











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La ganadería bovina productora de carne en México ocupa el lugar diez en el mundo, con una tasa media de crecimiento anual en el periodo 1990-1997 de 2.6%.1 La producción nacional fue de 1379 941 ton en 1998.2 Como consecuencia de la importancia de la producción de carne bovina en el país y en el mundo, resulta prudente optimizar dicha producción; entendiéndose con esto último, que se envíen al mercado canales de buena calidad de animales sanos, pero que sean del agrado del consumidor final.
En México la mayor parte de los rumiantes son engordados y finalizados bajo condiciones de pastoreo.2 Se ha observado que bajo estas condiciones, el tejido adiposo de los bovinos para abasto puede presentar coloración casi amarilla debido a los carotenos y carotenoides provenientes de su ingesta. A pesar de que la coloración de la grasa no afecta el sabor ni el valor alimentario de las porciones comestibles, la presencia de los pigmentos representa un problema económico serio como resultado del rechazo del producto por parte del consumidor, tanto nacional como extranjero.
El ganadero ha tratado de superar el problema mediante el confinamiento de sus animales durante algunos meses previos al sacrificio, lo que no sólo no garantiza la efectividad de la despigmentación sino que resulta en un incremento innecesario en los costos de producción, así como en la economía del consumidor final.
Hasta el momento se han dilucidado algunos de los procesos metabólicos que explican el depósito de carotenoides en el tejido adiposo de los bovinos que se detallan en este trabajo; sin embargo, resulta indispensable ampliar esta visión del problema y proponer soluciones viables y económicas al ganadero, de forma que se puedan obtener canales “blancas” de animales finalizados en pastoreo.
Antecedentes
Magnitud económica del problema de grasa amarilla
La producción de carne de bovino ha evolucionado tecnológicamente a un ritmo menor que la avicultura y la porcicultura; sin embargo, el valor de su producción representa 39.3% del total de la carne contra 29.5% y 28.6% de la de cerdo y pollo, respectivamente.3
La producción de ganado bovino se desarrolla bajo diferentes contextos agroclimáticos, tecnológicos, de sistemas de manejo y de explotación. Los sistemas básicos de explotación de bovinos de carne son el intensivo (corral de engorda) y el extensivo (engorda en praderas y agostaderos).3 El Centro de Estadística Agropecuaria (CEA) señala que para 1998 se obtuvo una producción de 1 379 941 ton de carne de bovino en canal y cortes. Si el peso promedio al sacrificio en México es de 450 kg, en promedio, significa que durante ese año se sacrificaron 5,575,519 animales. Asimismo, si el total de bovinos sacrificados provenientes de corral fue de 1,200,000, entonces la carne de bovino proveniente de corral representa 21.5% y la de pastoreo 78.5% del total producido.2-5
En una consulta realizada en 1998 en los archivos de sacrificio del rastro TIF de la Asociación Ganadera de Tabasco, se observó que el color casi amarillo de la grasa afectó a 5% de las canales (Barrón et al., comunicación personal). Si esto sucediera en la población total de animales finalizados en pastoreo, significaría que aproximadamente 54 mil toneladas de carne provendrían de canales con grasa amarilla. Al considerar un consumo per capita de 16.3 kg de carne de bovino, la depreciación del 10% del tonelaje mencionado equivaldría al consumo anual de carne bovina de cerca de 300 mil mexicanos o a una pérdida de 100 millones de pesos.2-4
Carotenoides
Según la Unión Internacional de Química Pura y Aplicada,5 el término carotenoide se aplica a un gran número de compuestos liposolubles que colorean de amarillo a rojo. Esta característica cromófora se debe a la presencia de una serie de dobles enlaces conjugados. Esta gran familia de pigmentos se ha agrupado como hidrocarbonados (licopeno, a, b, g carotenos) y los derivados oxigenados (oxicarotenoides o xantofilas)6,7.
La estructura básica de los carotenoides consta de cinco a ocho unidades isoprenoides, unidas de manera normal de cabeza a cola, excepto en el centro de la molécula, donde el orden es inverso al esqueleto de C40. Los dos grupos metilo centrales están en las posiciones 1,6; los grupos metilo restantes no terminales están en posiciones 1,5.7
Los carotenoides están ampliamente distribuidos en la naturaleza y son sintetizados únicamente por medio de las plantas y las bacterias fotosintéticas. Los animales dependen completamente del alimento como fuente de carotenoides y no tienen vías metabólicas para la síntesis de novo de estos compuestos.7,8
Está suficientemente documentado que los carotenoides son constituyentes normales de la sangre y tejidos de los humanos, bovinos, equinos, hurones, gerbos, peces, aves y algunos crustáceos.8-11
En los bovinos el color amarillo de la grasa se debe a la presencia de este tipo de pigmentos, de los cuales domina el β-caroteno, aunque también han sido detectadas pequeñas cantidades de luteína.12-14
Se ha observado que al incrementarse el contenido de carotenoides en el tejido adiposo subcutáneo de bovinos, hay un incremento significativo del porcentaje total de ácidos grasos cis-monoinsaturados y un decremento de los ácidos grasos saturados;15 aunque esto último no implica que los carotenoides sean los responsables de cambiar la relación de los ácidos grasos en el tejido adiposo.
En los animales para abasto la apariencia es muy importante; por ejemplo, en el pollo de engorda, es deseable la pigmentación amarillo-anaranjada de la piel y de la grasa. Con ese propósito se suplementan en la dieta diversos productos que contienen oxicarotenoides;8,16 sin embargo, en los bovinos lo deseable es el color blanco de la grasa, pero en estos animales los carotenoides se depositan de manera natural cuando consumen forrajes verdes.13,14,17
Conversión de los carotenoides a vitamina A
En los animales los carotenoides pueden convertirse a vitamina A (Figura 2). La conversión de β-caroteno a retinol es un proceso importante, ya que la mayoría de la vitamina A que utilizan los animales deriva de este proceso. Después de su absorción, el carotenoide es separado en las células de la mucosa intestinal para formar vitamina A.7
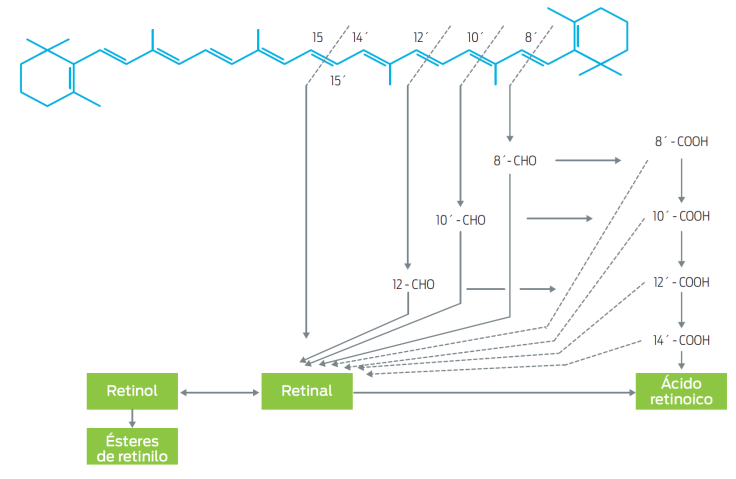
La conversión de β-caroteno en vitamina A fue demostrada en 1960,18 en ratas deficientes en vitamina A alimentadas con β-caroteno.
Glover19 propuso dos vías principales mediante las cuales estos compuestos pueden ser convertidos en vitamina A; ambas son reacciones oxidativas: 1) ruptura central, la cual ocurre en el doble enlace central 15,15´. El producto de este tipo de división son dos moléculas de retinal. La enzima responsable de convertir una molécula de β-caroteno en dos moléculas de retinal es la 15,15’dioxigenasa (E.C. 1.13.11.21); 2) ruptura excéntrica, que puede ocurrir en uno o más de otros dobles enlaces. Sus productos pueden ser un β-apo-carotenal largo y otro corto; por ejemplo, ciclocitral y β-apo-8’-carotenal, si acaso la ruptura ocurriera en el doble enlace 8’. Experimentalmente está demostrado que los b-apocarotenales pueden ser convertidos a β-caroteno.20
La separación enzimática de β-caroteno en retinaldehído fue registrada por dos grupos de investigadores en 1965.21,22 Goodman et al.23 observaron que la enzima 15,15´ dioxigenasa es estimulada por la adición de tioles y es fuertemente inhibida por sulfhidrilos y agentes quelantes. Subsecuentes estudios han mostrado que el oxígeno molecular reacciona con los dos carbonos centrales del β-caroteno, seguido por la separación del doble enlace central del caroteno para formar dos moléculas de retinaldehído.20,24
Formas parcialmente purificadas de esta enzima se han obtenido en intestinos de ratas, cerdos, conejos, hurones, gerbos, bovinos, cabras y humanos.20,21,23,25-29 La enzima también ha sido parcialmente purificada de hígado.20,24,28 Sin embargo, debido a su labilidad durante los procesos de purificación, la enzima pura no ha sido aislada todavía, razón por la cual los estudios sobre la actividad de dicha enzima utilizan fracciones proteínicas solubles de mucosa intestinal (obtenidas mediante centrifugaciones).
Asimismo, se ha observado que existe una inhibición de la actividad de la 15,15’ dioxigenasa por otros carotenoides, tales como licopeno, luteína, astaxantina y cantaxantina.30-34
La eficiencia de conversión de β-caroteno a vitamina A en la mucosa intestinal varía con la especie.7 A diferencia de los mamíferos, algunos peces y aves pueden formar vitamina A a partir de astaxantina, cantaxantina e isozeaxantina.24 Los peces pueden convertir la luteína en 3-hidro-retinol. 20 Es importante destacar que una porción de los carotenos absorbidos no es metabolizada en la mucosa intestinal, sino que es incorporada a los quilomicrones y pasa a la sangre vía linfática como tal. Estos carotenoides pueden ser depositados en el hígado o en otros lugares, tales como tejido adiposo u otros órganos.24
En los estudios con especies rumiantes se observó una actividad intestinal de la 15,15’ dioxigenasa cinco veces menor en bovinos, comparada con la observada en caprinos,28 esto último sugiere que en gran medida la escasez de esta enzima es responsable de que en los bovinos se absorban grandes cantidades de β-caroteno y se depositen en el tejido adiposo, lo que le da la coloración un tanto amarilla. La conversión de β-caroteno a retinal en el intestino de la rata se lleva a cabo a un pH óptimo de 7.5 a 8, con una Vmáx de 8 nmol de retinal formado por hora y una Km de 3 × 10 M. En conejos la Vmáx es aproximadamente de 1.4 nmol de retinal formado a partir de β-caroteno /mg de proteína × hora−1, a una temperatura de 37 ºC. 20
Mediante los valores de esta enzima, se puede considerar que la enzima posiblemente no se satura de manera total; en consecuencia, la tasa fisiológica bajo condiciones normales es sólo una fracción de la tasa máxima. De igual manera, los cálculos se refieren sólo a la actividad intestinal de la enzima, el hígado también la contiene, por lo que la actividad máxima en el organismo es mayor a lo que hasta ahora se ha calculado. Asimismo, se deben considerar las observaciones de que la tasa máxima es menor que la cantidad requerida para producir toxicidad crónica, por lo que aun cuando se den altas dosis de β-caroteno, no se produce hipervitaminosis A.20
La composición de los ácidos grasos de la dieta puede afectar la actividad de la 15,15’dioxigenasa, así como de la proteína ligadora de retinol celular.35,36 En este sentido, se ha observado que una dieta rica en ácidos grasos polinsaturados puede aumentar hasta 2.7 veces la actividad de dicha enzima, comparada con la mostrada en animales que consumen una dieta testigo.36
Desaparición ruminal de los carotenoides
Existen relativamente pocas investigaciones acerca del metabolismo ruminal de β-caroteno y la vitamina A. Shorland et al.37 realizaron estudios in vitro con líquido ruminal de ovinos. Se concluyó que los carotenos no son afectados por la fermentación ruminal. Sin embargo, existen informes de pérdidas retículo-ruminales de vitamina A y de carotenos. King et al.38 observaron destrucción de ambos cuando se incubaron in vitro en líquido ruminal. La recuperación de β-caroteno en muestras de líquido ruminal que fueron adicionadas de vitamina A y caroteno, y que no fueron incubadas, fue de 97.5%; sin embargo, en muestras que fueron incubadas durante nueve horas, el porcentaje de recuperación fue de 65.6. Los antioxidantes (santoquina y tocoferol) pueden reducir las pérdidas en periodos de incubación de diez horas.
Keating et al.39 utilizaron etoxiquina y nitrato de potasio para determinar la destrucción in vitro de acetato de vitamina A y β-caroteno. En la determinación de vitamina A no se encontraron diferencias entre tratamientos (basal + nitrato + etoxiquina + nitrato + etoxiquina) después de cuatro horas de incubación; sin embargo, después de 16 horas de incubación los porcentajes de recuperación fueron de 39 para el basal, 87.6 para el que contenía nitrato, 87.6 para el que estaba adicionado con etoxiquina y 75 para el que contenía ambos. Para el caso de β-caroteno no se encontraron diferencias entre tratamientos ni a las 3.5 (83.4%) ni a las 16 horas de incubación (72.7%), aunque no se indica si hubo utilización.
Klatte40 observó que los microorganismos ruminales destruyen a la vitamina A, por lo que un cambio en la población microbiana puede alterar la destrucción de vitamina A.
Van Soest41 señala que si bien los carotenoides pueden ser atacados por las bacterias anaeróbicas e hidrogenarse, el sistema de dobles enlaces conjugados de estos compuestos (posición trans) los hace más resistentes a la hidrogenación, comparados con compuestos insaturados no conjugados, lo que permite que éstos lleguen prácticamente intactos al intestino delgado.
Informes recientes señalan que utilizando el método in vitro, existe una menor desaparición de β-caroteno en novillos (rumiantes que pigmentan la grasa) y caprinos (rumiantes que no pigmentan la grasa), que la informada anteriormente en la literatura. Mora et al.42 señalaron tasas de desaparición (kd) de β-caroteno de 0.13% y 0.37% × h−1, y para carotenos totales de 0.20 y 0.62% × h−1 para novillos y cabras, respectivamente. Estos resultados contrastan con los notificados por otros investigadores,38,39 donde las desapariciones son mucho mayores ya que obtuvieron kd de 3.03% × h−1 y 2.25% × h−1 para β-caroteno.
Las degradaciones tan altas indicadas por King et al.38 y Keating et al.39 los hicieron pensar que el β-caroteno estaba siendo destruido a nivel ruminal, lo que se atribuyó a los microorganismos ruminales.43 Van Soest,41 por su parte, señala que los carotenoides tienen una digestibilidad de 0.1% a nivel ruminal (en forma de una hidrogenación destructiva), pero que estos compuestos son muy poco utilizados en ese ámbito. Con esta información 41,42 quizá se puedan atribuir las altas kd más a la degradación que sufren los carotenoides debido a la luz o a la temperatura durante el análisis, que a la acción de la microflora ruminal tal como lo informan Schmitz et al.44
Mora et al.42 registran tasas de degradación in situ de β-caroteno y carotenos totales de 2.5% y 1.2 % × h−1 para novillos, respectivamente, y de 2.2% y 1.0 % × h−1 para cabras, respectivamente. King et al.38 informaron pérdidas in vivo de 40% de β-caroteno en rumen a las 12 horas, mientras que Mora et al.42 encontraron pérdidas de 10.52% y 10.56 % entre las ocho y dieciséis horas, muy por debajo de lo registrado anteriormente por King et al.38
Es posible que la desaparición in situ de este compuesto esté asociada a la desaparición del contenido celular o de la materia seca; es decir, desaparecen por la sola desaparición de la materia seca (digestibilidad y paso al intestino delgado), y no por degradación per se. La comparación de las kd de β-caroteno y materia seca realizada por Mora et al.42 reveló resultados muy similares (89%, p < 0.01), lo que apoya esta suposición.
Aparentemente el rumen no es la causa de que existan especies de rumiantes que pigmenten o no la grasa, pues la degradación de carotenoides a nivel ruminal es baja tanto en bovinos como en caprinos, de tal forma que grandes cantidades de estos compuestos pasan intactos al intestino delgado.
Absorción y utilización de los carotenoides
Después de la ingestión del alimento los carotenoides son liberados por acción enzimática en el intestino delgado, y junto con la vitamina A son disueltos en los glóbulos grasos que pasan a través del lumen del duodeno. En este sitio, los glóbulos se encuentran con las sales biliares y las enzimas pancreáticas, las cuales liberan productos de la digestión lipídica. Los ésteres de vitamina A son hidrolizados en ese momento. Estos productos de la digestión interactúan con las sales biliares y el colesterol y forman micelas mixtas, en las cuales se solubilizan la vitamina A y los carotenos. Estas micelas se difunden mediante la glicoproteína de la membrana de las células de la mucosa. Los componentes de la micela, excepto las sales biliares, penetran individualmente a la fase lipídica de la membrana de las células de la mucosa.7
La presencia o ausencia de otros componentes de la dieta, así como el estado nutricional del individuo, afectan la eficiencia de dispersión. En primer lugar, las proteínas solubles y los péptidos derivados de ellas ayudan a la dispersión de la vitamina. El incremento en el nivel de proteína en la dieta ayuda a la conversión in tracelular del caroteno a retinaldehído.7 Grownowska-Senger y Wolf45 informan que la actividad de la enzima intestinal 15,15’-dioxigenasa se deprime en aproximadamente 50% con consumos de proteína muy bajos (5%). En segundo término, la grasa de la dieta provee el medio de transporte de la vitamina A y de los carotenoides del estómago al lumen intestinal. Además, algunos lípidos de la dieta como los aceites de semillas, que contienen α-tocoferol, tienen un efecto protector antioxidante sobre la vitamina A. En tercer lugar, para que se realice una eficiente absorción de la vitamina A y de los carotenoides, es necesaria la presencia de las sales biliares.7
Los niños muestran una capacidad ineficiente de absorción de carotenos. La tasa de absorción se encuentra disminuida debido a que la solubilidad de β-caroteno y de otros carotenoides en la grasa es limitada.46 Sin embargo, en becerros Holstein recién nacidos, al administrar una dosis oral de 20 mg de β-caroteno se observó un aumento en el suero entre las 12 y 30 horas posdosificación, que declinó después lentamente.47
Los carotenos presentes en los alimentos son menos absorbidos en el intestino que la vitamina A preformada, pero esto puede variar según la especie animal de que se trate.48 En bovinos se ha observado que los carotenoides suplementados en el alimento poseen una alta digestibilidad (hasta del 90%), y cuando son consumidos de manera natural en los forrajes, su digestibilidad es de aproximadamente 60%.28,49
Bajo condiciones normales en humanos, cerca de 90% de la vitamina A ingerida es absorbida, y la eficiencia de absorción disminuye muy poco al incrementar la dosis. Por otra parte, cerca de 70% de los β-carotenos ingeridos son absorbidos, y la eficiencia de absorción disminuye fuertemente al incrementar la dosis.7
Después de la absorción, los β-carotenos alojados en las células de la mucosa intestinal se convierten en retinaldehído, el cual es reducido a retinol. Durante su paso a través del epitelio intestinal, cerca de 75% del retinol es esterificado con ácidos grasos de cadena larga. Estos ésteres, comúnmente en forma de palmitato o ácido esteárico, son incorporados junto con otros lípidos y junto con las apoproteínas, a los quilomicrones, los cuales pasan a la linfa.7,24
Distribución en sangre y tejidos
Como ya se mencionó, los carotenoides son constituyentes normales de la sangre y tejidos de algunas especies,9-11 lo cual probablemente se debe a que no todos estos compuestos se convierten eficientemente en vitamina A, y parte de ellos son absorbidos y depositados como carotenoides.
No existe evidencia de que exista absorción y circulación de β-caroteno en ratas, cerdos, pollos, cuyes y conejos,9 debido a una conversión más eficiente de β-caroteno a vitamina A.
La cantidad total y las concentraciones específicas de cada carotenoide en la sangre y en los tejidos estarán en función del consumo diario de estos pigmentos.24 En bovinos se ha observado49 un aumento lineal (r2 = 0.97, p < 0.01) en la concentración plasmática de β-caroteno después de 30 días de suplementación con dicho compuesto.
Los carotenoides son transportados en la sangre en asociación con lipoproteínas, principalmente con lipoproteínas de baja densidad (LBD) en humanos y gerbos.7,11,24 Sin embargo, en bovinos y hurones están asociados con lipoproteínas de alta densidad (LAD).11
En humanos bien alimentados, los carotenoides están presentes principalmente en el tejido adiposo (80%-85%), en el hígado (8%-12%) y en el músculo (2%-3%). En otros tejidos se pueden observar bajas concentraciones, principalmente en el cuerpo lúteo y las glán dulas adrenales. De la reserva total de carotenoides (100-150 mg), el suero contiene normalmente 1%.24 El suero humano contiene como mayores componentes β-caroteno, γ-caroteno, criptoxantina, licopeno y luteína, y puede presentar bajas concentraciones de zeaxantina y otras xantofilas y polienos, tales como fitoflueno y fitoeno. 50
En bovinos neonatos se ha observado que la concentración plasmática de β-caroteno se incrementa en relación lineal con la dosis administrada, de dos hasta dieciséis µmol de β-caroteno /L, hasta alcanzar una meseta a las cuatro semanas de la suplementación con β-caroteno.51 En adultos existen informes que señalan que para animales que estuvieron cuatro meses en pastoreo, se pueden observar en suero concentraciones de 2.19 µg / mL, 52 la suplementación durante 30 días con β-caroteno, en dosis equivalentes a que los animales estuvieran en pastoreo, 49 produjo concentraciones en suero de 3.6 µg/mL. Knight et al.17 encontraron que la intensidad del color de la grasa está altamente correlacionada (r= 0.92) con el contenido de carotenoides y que la concentración de β-caroteno en el plasma está correlacionada con el color de la grasa (r = 0.67). Estos investigadores informan que la concentración de carotenos en plasma de bovinos que están en libre pastoreo es de 11.24 ± 0.12 µg β-caroteno/mL.
En cuanto al β-caroteno depositado en el tejido adiposo, la literatura registra de 0.81 a 3.7 µg/g 49,51 en el tejido adiposo subcutáneo y 0.23 µg/g en el perirrenal.49
Está demostrado que en el ganado bovino, el β caroteno depositado en el tejido adiposo representa entre 85% y 90% del color. 13,14,17 En este sentido, se ha observado que pueden existir factores genéticos asociados con el color de la grasa en bovinos. Barton y Pleasants12 calificaron el color de la grasa subcutánea de diferentes razas durante cinco años consecutivos, con los siguientes resultados: dentro de las razas de carne, más del 60% de las canales se clasifican como blancas; Angus es la que tiene una coloración de la grasa más amarilla, ya que posee alrededor de 40% de canales con este problema. De las razas especializadas en producción de leche, Jersey y Holstein son las que presentaron mayor cantidad de canales amarillas.
En cuanto a las técnicas para medir el color, la mayoría de los informes señalan el uso de la colorimetría como una herramienta adecuada, debido a que existe una buena correlación entre ésta y la cromatografía de líquidos, r = 0.6.15 También existen métodos subjetivos que implican el uso de cartas fotográficas o abanicos de colores que van de blanco a muy amarillo y se usan en el área del lomo o de la región costal.13
Almacenamiento en el hígado
Después de abandonar la célula intestinal, los ésteres de retinilo acarreados en las proteínas de baja densidad del plasma son probablemente hidrolizados por esterasas de las membranas de las células hepáticas.7 Intracelularmente el retinol no esterificado está ligado a una proteína específica y es transportado al retículo endoplásmico, una vez que es esterificado a ésteres de palmitato. Éste es transferido y almacenado en un complejo soluble o macromolécula, formada por varios tipos de lípidos, varias cadenas de polipéptidos y carbohidratos unidos covalentemente a proteínas.6
La vitamina A tiene una marcada preferencia para almacenarse en el hígado y en ese sitio es muy estable. La vitamina A hepática (95% en forma de palmitato de retinilo) representa cerca de 90% del total de las reservas corporales de esta vitamina. En cualquier individuo, la magnitud del almacenamiento depende no sólo del consumo de vitamina A y de sus provitaminas, sino también de la eficiencia de absorción y de la tasa de gasto. Estos factores están influidos por el sexo, la tasa de crecimiento, el estado de salud, etcétera.6,7
El hígado almacena vitamina A continuamente y la libera a la sangre cuando los niveles en la dieta son deficientes. Almacenada como retinol, sufre la acción hidrolítica de la retinil éster hidrolasa. Después de la hidrólisis, el retinol es directamente transferido al sitio de unión de almacenamiento del complejo a la proteína ligadora de retinol para ser transportado a otros tejidos. La proteína ligadora de retinol es el acarreador primario del retinol en el plasma y puede existir en forma holo o apo.6,7
En bovinos en pastoreo 52 se registran en el hígado concentraciones de 7.01 µg/ g, y en animales suplementados durante 30 días con β-caroteno 49 se registran concentraciones de 8.1 µg/g.
Despigmentación de la grasa en la actualidad
Para tratar de eliminar la pigmentación amarilla de la grasa, se señala53 que mantener a los animales durante periodos de 28 a 56 días en corral disminuye el color de aquélla, de manera que resulta aceptable para el mercado canadiense. Forrest encontró que tras 28 días de mantener a los animales en corral se disminuye la concentración de β-caroteno en grasa de 0.28 a 0.17 µg/g.
Sin embargo, se desconoce si el cambio de color al alimentar a los animales con dietas con base en granos se debe a la movilización y oxidación de los carotenoides de los depósitos grasos, a la dilución del efecto por la engorda o a ambos.13
Otro tipo de propuesta54 es el uso de lipoxigenasas de pasta de soya a nivel ruminal (las cuales se sabe que decoloran a los carotenoides); sin embargo, no se han obtenido resultados positivos.
En investigaciones recientes55 se probó la suplementación con vitamina A para tratar de reducir el depósito de β-caroteno en el tejido adiposo. Los resultados obtenidos muestran que dicha suplementación reduce la concentración hepática de β-caroteno entre 40% y 48% e incrementan la concentración de retinol a este nivel; sin embargo, el color de la grasa y la concentración de los carotenoides en tejido adiposo no se ven afectados con este tipo de tratamientos.
Conclusiones
Hasta el momento se encuentran dilucidados algunos de los factores metabólicos responsables del depósito de carotenoides en el tejido adiposo de los bovinos. Así está demostrado que el β-caroteno y la luteína no son degradados a nivel ruminal, ni en especies rumiantes que pigmentan la grasa de amarillo (bovinos), ni en aquellas que son de grasa blanca (caprinos) y que la tasa de desaparición de β-caroteno a nivel ruminal está asociada a la tasa de desaparición de la materia seca y del contenido celular.
Se ha observado que el intestino posee una amplia capacidad de absorción de β-caroteno y que los tejidos hepático y adiposo poseen una capacidad limitada de almacenamiento de retinol.
En cuanto a la actividad intestinal de la enzima 15,15´dioxigenasa, se ha observado que en caprinos es cinco veces mayor, comparada con la de bovinos, esto último podría explicar las diferencias en la coloración de la grasa entre ambas especies.
Los mecanismos por los cuales el β-caroteno se deposita en el tejido adiposo de ciertas especies no están completamente dilucidados; en este sentido es necesario realizar más los estudios sobre la cinética de la enzima 15,15´dioxigenasa, para comprender el mecanismo de acción de dicha enzima y sus diferencias entre especies de grasa amarilla y blanca. Asimismo, es prudente estudiar el transporte y recambio del β-caroteno depositado en el tejido adiposo y en el hígado, para conocer en qué proporción contribuye este β-caroteno depositado a las reservas de vitamina A.
Una vez que los procesos metabólicos se encuentren mejor dilucidados, será necesario instrumentar procedimientos o técnicas que permitan prevenir la pigmentación amarilla de la grasa en bovinos finalizados en pastoreo.

