistema (Tresierra & Culquichicom, 1995).
Las diferencias entre las dos lagunas en cuanto al consumo y preferencia temporal de los distintos grupos tróficos en la dieta de F. polyommus pueden estar en función de la disponibilidad de los recursos ocasionada por los cambios hidrológicos presentes en cada época climática; sin embargo, en general la dieta de esta especie está constituida principalmente por microcrustáceos, fitoplancton y macrófitas en ambos sistemas lagunares.
Agradecimientos
Deseamos agradecer a la bióloga Mirella Hernández de Santillana por la ayuda proporcionada en la revisión de las muestras de contenido estomacal. Esta investigación fue apoyada financieramente por CANABIO (Y027) y CONACyT-SISIERRA.
Referencias
BLABER, J.M. 1997. Fish and Fisheries of Tropical Estuaries. Chapman & Hall. Londres. 367 p. [ Links ]
CANTO, W.G. 2004. Análisis trófico de las especies ícticas dominantes en el estero de Yucalpetén. Tesis de Licenciatura, Instituto Tecnológico Agropecuario, Conkal, Yucatán. 85 p. [ Links ]
COLLÍ, R.C. 2001. Diagnóstico de la salud ambiental de la laguna de Chelem. Tesis de Maestría. Instituto Tecnológico de Mérida. 160 p. [ Links ]
DECLERCK, S., G. LOUETTE, T. DE BIE & L. DE MEESTER. 2002. Patterns of diet overlap between populations of non-indigenous and native fishes in shallow ponds. Journal of Fish Biology 61: 1182-1197. [ Links ]
DEUDERO, S. 2001. Interspecific trophic relationship among pelagic fish species underneath FADs. Journal of Fish Biology 58: 53-67. [ Links ]
GARCÍA-BERTHOU, E. & R. MORENO-AMICH. 2000. Food of introduced pumpkinseed sunfish: ontogenetic diet shift and seasonal variation. Journal of Fish Biology 57: 29-40. [ Links ]
GERKING, S. D. 1994. Feeding Ecology of Fish. Academic Press. Nueva York. 416 p. [ Links ]
GONZÁLEZ A., A. F. 1995. La Comunidad de Peces Asociada al Manglar de la laguna Costera de Celestún, Yucatán, México. Tesis de Licenciatura, Universidad Nacional Autónoma de México, México. 83 p. [ Links ]
HERRERA-SILVEIRA, J.A., M. MARTÍN & V. DÍAZ-ARCE. 1999. Variaciones del fitoplancton en cuatro lagunas costeras del estado de Yucatán, México. Revista de Biología Tropical 47 (Supl. 1): 47-56. [ Links ]
HERRERA-SILVEIRA, J.A., J. RAMÍREZ & A. ZALDIVAR. 1998. Overview and characterization of the hydrology and primary producer communities of selected coastal lagoons of Yucatán, México. Aquatic Ecosystem Health and Management 1: 353-372. [ Links ]
HYSLOP, E.J. 1980. Stomach contents analysis: a review of methods and their application. Journal of Fish Biology 17: 411-429. [ Links ]
PREJS, A. & G. COLOMINE. 1981. Métodos para el Estudio de los Alimentos y las Relaciones Tróficas de los Peces. Universidad Central de Venezuela. Caracas. 127 p. [ Links ]
SCHMITTER-SOTO, J.J. 1998. Catálogo de los Peces Continentales de Quintana Roo. El Colegio de la Frontera Sur. San Cristóbal de Las Casas. 239 p. [ Links ]
SUMTON, W. & J. GREENWOOD. 1990. Pre-and post-flood feeding ecology of four species of juvenile fish from the Logan-Albert estuarine system, Moreton Bay, Queensland. Australian Journal of Marine and Freshwater Research 41: 795-806. [ Links ]
TRESIERRA, A.E. & Z.G. CULQUICHICOM 1995. Manual de Biología Pesquera. Trujillo. México. 227 p. [ Links ]
VARGAS, K.E. 2004. Composición y distribución íctica durante la temporada de lluvias y nortes en el estero de Yucalpetén, Yucatán, México. Tesis de Licenciatura, Universidad Autónoma de Yucatán. Mérida. 57 p. [ Links ]
VEGA-CENDEJAS, M.E. 1990. Interacción trófica entre los bagres Arius melanopus (Agassiz, 1829) y Arius felis (Linnaeus, 1766) en las costas de Celestún, Yucatán, México. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 17 (2): 271-285. [ Links ]
VEGA-CENDEJAS, M.E., M. HERNÁNDEZ & F. ARREGUÍN-SÁNCHEZ. 1994. Trophic interrelations in a beach seine fishery from the northwestern coast of the Yucatan peninsula, Mexico. Journal of Fish Biology 44: 647-659. [ Links ]
VEGA-CENDEJAS, M.E. 1998. Trama trófica de la comunidad nectónica asociada al ecosistema de manglar en el litoral Norte de Yucatán. Tesis de Doctorado, Facultad de Ciencias, Universidad Nacional Autónoma de México, México. 170 p. [ Links ]
VEGA-CENDEJAS, M.E., M. HERNÁNDEZ, G. MEXICANO, G. AVILÉS, E. FALFÁN & V. GARCÍA. 2000. Informe técnico final del proyecto de investigación "Ecología y Evaluación de las Poblaciones de Peces en los Sistemas Costeros del Parque Marino Isla Contoy". Recursos del Mar, Centro de Investigación y Estudios Avanzados, Instituto Politécnico Nacional. Mérida. 100 p. [ Links ]
VEGA-CENDEJAS, M.E. & M. HERNÁNDEZ DE S. 2004. Fish community structure and dynamics in a coastal hypersaline lagoon: Río Lagartos, Yucatán, México. Estuarine, Coastal and Shelf Science 60: 285-299. [ Links ]
WOOTTON, R.J. 1990. Ecology of Teleost Fishes. Chapman and Hall. Londres. 404 p. [ Links ]
YÁÑEZ-ARANCIBIA, A. 1975. Estudios de peces en las lagunas costeras: nota científica. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 2 (1): 53-60. [ Links ]
]]>