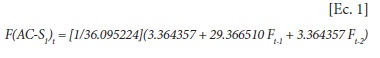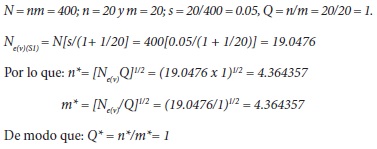
Endogamia en el maíz en la selección recurrente para aptitud combinatoria
Inbreeding in maize submitted to recurrent selection for combining ability
Fidel Márquez-Sánchez†
Centro Regional Universitario Occidente, Universidad Autónoma Chapingo. Rosario Castellanos 2332, Col. Residencial la Cruz. 44950, Guadalajara, Jalisco, México.
]]>Recibido: 20 de Diciembre del 2013
Aceptado: 13 de Julio del 2014.
Resumen
En los tres tipos de selección recurrente para aptitud combinatoria en el maíz (Zea mays L.) se usan como unidades de selección a líneas S1, mientras que los criterios de selección son los mestizos hechos por las cruzas entre las unidades de selección y los probadores: una variedad de polinización libre, una línea altamente endogámica y la población B para las líneas S1 derivadas de A, y la población A para las líneas S1 derivadas de B. La endogamia que se genera ocurre por los cruzamientos entre las líneas seleccionadas S1 en los tres tipos de selección recurrente, aunque las líneas S1 no serán las mismas en los tres tipos de selección. En los cálculos que aquí se presentan se usaron 20 líneas S1 y 20 plantas por línea, que produjeron una alta endogamia (0.59) en el ciclo 10 de selección. Para reducir la endogamia a 0.20 basta con hacer solamente dos o tres ciclos de selección.
Palabras clave: Zea mays, endogamia, selección recurrente, aptitud combinatoria.
Abstract
In the three types of recurrent selection for combining ability in maize (Zea mays L.), S1 lines are used as selection units. The selection criteria are the top crosses made among S1 lines and their respective testers: an open-pollinated variety; a highly homogeneous line; and variety B for the S1 lines from variety A, and variety A for S1 lines from variety B. Generated inbreeding occurs because of crossing among selected S1 lines in the three types of recurrent selection, though selected S1 lines are not the same in the three types of selection. In this paper, considering 20 lines S1 y 20 plants per line, inbreeding in cycle ten reached a very high value (0.59). Reduction of inbreeding to 0.20 can be achieved by making only two or three selection cycles.
]]> Key words: Zea mays, inbreeding, recurrent selection, combining ability.
INTRODUCCIÓN
En los ciclos de selección recurrente aplicada al maíz (Zea mays L.) se presenta endogamia debido a que el número de familias seleccionadas es relativamente pequeño, aún sin considerar el ajuste por el número efectivo de varianza que participa en el cálculo del coeficiente de endogamia. Para reducir el efecto nocivo de la endogamia, o bien se incrementa el número de familias o se incrementa la presión de selección, pero la segunda opción causará una reducción del número de familias por probar.
En el cálculo de la endogamia que ocurre en la selección familiar aplicada al maíz, Márquez-Sánchez (2009) usó 180 familias con selección de las mejores 10 en cada ciclo. Posteriormente, Márquez-Sánchez (2013) propuso aumentar a 20 el número de familias (n = 20) con 20 plantas por familia (m = 20), para un total de 400 plantas.
Para la selección recurrente de líneas S1, de familias de hermanos completos y de familias de medios hermanos, Márquez-Sánchez (2009) calculó el número efectivo de varianza de acuerdo con Crossa y Venkovsky (1997), y para estos tres tipos de selección obtuvo que los números de familias por probar son iguales a 9, 6 y 7, respectivamente. Aquí es de destacar la notable contradicción de que el número de líneas en las familias de hermanos completos resulta menor que el número en las familias de medios hermanos.
La finalidad del presente escrito es calcular la endogamia al incrementar el número de familias bajo prueba en la selección recurrente para aptitud combinatoria (AC), con la presión de selección acostumbrada y con 400 plantas.
MATERIALES Y MÉTODOS
Los cálculos en este apartado pueden aplicarse a los tipos de selección recurrente para: aptitud combinatoria general (ACG), aptitud combinatoria específica (ACE) y la selección recíproca recurrente (SRR) para dos variedades (A y B), pues aunque el cálculo de la endogamia es el mismo para los tres tipos de familias, las líneas seleccionadas son diferentes debido a los tipos de probadores que se usan en los tres tipos de selección recurrente: variedad de polinización libre para ACG; línea pura para ACE; y en la SRR la variedad A para la variedad B, y la variedad B para la variedad A.
]]> El cálculo de la endogamia se hizo de acuerdo con el número de líneas S1 seleccionadas, antes de la obtención de nuevas líneas S1 para el siguiente ciclo de selección. El cálculo depende del número efectivo de varianza [Ne(v)] el cual se obtendrá de acuerdo con la intensidad de selección (s), del número total de líneas (n) y del número de plantas por línea (m), según Crossa y Venkovsky (1997). Con Ne(v) se calculan los números ajustados n* y m*, con Q = n/m.Entonces:
La recombinación entre las líneas seleccionadas se hizo conforme al diseño dialélico en un solo sentido. En el Cuadro 1 se muestra el dialélico hecho con n líneas; los cruzamientos se encuentran en la columna; en la diagonal están los cruzamientos dialélicos en número m(m-1)/2; arriba de la diagonal, en cada celda hay m2 cruzamientos entre plantas.

Se debe considerar también que en total de n líneas, los cruzamientos entre ellas son: nm(m-1)/2 y nm2(n-1)/2; a su vez, estos números se codifican al dividir a cada uno por nm/2, de manera que quedan como: m-1 y m(n-1). Con el objeto de calcular la endogamia en cada tipo de cruzamientos (dentro de las líneas y entre las líneas), es necesario tomar en cuenta a las coancestrías en el ciclo 0 de selección, las que en el ciclo 1 de selección darán lugar a los respectivos valores de la endogamia.
En los cruzamientos dentro de las líneas S1 la endogamia es la misma que la de las líneas S1 (Márquez-Sánchez, 2007), y los cruzamientos entre las líneas producirán híbridos entre líneas S1 con endogamia igual a F1. En el Cuadro 2 se muestra el total de esta información. Finalmente, la endogamia de la recombinación entre las líneas S1 se calculó como el promedio de la suma de los números codificados por las coancestrías, y se divide entre en total de los números codificados (nm-1).
]]> RESULTADOS Y DISCUSIÓN
Como la población de donde se derivaron las líneas S1 corresponde al ciclo 0, entonces el ciclo 1 corresponde al de las líneas S1, y la endogamia del ciclo 2 sería:
Por extrapolación, la ecuación para cualquier ciclo de selección t es:
En la Figura 1 se muestra la curva de endogamia de la Ec. 1 para 100 ciclos de selección. Puede verse que en el ciclo 10 (un número muy usado de ciclos de selección) la endogamia es igual 0.60, valor que es muy alto. Para alcanzar un valor endogámico de alrededor de 0.20, sólo serían necesarios dos o tres ciclos de selección.
CONCLUSIONES
]]> La selección recurrente para aptitud combinatoria general es un método que genera valores muy altos de la endogamia en los primeros ciclos de selección.
BIBLIOGRAFÍA
Crossa J. and R. Venkovsky (1997) Variance effective population size for two-stage sampling of monoecious species. Crop Science 37:14-26. [ Links ]
Márquez-Sánchez F. (2009) Expected inbreeding in recurrent selection in maize. III: Selection in S1lines and full-sib and half-sib families. Maydica 54:109-111. [ Links ]
Márquez-Sánchez F. (2013) Cálculo de la endogamia en la selección recurrente de familias en el maíz. Revista Mexicana de Ciencias Agrícolas 4:153-158. [ Links ]
]]>