











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
Huntington’s disease (HD) is a progressive neurodegenerative disorder whose incidence is approximately 0.38 per 100,000 individuals around the world annually. Europe, North America and Australia have the highest prevalence at 5.70 per 100,000 inhabitants.1
The genetic mutation responsible for HD is located on chromosome 4,2 specifically, gene IT15 contains an unstable expansion of CAG triplet repeats, coding for glutamine amino acid. Therefore, at the N-terminal region of the protein, named Huntingtin (htt), there is a polyglutamine (polyQ) tract.3 The number of CAG repeats is inversely related to the age of onset of the symptomatology. Consequently, there are three types of HD: childhood, juvenile and adult-onset.4 Adult-onset HD is the most common type, starting between ages 30 and 40, when humans show 40 or more CAG repeats. Juvenile HD represents 10% of cases; its onset occurs before age 20, and humans show more than 60 CAG repeats. Finally, childhood HD represents 5% of cases, and its onset occurs before age 10. These humans show between 80 and 100 CAG repeats, and the stiffness occurs from the early stages.5
Through genetic manipulation, it is possible to generate transgenic organisms, which may be used as animal models that simulate human diseases. Since HD occurs exclusively in humans, transgenic models are the best tool available for studying this disease. Animal models recreate the symptomatology of the disease and allow the study of cellular, molecular and behavioural aspects of HD; it is also possible to try out new treatments and drugs.
The organisms used for HD study are worms (Caenorhabditis elegans) fruit flies (Drosophila melanogaster), rats (Rattus norvegicus)6 swine (Sus scrofa domesticus)7 and, more recently, monkeys (Macaca mulatta)8 and sheep (Ovis aries).9 However, mouse models are the most used since the establishment of the polyQ transgenic mouse, which has facilitated the understanding of HD pathology, taking care that the inbreeding is maintained to conserve the stability and homogeneity of the strain.10
Transgenic HD mice have a fragment or the complete Htt gene randomly inserted into their genome, but the mice also express their own murine Htt gene. In contrast, in the HD knock-in model, the fragment or the complete Htt gene is inserted within the murine Htt gene. For this reason, the endogenous promotor controls the inserted gene that expresses the polyQ tract.11,12 Transgenic mouse models R6/1 and R6/2 express the first exon of human htt, with 116 and 144 CAG repeats in each one. The expression in both models is under the control of the human htt promotor.
The R6/1 line has a later onset and progression of the disease, showing a motor deficit at 18 - 20 weeks of age, loss of muscular mass at week 22 and a lifespan of up to 48 weeks.13 In contrast, the R6/2 line presents a motor deficit between weeks 5 and 6, weight loss at 8-9 weeks and a lifespan no longer than 13 to 16 weeks. Consequently, R6/1 is more suited for modelling adult-onset HD; R6/2, for modelling juvenile onset HD.14
Referring to neuropathology, line R6/1 mice present cerebral volume reduction at week 18 and neural atrophy but no neuron loss, while intraneural inclusions of htt start at 2 months. Meanwhile, R6/2 mice develop progressive neural disorders, such as cerebral weight loss at week 4, cerebral atrophy at 8 weeks, ventricular enlargement and astrogliosis. In addition, neuronal loss starts at week 12, and extensive htt aggregates are seen from day one.15 On the other hand, the R6/2 line develops diabetes as a consequence of transcriptional deregulation in pancreatic islets.16
Therefore, establishing and maintaining a transgenic mouse colony with similar biochemical changes to those of late-onset HD is of great importance. Line R6/1 is the most suited to study damage mechanisms and research the effectiveness of new neuroprotective treatments.
Materials and methods
Colony founder
A litter of transgenic mice (R6/1) with the genetic background B6CBA-Tg(HDexon1)61Gpb/1J and a Wild-type litter (Wt) with the genetic background B6CBAF1/J, both provided by Jackson laboratories, arrived in Mexico in 2014. We acquired 9 females and 19 males of the R6/1 line and 2 males and 2 females of the Wt line.
Reproduction of the R6/1 transgenic mice
The colony is maintained in the vivarium of Instituto Nacional de Neurología y Neurocirugía “Manuel Velasco Suárez” (INNN). All of the studies were performed following the internal regulations of the vivarium under the “Especificaciones técnicas para la producción, cuidado y uso de los animals de laboratorio” (NOM-062-ZOO-1999, 2001) and the approval of the Ethics Committee (CICUAL/SO/VI/22617/017/2017). The reproduction of the R6/1 transgenic mouse colony started with four mating boxes containing one male and two females in each one, whereas the mating of the Wt colony started with one mating box containing one male and two females. The resulting litters were used to maintain the colony growth and perform different experiments. The subsequent generations were maintained by inbreeding, and the offspring averaged 4 to 6 mice per litter. Weaning was performed at 3-4 weeks of age. Since the htt protein mutation of R6/1 transgenic mice follows a Mendelian inheritance pattern, different genotypes depending on the parental cross are expected.
Mouse maintenance
We established the necessary conditions to maintain strict control of the macro- and micro-environmental variables as well as the conditions for the sanitary protection of the colony. Animals were kept under the following macro-environmental conditions: 22 ± 2 ºC, 55-65% humidity and light/dark periods of 12/12 hours for each one. The micro-environmental conditions consisted of cages, bedding, food, water and hygienic measures. Mice were allocated in polycarbonate cages with stainless steel lids and electrostatic filters, which were individually marked with cards. The bedding was composed of wood shavings, which were 100 % natural fibre, UV radiated, 95 % dust-free and autoclaved. The animals were given access to food pellets and water ad libitum. Water was placed in 500 mL drinking bottles with neoprene caps and stainless-steel pipettes. The administered food was a constant formula of Labdiet®. Mice were kept in groups of 2 to 5 individuals per cage, and the bedding was changed twice a week. Visual examination was performed on a daily basis to assess all of the features exhibited by the animals related to the disease phenotype and their genetic condition.
DNA extraction
The correct identification of each animal is of great importance when handling transgenic mice; therefore, we performed DNA extraction to determine the genotype of each individual of the colony. A biopsy from the tail, 0.5 cm long, was taken from every mouse at the age of 4 weeks. For tissue digestion, 500 µL of lysis buffer was added (Tris-base 1 M, pH 8.5; EDTA 0.5 M; SDS 20%; NaCl 1 M) and 1.25 µL of proteinase K (20 mg/mL) to each sample. The samples were agitated at 50°C overnight, and after the incubation period, each tube was centrifuged at 13,000 rpm for 5 minutes at room temperature. The supernatant was transferred to a clean tube, and 500 µL of isopropanol was added and mixed vigorously until precipitated DNA was observed. The resulting pellet was washed with 500 µL of 70% ethanol. The excess ethanol was eliminated, and the DNA pellet was left to dry for 5 minutes. To resuspend the DNA, we added the necessary distilled water, and the DNA integrity was tested in an electrophoresis gel (1% agarose). Quantification and purity was confirmed by a spectrophotometer.
Animal genotyping
Genotyping was performed by end-point PCR, amplifying the mutated fragment of the human Huntingtin gene using the primer (3´ GCA GCAGCAGCAGCA ACA GCC GCC ACC GCC) and (5´ CGG CTG AGG CAG CAG CGG CTGT). The conditions used for a final reaction volume of 25 µL were as follows: 2.5 µL of buffer solution (Tris-base 670 mM, NH4SO4 166 mM, MgCl2 20 mM, bovine serum albumin 1.7 mg/mL, beta mercaptoethanol 10 mM, pH 8.8), 2.5 µL of 100% DMSO, 1 µL of each primer 25 pmol/µL, 0.825 µL of MgCl2 50 mM, 1 µL of dNTPs 12.5 mM, 0.25 µL of Taq polymerase, 1 µL of extracted DNA (100-150 ng) and 12.92 µL of sterile distilled water. Additionally, two independent reactions were performed, one as a negative control with 1 µL of sterile water and the other as a positive control with DNA obtained from a patient from INNN. The cycling conditions were 1 cycle at 94°C for 2 minutes, 35 cycles at 94°C for 30 seconds, 65°C for 30 seconds, 72°C for 90 seconds and, finally, 1 cycle at 72° C for 10 minutes. The PCR products had a 110-base pair size and were analysed in a 3% agarose gel, which was dyed with 3 µL ethidium bromide solution (10 mg/mL) and observed with UV light.
Motor tests
The spontaneous motor activity test was performed using Auto-track Opto-Varimex 4 (Columbus Instruments) equipment. A mouse was placed for 10 minutes in a translucent acrylic box (42.2 × 42.5 × 20.5 cm) equipped with movement sensors for the analysis of different parameters: horizontal movements (total counts of x and y-axes combined) and vertical movements (total counts of the z-axis sensor) in addition to the map of the total walked distance. Coordination and equilibrium were evaluated through a rotarod test, which consisted of training mice for three consecutive days at different constant speeds (10, 20 and 30 rpm); the test was performed on the fourth day. In the test, mice were put through two ramp assays at speeds from 2 to 40 rpm for 5 minutes. Latency of falling from the rotarod and the speed of the rotarod at that moment were recorded for each mouse, and the average of the two assays was used for the statistical analysis. For both of the tests, the equipment was cleaned between individuals to avoid added effects caused by any odours of the previous animal.
Statistical analysis
All results are expressed as the mean ± standard error. The analysis was performed with a two-factor ANOVA with a Tukey or Šidak post hoc test depending on the analysed test. Two-way ANOVA was selected to compare between genotypes and different time points, while Tukey and Šidak tests were chosen to compare all groups between them or some groups within a set (two genotypes at a single time point). A P < 0.05 value was considered statistically significant. All the analyses were performed using Prism 6.0 software (GraphPad).
Results and Discussion
Mouse genotyping by PCR was very reliable given that differentiating between the Wt and transgenic mice is possible (Figure 1). In addition, the phenotype of transgenic animals that show physical characteristics and progressive neurological changes is evident. At 15 weeks of age, the R6/1 mice showed the feet-clasping posture when they were suspended by the tail for less than 2 minutes (Figure 2A). In contrast, wild-type mice did not show this posture (Figure 2B). Additionally, the physical changes of R6/1 mice versus those of Wt mice are obvious at week 35 (Figures 2C and 2D).
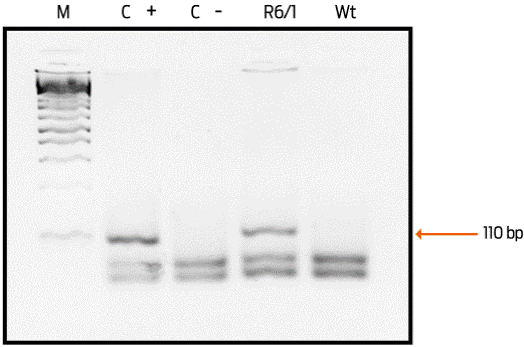
Figure 1 Representative image of the genotype identification. An agarose gel is shown representing the PCR products obtained to determine the presence of the mutated Huntingtin gene. Lane (C +): positive control showing the 110-base pair band (bp). Lane (C -): negative control of the reaction. Only the band of the expected molecular weight for an R6/1 transgenic mouse is shown, but it is not observed in the wild-type mouse. M = molecular weight marker.
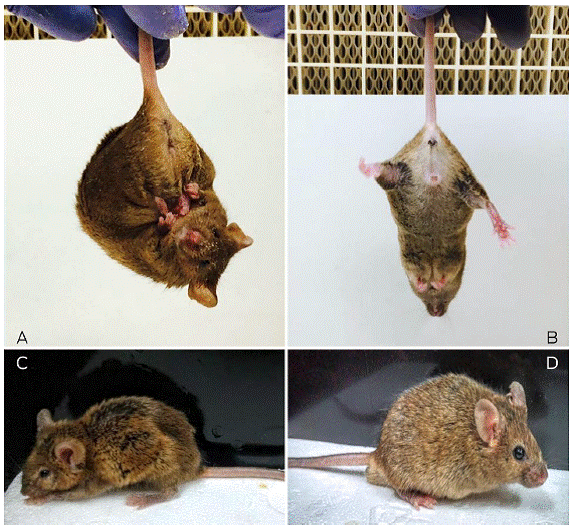
Figure 2 Representative image of animal’s phenotype. A) Male transgenic mouse R6/1 showing the typical feet-clasping posture. B) The feet-clasping posture is absent in the Wt mouse. C) The typical back-curvature posture and low weight are shown in the R6/1 male transgenic mouse. D) Wt mouse with normal fur and weight.
Bodyweight was measured at 10, 24 and 35 weeks. At week 10, the average weight of the Wt mice was 25.3 ± 0.77 g, while the average weight of the R6/1 mice was 26.5 ± 0.93 g. At week 24, the Wt mice had an average weight of 33.2 ± 0.81 g. On the other hand, R6/1 mice had an average weight of 29.9 ± 1.52 g, and at week 35, the Wt mice had an average weight of 40 ± 1.16 g in contrast to the R6/1 mice with an average weight of 27.7 ± 1.40 g. These data suggest that the R6/1 weight is significantly lower than the Wt weight at weeks 24 and 35 (Figure 3). Since the R6/1 line shows a loss of muscular mass around week 22, this transgenic model displays the previous reported physical traits.
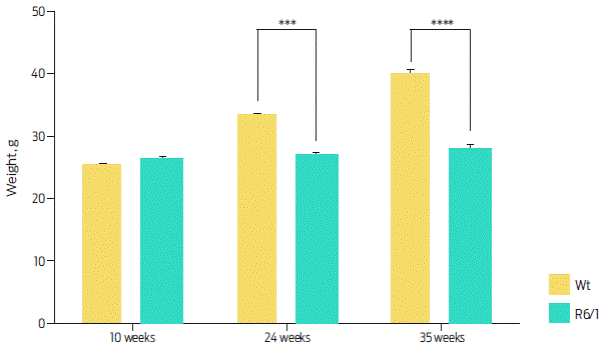
Figure 3 Corporal weight of R6/1 and Wt mice at 10, 24 and 35 weeks. R6/1 mice showed significant differences from week 24 in comparison with Wt mice. Two-way ANOVA, post hoc Šidak: time, F (2, 87) = 28.66, P < 0.0001; genotype, F (1, 87) = 42.36, P < 0.0001. Data are presented as the mean ± SEM, n = 4 per group, ***P < 0.001, ****P < 0.0001.
In addition to evaluating the genotype and phenotype of the R6/1 mice, the behaviour of the animals was monitored to ensure the presence of motor pathologies since there are reports on motor problems in murine transgenic models of HD. The motor tests were performed on three occasions (10, 24 and 35 weeks).
The open field test evaluates the exploration movements of mice. In this work, the number of horizontal and vertical movements were recorded for Wt and R6/1 mice. The 35-week-old R6/1 mice had significantly fewer movements, both horizontal and vertical, in comparison with Wt mice of the same age (Figures 4A and 4B). The distance map clearly shows how transgenic mice had less motor activity than Wt mice as time passed (Figure 4C). The motor activity of R6/1 and Wt mice was also evaluated by other parameters, such as exploratory movements, rest time and stereotypic movements. Despite the absence of a no significant difference in stereotypic movements between the R6/1 and Wt mice, there was an observed increase of 30% in stereotypic movements of the R6/1 mice at weeks 24 and 35. On the other hand, there was a significant increase in resting time (inactivity) in the R6/1 mice at week 35, while there was a significant decrease in the ambulatory activity of transgenic mice for HD in comparison with Wt animals (Table 1).
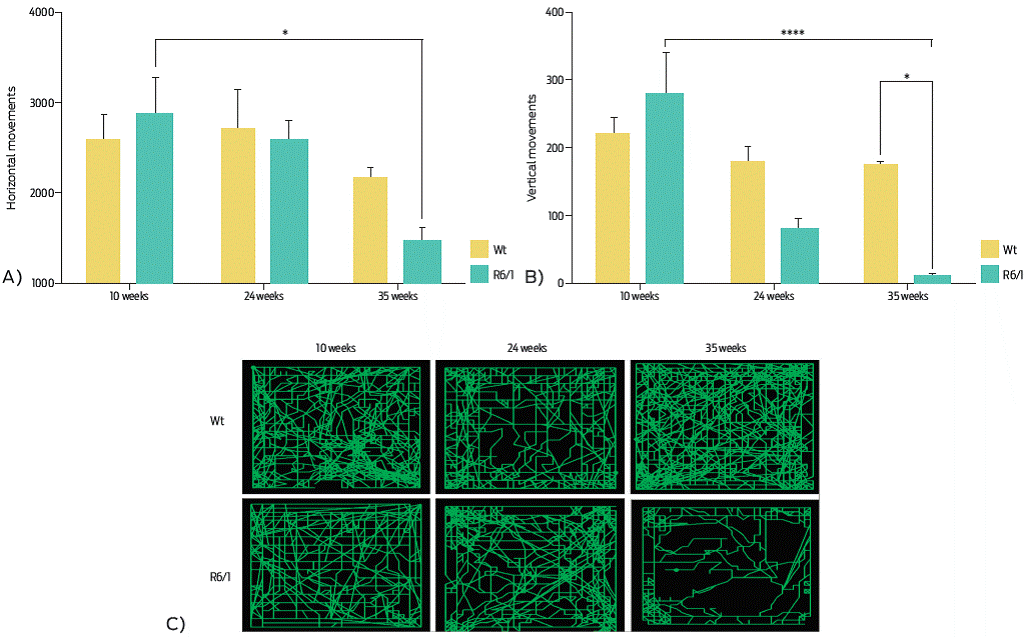
Figure 4 Motor activity. The number of horizontal (A) and vertical (B) movements diminish in a significant way across weeks in R6/1 mice; Wt mice showed constant motor activity. C) Map of the total walked distance. A) Horizontal movements. Two-way ANOVA, post hoc Tukey: time, F (2, 18) = 6.057, P = 0.0097; genotype, F (1, 18) = 0.6263, P = 0.4390. B) Vertical movements. Two-way ANOVA, post hoc Tukey; time, F (2, 18) = 15.02 P = 0.0001; genotype, F (1, 18) = 46.02 P = 0.0113. Data are displayed as the mean ± SEM; n = 4 per group, *P < 0.05, ****P < 0.0001.
Table 1 Spontaneous motor activity of the R6/1 transgenic and Wt mouse colony
| Weeks | 10 | 24 | 35 | |||
| Genotype | Wt | R6/1 | Wt | R6/1 | Wt | R6/1 |
| Resting time | 87.2 ± 8.6 | 93.5 ± 15.6 | 98.2 ± 20.1 | 107 ± 30.0 | 96.7±7.1 | 187.2 ± 15.0* |
| Stereotypic time | 102.5 ± 7.9 | 84 ±11.5 | 108.7 ± 10.5 | 144.5 ±12.7 | 117.7± 8.0 | 153.2 ± 14.4 |
| Ambulatory time | 410.2 ±16.2 | 422.5 ± 26.0 | 393 ± 25.7 | 348.5 ± 33.2 | 387.7±11.1 | 257.2 ± 10.0* |
Two-way ANOVA, post hoc Šidak; n = 4 by group, *P < 0.05. Resting time: time F (2, 6) = 2.639, P = 0.1506; genotype F (1, 3) = 25.80, P = 0.0147. Ambulatory time: time F (2, 6) = 6.688, P = 0.0297; genotype F (1, 3) = 28.22, P = 0.0130.
The rotarod test is the most commonly used procedure to evaluate motor coordination since it tests the ability of mice to maintain equilibrium on a rotating rod. Both R6/1 and R6/2 mice present progressive problems in the performance of this test.12 In the present work, we trained the Wt and transgenic mice aged 10, 24, and 35 weeks in the rotarod test. We clearly found a decrease in coordination, equilibrium and performance in the R6/1 mice throughout the weeks. Additionally, in all of the tested weeks, the performance of R6/1 mice was significantly lower than that of the Wt mice. At week 35, the difference was more evident: Wt mice remained stable on the rotarod for 127.3 ± 12.709 seconds and fell at an average speed of 8.5 ± 1.841 rpm, while R6/1 mice remained stable for 18.281 ± 7.034 seconds and fell at an average speed of 2.5 ± 0.340 rpm (Figure 5).
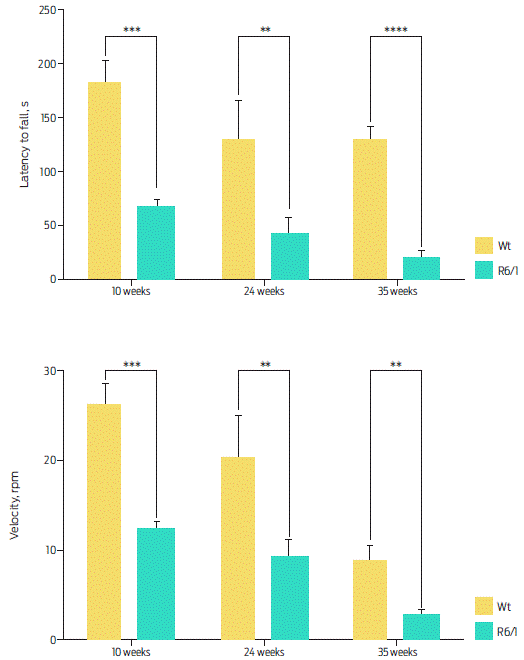
Figure 5 Rotarod activity. A) Duration. The R6/1 mice stayed less time on the rotarod compared with Wt mice in all tested weeks. Two-way ANOVA, post hoc Šidak; time, F (2, 34) = 5.773, P = 0.0069; genotype, F (1, 34) = 62.04, P < 0.0001. B) Speed. R6/1 mice fell from the rotarod at a lower speed than Wt mice. Two-way ANOVA, post hoc Sidak; time, F (2, 34) = 34.66 P < 0.0001; genotype, F (1, 34) = 46.02 P < 0.0001. Data are presented as the mean ± SEM; n = 10 per group, **P < 0.01, ***P < 0.001, ****P < 0.0001.
After the genotyping validation by PCR and the behaviour evaluation across the three ages of the transgenic model (R6/1) for Huntington’s disease, we observed, quantified and confirmed that the established colony showed the previously reported motor behaviour17.
The mice have remained healthy, and neither alteration nor contamination of the colony have been reported. At INNN, we currently have 4 generations (Table 2), which we continue breeding. To maintain the colony, it is necessary to have many mating boxes that contain mice from 5 to 15 weeks old. The obtained litters of the different genotypes are and will be employed in research and in the maintenance of the colony.
Table 2 Summary of the reproduction and genotype of the R6/1 transgenic and Wild-type (WT) mouse colony at INNN.
| R6/1 | Wt | Litter size | % R6/1 | % Wt | |
| Generation F1 | 8 | 12 | 20 | 40.0 | 60.0 |
| 1 | 12 | 13 | 7.7 | 92.3 | |
| 16 | 8 | 24 | 66.7 | 33.3 | |
| 3 | 5 | 8 | 37.5 | 62.5 | |
| Generation F2 | 2 | 5 | 7 | 28.6 | 71.4 |
| 9 | 10 | 19 | 47.4 | 52.6 | |
| 7 | 16 | 23 | 30.4 | 69.6 | |
| Generation F3 | 3 | 3 | 6 | 50.0 | 50.0 |
| 12 | 7 | 19 | 63.2 | 36.8 | |
| 11 | 11 | 22 | 50.0 | 50.0 | |
| 6 | 7 | 13 | 46.2 | 53.8 | |
| Generation F3 | 19 | 5 | 24 | 79.2 | 20.8 |
| 4 | 9 | 13 | 30.8 | 69.2 | |
| 10 | 11 | 21 | 47.6 | 52.4 | |
| Total | 111 | 121 | 232 | 44.7 | 55.3 |
Conclusions
The colony establishment of transgenic mice R6/1 at the INNN vivarium contributes to the research of HD in animal models, allowing the study of the mutant htt and its possible interaction with other proteins involved in neurological disorders or in neural protection. The successful reproduction of the colony reflects the good supervision and maintenance of the environmental conditions of the animals. This allows for less variability in the subjects and in the experimental response. In addition, this model has the advantage of showing progressive development of the pathology compared to other models, giving the opportunity to evaluate changes in different stages of the disease.

