











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Las aplicaciones de los principios de la ingeniería y de resistencias de materiales a la geometría de la diáfisis de los huesos largos, en el ámbito particular de la antropología física, han sido usadas extensivamente para inferir formas de comportamiento, especialmente de patrones de actividad, a través de restos óseos que provienen de las investigaciones arqueológicas (Bridges 1989; Brock et al. 1988; Fox et al. 2001; Ruff et al. 1982; Ruff et al. 1983; Ruff 1984; Ruff et al. 1984; Ruff et al. 1986). Por otra parte, también han sido usadas para mostrar la variabilidad de la sección transversal de los huesos largos, que está asociada con diferencias en el tamaño corporal (Ruff 1984), con la edad (Bouxsein et al. 1994; Lazenby 1990; Feik et al. 1997; Ruff 1981; Ruff et al. 1982), con la asimetría bilateral en el uso de los miembros (Fresia et al. 1990; Trinkaus et al. 1994, 2000), con el sexo (Feik et al. 1997; Mays 1999) y con actividad física relacionada con diferentes estrategias de subsistencia (Bridges 1989; Ruff et al. 1984).
En esta investigación, pretendemos comprender el vínculo existente entre las estrategias de subsistencias y la actividad física en restos óseos arqueológicos poscraneales, considerando solo el fémur. Utilizamos el método de la geometría de la sección transversal de la diáfisis de los huesos largos para estimar los parámetros biomecánicos de los mismos, así como su sensibilidad al estrés mecánico. Estudiamos grupos de población prehispánica del periodo Clásico (250 dC al 900 dC) y Posclásico (1000 dC a 1500 dC) de lo que hoy es el estado de Oaxaca y los comparamos con grupos de la península de Yucatán.
Se ha constatado una estrecha vinculación entre la estructura morfológica del cuerpo humano y la respuesta que se produce en el sistema óseo a las presiones mecánicas originadas en las actividades cotidianas que producen un estrés biomecánico susceptible de ser medido. Y que necesariamente nos obliga a plantear algunas preguntas que nos servirán como hilo conductor. ¿Podemos dar una explicación suficiente en torno a las diferencias y semejanzas biomecánicas que se presenten entre individuos de la costa y planicie yucateca y de las áreas montañosas oaxaqueñas? ¿En el caso de encontrar diferencias, éstas son causadas por sus diferentes formas de estrategias de subsistencias? ¿Qué elementos del entorno físico o social tienen mayor peso en la expresión de esas diferencias? ¿Cómo se reflejan las asimetrías sociales en las asimetrías corporales y viceversa?
Nos centramos en obtener un patrón de variabilidad intragrupal a nivel diacrónico y explicarlo en un marco de interpretación funcional como respuesta del sistema óseo a los procesos de actividad y movilidad física, como producto de las formas de organización social y actividades de subsistencia (caza, pesca, recolección, agricultura, entre muchas otras), entre grupos de población prehispánica oaxaqueña (N=38) de Monte Albán, con temporalidades del Clásico y Posclásico, que se comparan con fémures de mayas peninsulares (14 sitios, N=107), con temporalidades que van desde el periodo Clásico hasta el periodo Posclásico (900 a 1500 dC). También nos centramos en comprender las diferencias y semejanzas intra e intergrupales a partir de parámetros biomecánicos aplicados a los fémures, y obtener un patrón general de variación de su geometría transversal entre grupos de individuos de distintos ambientes ecogeográficos, como sería la costa, planicie yucateca y áreas de los valles centrales oaxaqueños.
Se ha visto que las propiedades de la sección transversal muestran diferencias asociadas con estrategias de subsistencia y actividad física, sexo y tamaño corporal. Es así que el estudio de la variación intrapoblacional de la geometría de la sección transversal de los huesos largos permite relacionar la variación geométrica con diversos factores mecánicos; por ejemplo conocer si hay diferencias sexuales durante los procesos de actividad y movilidad física. Desde una perspectiva antropológica, las diferencias intrapoblacionales en la estructura ósea nos pueden ayudar a reconstruir el comportamiento humano en medios ambientes específicos de las poblaciones prehispánicas oaxaqueñas y mayas peninsulares.
Marco teórico
Los modos de producción implican un acceso diferencial y desigual a los alimentos, esto se puede analizar por medio de datos bioarqueológicos del estado nutricional, valorados con los marcadores de estrés episódico o crónico que pueden delatar pautas asimétricas en el consumo de alimentos (Cadena 2016). Los huesos son capaces de reflejar las huellas de uso del cuerpo en las actividades cotidianas, como microadaptaciones fenotípicas a causa de la fuerza de trabajo, estas relaciones están mediadas por diversas formas de apropiación de los recursos naturales necesarios para su reproducción biológica y crean diferentes formas de organización social. Las actividades físicas que el individuo desarrolla en vida y que cubren un espectro amplio de comportamientos y actitudes a nivel productivo y recreativo que le permiten enfrentar los embates ambientales cotidianos que pueden conlle- var a serias repercusiones sobre su estado de salud merecen especial atención, ya que los procesos de ajuste corporal ocurren constantemente; cabe aclarar que el cuerpo humano tiene un número limitado de grados de libertad acotado por su propia morfología para realizar sus actividades físicas que son bastantes diversas (Stock y Pfeiffer 2001; Stock y Jay 2005; Macintosh et al. 2014, 2017).
Al igual que en el sistema óseo queda una “huella metabólica” susceptible de ser analizada y que aporta información importante sobre el estado nutricio de las poblaciones desaparecidas, éste también refleja una “huella mecánica”, reflejo de la carga, actividad y movilidad física, susceptible de ser medida a través de variables biomecánicas.
Así, por ejemplo, se ha encontrado, respecto a los miembros inferiores y para muestras europeas, un decremento en la resistencia sobre el plano AP a la mitad del fémur, lo cual se ha interpretado como una reducción de la carga física a través del tiempo; la evidencia paleontológica combinada con datos arqueológicos sugiere un medioambiente en degradación que provocó que el tamaño de los territorios habitables se redujeran, lo que ocasionó una disminución en los entornos ecológicos y en las estrategias de subsistencias y, por ende, una reducción en la movilidad para entornos geográficos cerrados (Trinkaus et al. 1994). Recientemente se ha investigado este fenómeno en poblaciones de Europa central que practicaban la agricultura durante la segunda parte del sexto milenio, donde la diversidad de actividades se amplió, pero la movilidad se redujo y los individuos se hicieron más sedentarios. En estas poblaciones, la diáfisis del fémur presenta una forma circular y una robustez marcada en ambos sexos (Macintosh et al. 2014, 2017).
La esquematización antes expuesta no significa que bajo las mismas causas siempre encontraremos los mismos efectos, en realidad, el conjunto de actividades físicas desarrolladas por un individuo durante toda su vida, y para el caso específico de las poblaciones antiguas de México, es algo mucho más complejo que la clasificación simple utilizada aquí (Fedick 1996; Flannery 1982; Melgar 2008); puesto que la esquematización está modulada por el tipo de sociedad en la que se desenvuelve el individuo y por el nivel de desarrollo de las fuerzas sociales y técnicas de producción, las cuales intensifican o minimizan la acción de los impactos sobre todo el organismo y en especial sobre el sistema óseo, ocasionando respuestas de diversas magnitudes que son susceptibles de ser medidas en los huesos.
Tradicionalmente, los indicadores de actividad física que se han utilizado se pueden clasificar en los siguientes rubros (Nadal y Morador 2002): a) Modificaciones de las regiones articulares, relacionadas con el desgaste-degeneración de las articulaciones o artrosis; b) Modificaciones no articulares, son remodelaciones óseas en las regiones de inserción muscular sometidas a presión, denominadas entesopatías; c) Modificaciones estructurales, son las alteraciones en la estructura interna del hueso que modifican su geometría cuando éste es sometido a presión mecánica. Este último rubro es el que nos interesa tratar en el contexto de la actividad física de los grupos prehispánicos mayas-yucatecos y oaxaqueños.
Marco Geográfico
Yucatán
El territorio que ocupa el actual estado de Yucatán se ubica en la península del mismo nombre. Es una losa caliza, casi plana, de 525 km de longitud y de 322 km de anchura que se inclina de sureste a noreste con una superficie total de aproximadamente 150,000 km2. Con algunas alturas que oscilan entre los 10 y 130 msnm. Colinda al oeste con Tabasco, al sur con Chiapas y con la nación de Belice y los departamentos del Petén e Izabal de Guatemala.
Por consiguiente, la península se puede dividir en tres grandes zonas fisiográficas (Miranda 1976; Lugo et al. 1992; Chiappy 2000), (Figura 1): a) El macizo de Campeche y zonas aledañas. Comprende el estado de Campeche, el sur de Yucatán y el oeste de Quintana Roo. Se trata de una formación miocénica que constituye la parte más elevada de la península. Con relieves moderados, con dolinas de gran extensión, cuyo fondo está recubierto por una espesa capa de arcilla negra fértil. b) La planicie del Caribe y el noroeste. Abarca el norte de Belice, el este y norte de Quintana Roo, y el este y sudeste de Yucatán. Es una región calcárea oligocénica de sedimento fluvial. c) La planicie del norte y el noroeste. Está constituida por la región central de Yucatán y conformada por terrenos calcáreos del Pleistoceno con excepción de la costa norte de Campeche (Miranda 1976).
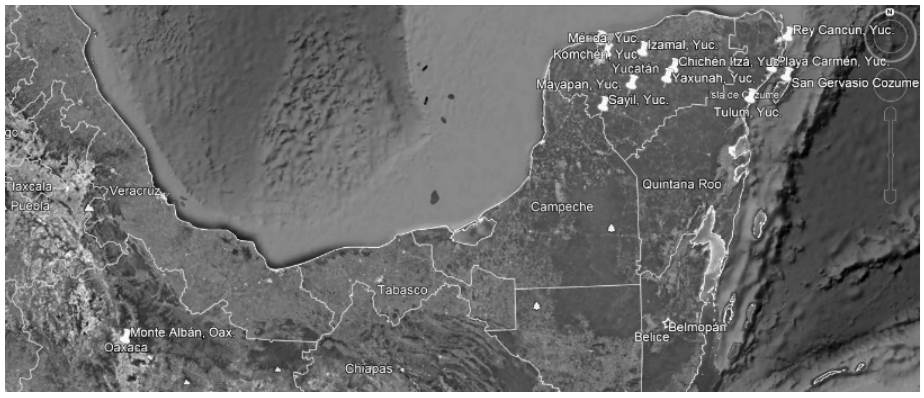
(fuente: Datos propios 2020)
Figura 1 Distribución relativa de la muestra de Yucatán (N=107, 14 sitios); según periodo cronológico y sexo.
En estas dos últimas regiones se ubican los sitios arqueológicos de donde proceden los materiales óseos de la península de Yucatán (Chichen Itzá, Dzibilchaltún, Izamal, Mayapán, Mérida, Rey Cancún, Rescate Súper, San Gervasio Cozumel, Sayil, Tulum, Xkipche, Playa del Carmen, Komchén, Yaxyunah).
Oaxaca
Oaxaca constituye una de las cinco grandes regiones culturales de Mesoamérica. Su extensión es cercana a los 100,000 km², con una altura sobre el nivel del mar que fluctúa desde una faja costera y algunas zonas de cuencas bajas inferiores a los 500 m, hasta la parte superior de algunas montañas que alcanzan alturas cercanas o mayores de 3,000 m. Su territorio se ubica en el sureste del país, colinda al norte con Puebla y Veracruz, al este con Chiapas, al sur con el Océano Pacífico y al oeste con Guerrero.
Este territorio comprende cinco provincias fisiográficas: la provincia Sierra Madre del Sur, la provincia Cordillera Centroamericana, la provincia Llanura Costera del Golfo del Sur, la provincia Eje neovolcánico y Sierras de Chiapas y Guatemala. Pero, para obtener una visión más detallada se debe dividir en las siguientes áreas: Valle de Oaxaca, Mixteca alta, Mixteca baja, Mixteca de la costa, Costa, Istmo oaxaqueño, Sierra sur, Chimalapas, Región mixe, Sierra zapoteca o Sierra Juárez, Chinantla, Mazateca y Cañada (Winter y Sánchez 2014: 3-4).
Esta primera provincia, que constituye la mayor extensión del territorio (79.57%), se subdivide en seis provincias: Cordillera costera del sur, Sierras orientales, Sierras centrales, Mixteca alta, Costas del sur y Sierras y Valles de Oaxaca.
Como se puede ver en estas breves descripciones fisiogeográficas de Oaxaca y de la península de Yucatán, en Oaxaca tenemos una orografía compleja que nos permite apreciar una región bastante diversa en lo geográfico, ecológico y social, totalmente diferente a la península de Yucatán. Otra diferencia importante, desde la perspectiva del entorno natural, podría ser la altitud; mientras que la península está en promedio a 10 msnm, la región oaxaqueña está entre los 500 y 3000 msnm. Entonces, podríamos señalar que la adaptación biológica, y en específico la biomecánica de estos grupos, es diferente debido a su ambiente geográfico montañoso y contrastante, pero precisamente lo que pretendemos demostrar en esta investigación es la mayor influenza del entorno sociopolítico en la plasticidad ósea (Figura 2).

Materiales
Península de Yucatán
Los materiales que se utilizan provienen de distintos rescates y proyectos de investigación arqueológica que se han realizado en la península en los últimos cincuenta años. La totalidad de sitios se ubican en distintas regiones geográficas de la península y se encuentran bajo resguardo en la sección de antropología física del Centro INAH Yucatán, en la ciudad de Mérida; comprenden una serie de colecciones óseas provenientes de entierros completos (individuales y múltiples), explorados principalmente en la zona costera de Playa del Carmen en unidades habitacionales y osarios con distintos grados de conservación (algunos completos y otros restaurados), y son representativos tanto de la costa como de la planicie yucateca. Un resumen del total y los parciales utilizados, por sitio arqueológico y cronología, se muestra en el Cuadro 1 y Figura 1.
Cuadro 1 Distribución de la muestra de Yucatán; según, sitio, cronología* y sexo
| Sitio | Cronologia | Masculinos | Femeninos | Total | |||
|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | ||
| Chichén | Clásico | 3 | 2.80 | 3 | 2.80 | ||
| Dzibilchaltún | Clásico | 5 | 4.67 | 3 | 2.80 | 8 | 7.48 |
| Izamal | Clásico | 1 | 0.93 | 1 | 0.93 | ||
| Mayapan | Clásico | 4 | 3.73 | 1 | 0.93 | 5 | 4.67 |
| Mérida | Clásico | 3 | 2.80 | 1 | 0.93 | 4 | 3.74 |
| Rey Cancún | Clásico | 3 | 2.80 | 3 | 2.80 | ||
| Rescate Super | Clásico | 4 | 3.74 | 3 | 2.80 | 7 | 6.54 |
| San Gervasio Cozumel | Clásico | 6 | 5.61 | 1 | 0.93 | 7 | 6.54 |
| Sayil | Clásico | 7 | 6.54 | 2 | 1.87 | 9 | 8.41 |
| Tulum | Clásico | 1 | 0.93 | 1 | 0.93 | ||
| Xkipché | Clásico | 1 | 0.93 | 1 | 0.93 | ||
| Playa Carmen | Postclásico | 42 | 39.25 | 42 | 39.25 | ||
| Komchen | Postclásico | 2 | 1.87 | 2 | 1.87 | ||
| Yaxuna | Postclásico | 14 | 13.08 | 14 | 13.08 | ||
| Total | 96 | 89.72 | 11 | 10.28 | 107 | 100 | |
* Desde 300 hasta 1500 dC. Fuente: Datos propios 2020.
La mayoría de los fémures analizados provienen de esqueletos explorados en osarios y el resto en estructuras habitacionales. Por el criterio de edad se eligieron fémures de adultos con una edad comprendida entre los 25 y 45 años.
Para la asignación de edad se emplearon principalmente los cambios que sufrieron tanto la sínfisis púbica como la superficie auricular propuestas por Todd (Loth e Iscan 1989; Meindl y Lovejoy 1985). Para la determinación del sexo se usaron principalmente los parámetros para la pelvis y el cráneo, así como el grado de robustez en miembros superiores e inferiores (Krogman e Iscan 1986). Es pertinente aclarar que la muestra se depuró de un total de 200 fémures, eliminando aquellos huesos largos que presentaban malformaciones, alteraciones por procesos diagenéticos o algún tipo de rasgo patológico como procesos artríticos, eburnación e inflamación perióstica, entre otros, porque podían alterar la estimación de los parámetros biomecánicos. Para la ordenación del material, en ambos sexos y solo para adultos, se aplicó la clasificación de Buikstra and Ubelaker (1994: 9).
En la clasificación por sexo, e independientemente de la cronología, se aprecia que 89% (96/107) de los individuos son de sexo masculino y el porcentaje de individuos de sexo femenino es de 10% (11/107).
Con la finalidad de abarcar el mayor espacio temporal, se incluyeron materiales del Clásico (300 al 900 dC) y del Postclásico (900 al 1500 dC), siendo escasos para el periodo Preclásico. Para el primer periodo se tienen 45.79% de fémures (49/107) y para el segundo periodo 54% (58/107), con base en la información de los Cuadros 3 y 4. Los sitios fueron elegidos inicialmente en función de la existencia del material óseo resguardado y posteriormente se hizo una clasificación en función de la región geográfica, esto es, si los individuos provenían del área costera o de la planicie peninsular.
Por último, en resumen, tenemos la muestra total distribuida por región geográfica, cronología y sexo, presentada en una tabla con dos entradas para una mejor visualización, esto nos permite saber cuántos individuos de la costa y de la planicie son masculinos y femeninos, así como la cronología a la que pertenecen (Cuadro 2). Del total de 107 fémures, 35 son de individuos de la costa y de sexo masculino (representan 32% del total de la muestra); 61 son de la planicie y representan 57% del total; 5 fémures son de individuos femeninos que vivieron en la costa (5% de la muestra) y 6 en la planicie (5%). Según la cronología en función del sexo, para el Clásico se tienen 38 fémures masculinos (35%) y 58 para el Postclásico (54%); finalmente hay 11 fémures de sexo femenino para el Clásico (10%) y ninguno para el Postclásico.
Cuadro 2 Distribución de la muestra de Yucatán (N=107, 14 sitios); según sexo, geografía y período cronológico
| Sexo | |||||||
|---|---|---|---|---|---|---|---|
| Masculinos | Femeninos | Total | |||||
| N | % | N | % | N | % | ||
| Región geográfica y Cronología | Costa | 35 | 32.71 | 5 | 4.67 | 40 | 37.38 |
| Planicie | 61 | 57 | 6 | 5.6 | 67 | 62.6 | |
| Total | 96 | 89.71 | 11 | 10.27 | 107 | 100 | |
| Clásico | 38 | 35.51 | 11 | 10.29 | 49 | 45.79 | |
| Posclásico | 58 | 54.2 | 58 | 54.2 | |||
| Total | 96 | 89.71 | 11 | 10.29 | 107 | 100 | |
* Datos propios del área Maya peninsular. Fuente: Datos propios 2020.
Oaxaca
Esta muestra está conformada por dos colecciones: una proveniente del sitio arqueológico de Monte Albán (N=30) y la otra de fémures esgrafiados depositados en el Museo de la casa del Mendrugo (N=8), procedentes de los valles centrales, Monte Albán (Olivares 2021) (Figura 1).
Colección de Monte Albán
Este sitio se encuentra ubicado en la región fisiográfica de los valles centrales, a una altitud de más de 2000 msnm en la región de las Tierras Altas Tropicales. La muestra ósea procede de las temporadas de 1972 a 1973 y de 1992 a 1994, del Periodo Clásico (Figura 2).
De este sitio se seleccionaron 30 fémures, 20 masculinos y 10 femeninos, en un estado de conservación bueno; provienen de contextos arqueológicos primarios, de unidades habitacionales construidas alrededor de la plaza mayor de Monte Albán.
Algunas características del contexto son; entierros primarios, la mitad de los individuos fueron depositados en posición decúbito dorsal extendido al interior de fosas rectangulares con paredes y techos de lajas (cistas) (Figura 3). Con relación a las características bioantropólogicas, la mayoría presentó afecciones bucales, hay presencia de trepanación craneal en dos individuos (entierros 26 y 28), tres casos de deformación craneal (entierros 24, 26 y 28) y un caso de modificación dental (entierro 44) (Winter et al. 1995). Los objetos asociados a los individuos son en su mayoría de uso doméstico.

Colección de huesos largos esgrafiados
Los huesos largos esgrafiados son parte de la Colección de la Fundación Casa del Mendrugo en Puebla que está conformada por huesos humanos, caracoles trompeta y piedras esculpidas de origen zapoteca; en su dimensión temporal, corresponden al periodo Monte Albán IIIB-IV (700-1100-1521 dC), provenientes de los valles centrales de Oaxaca. Recientemente un estudio isotópico del esmalte dental ha demostrado que los individuos comparten la misma huella isotópica de Monte Albán (Olivares 2021) por lo que se agrupa a esta gran muestra. La colección está conformada por 17 huesos largos (Figura 4). Se identificaron ocho fémures que corresponden a individuos adultos, dos derechos y seis izquierdos, cinco tibias izquierdas y dos húmeros izquierdos. Como elementos generales de diagnóstico para la estimación probable del sexo se siguieron los criterios de Scheuer y Black (2000) y William M. Bass (2005), se evaluó el tamaño y la robustez del hueso, ya que no se cuenta con el resto del esqueleto y faltan los extremos epifisíarios de los huesos. El NMI en esta serie es de ocho individuos, de acuerdo al número de fémures izquierdos; ninguno de los elementos fue pareable.

Estos huesos largos esgrafiados son un claro ejemplo de la utilización de elementos óseos con propósitos ceremoniales. Pudieron cumplir la función de cetros utilizados por personajes importantes en actos de gran relevancia social (Serrano et al. en prensa).
Debido a la manipulación póstuma de tales huesos, nos vimos limitados para calcular la longitud máxima del fémur, talla y masa corporal. Previamente realizamos algunas pruebas de imagen para estimar si estos relieves no afectarían el cálculo de los parámetros biomecánicos y resultó nula la afectación, por esta razón decidimos sumarla a la colección de Monte Albán. Agregamos a esta prueba una de orden estadístico al calcular sus estadísticos normales (media y desviación estándar) y comparar estos valores con los valores medios de los fémures de Monte Albán, con la idea de saber si teníamos una colección morfológicamente homogénea o heterogénea. A continuación, se presenta la distribución de las muestras de ambas regiones geográficas y culturales (Cuadro 3).
Cuadro 3 Distribución de las dos muestras (Yucatán y Oaxaca) según territorio, geografía, sexo y lado
| Territorio | Geografía | Cronología | Sexo | Masculino | Femenino | Total | |||
|---|---|---|---|---|---|---|---|---|---|
| Lado | N | % | N | % | N | % | |||
| Yucatán* | Costa | Clásico | Derecho | 4 | 23 | 1 | 1 | 4 | 40 |
| Izquierdo | 5 | 23 | 1 | 1 | 6 | 60 | |||
| Total | 9 | 71 | 1 | 100 | 10 | 100 | |||
| Izquierdo | 22 | 71 | 1 | 100 | 23 | 68 | |||
| Total | 31 | 94 | 1 | 100 | 34 | 100 | |||
| Planicie | Clásico | Derecho | 29 | 81 | 6 | 67 | 35 | 78 | |
| Izquierdo | 7 | 19 | 3 | 33 | 10 | 22 | |||
| Total | 36 | 100 | 9 | 100 | 45 | 100 | |||
| Posclásico | Derecho | 8 | 100 | 1 | 100 | 9 | 100 | ||
| Total Yucatán | 75 | 84 | 14 | 15 | 89 | 100 | |||
| Oaxaca** | Valle | Clásico | Derecho | 10 | 48 | 5 | 56 | 15 | 50 |
| Izquierdo | 11 | 52 | 4 | 44 | 15 | 50 | |||
| Total | 21 | 100 | 9 | 100 | 30 | 100 | |||
| Posclásico | Derecho | 2 | 29 | 2 | 25 | ||||
| Izquierdo | 5 | 71 | 1 | 100 | 6 | 75 | |||
| Total Oaxaca | 7 | 100 | 1 | 100 | 8 | 100 | |||
| GRAN TOTAL | 103 | 81 | 24 | 19 | 127 | 100 | |||
* Fémures de 14 sitios peninsulares de Yucatán
** Fémures de Monte Alban y Casa del Mendrugo
Fuente: Datos propios 2020.
Objetivos
El primer objetivo es obtener un patrón de variabilidad intragrupal a nivel diacrónico y explicarlo en un marco de interpretación funcional como respuesta del sistema óseo a los procesos de actividad y movilidad física, como producto de las formas de organización social y actividades de subsistencia (caza, pesca, recolección, agricultura, entre muchas otras), entre grupos de población prehispánica oaxaqueña de Monte Albán (N=38), con temporalidades del Clásico y Posclásico, que se comparan con fémures de mayas peninsulares (14 sitios, N=107). Con temporalidades que van desde el periodo Clásico hasta el periodo Posclásico (900 a 1500 dC).
El segundo es comprender las diferencias y semejanzas intra e intergrupales a partir de parámetros biomecánicos aplicados a los fémures, así como obtener un patrón general de variación de su geometría transversal, entre grupos de individuos de distintos ambientes ecogeográficos, como sería la costa, planicie yucateca y áreas de los valles centrales oaxaqueños.
Métodos y técnicas
Para este propósito se ha utilizado la tomografía computarizada o CT Scan, el cual es un método no invasivo para investigar la estructura de la sección transversal de los huesos largos (Brock et al. 1988; Bridges 1989).
Los fémures fueron escaneados con un equipo SIEMENS, Somaton sensation, CT/e helicoidal, dual multicorte, éste obtiene dos imágenes por corte virtual, de alta resolución y de construcción tridimensional. Las imágenes son tomadas cada 2 mm, en dirección proximal-distal, con una resolución de 512 x 512 pixel (pixel=12bit).
Posteriormente las imágenes son enviadas a una estación de trabajo donde son almacenadas en discos compactos bajo un formato comprimido en JPG o TIFF. De aquí se procede a leerlas en una PC con el programa de NIH-Image, que utiliza una macro escrita en Basic para el cálculo de los parámetros biomecánicos. Para los fines de esta investigación solo utilizaremos los resultados a 50% de la diáfisis.
Las características mecánicas del hueso y de la sección transversal (Figura 5) reflejan la rigidez (Strength) o dureza que le permiten al hueso resistir las fuerzas mecánicas que actúan sobre él. Estas fuerzas son aplicadas sobre los músculos y huesos, y pueden variar debido a un gran número de factores que incluyen diferencias en la masa y tamaño corporal, y se han clasificados en tipos (agricultura, pesca, horticultura, entre otras) y niveles de actividad física (caminar, correr, trepar; entre otras) (Ruff y Hayes 1983).

Así, tenemos que las fuerzas de cargas primarias que afectan al hueso son; la tensión, compresión, cizallamiento, flexión y torsión, o una combinación de ellas, tal como la torsión-compresión, torsión-cizallamiento o torsión-tensión. En la flexión y en la torsión de una estructura ahuecada, tal como un hueso largo, la magnitud del estrés mecánico es proporcional a la distancia del centro o del eje “neutral” del hueso. El eje neutral es donde el estrés es igual a cero; en este plano, el hueso tiene una gran resistencia a la fractura.
Una forma de cuantificar el estrés mecánico es calculando los siguientes parámetros biomecánicos (Figura 6); área total (AT), área cortical (AC), área medular (am), centroide (x, y), segundos momentos de inercia cercanos al centroide o eje neutral de carga Ix, Iy; segundos momentos de inercia (Imax, Imin) sobre los planos mediolateral (ml) y anteroposterior (AP), el ángulo de torsión (θ) medido en grados en sentido contrario a las manecillas del reloj a partir del eje mayor X; y el momento polar de inercia (J).

(elaboración propia)
Figura 6 Imagen digital de la sección transversal del fémur a la mitad de la diáfisis. Eje natural (línea gris continua). Ejes rotados (línea gris discontinua).
Para el análisis se usaron los parámetros básicos de las distribuciones normales (media, desviación estándar, coeficiente de variabilidad y error estándar), así como diversas pruebas estadísticas como el análisis de la varianza o distribución de F de Fisher y la T de Student.
Resultados y discusión
Variabilidad intragrupal (mayas-yucatecos)
En general, al comparar masculinos y femeninos (Cuadro 4), independientemente de la cronología, por medio de la prueba T, podemos apreciar que los valores correspondientes a los primeros momentos de inercia (AT y AC) que miden las áreas de la sección transversal del fémur presentan diferencias significativas y que los fémures de los hombres son más resistentes que el de las mujeres. Los segundos momentos de inercia, igualmente vinculados a procesos de actividad y movilidad física (momentos cercanos al eje, mínimos y máximos, torca e índices de Nordin) manifiestan valores más altos entre los hombres que entre las mujeres.
Cuadro 4 Prueba T y F, según sexo y geografía; Masculinos (N=52) y Femeninos (N=2) Mayas, Costa
| Masculinos | Femeninos | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | n | m | s | n | m | s | p (t)* | p(F)* |
| AT | 52 | 704.90 | 116.28 | 2 | 501.42 | 123.82 | 0.019 | |
| AC | 52 | 514.83 | 83.55 | 2 | 358.80 | 87.65 | 0.012 | |
| AM | 52 | 190.07 | 39.71 | 2 | 142.62 | 36.18 | 0.103 | |
| Ix | 52 | 21617.59 | 7071.52 | 2 | 10496.74 | 2732.32 | 0.032 | |
| Iy | 52 | 22635.28 | 7735.68 | 2 | 1113.47 | 7108.55 | 0.043 | |
| Imax | 52 | 24680.05 | 7689.67 | 2 | 12540.41 | 5323.13 | 0.032 | |
| Imin | 52 | 19556.29 | 6813.26 | 2 | 9069.78 | 4517.75 | 0.036 | |
| J | 52 | 44252.87 | 14270.69 | 2 | 21610.21 | 9840.87 | 0.031 | |
| Ix/Iy | 52 | 0.98 | 0.20 | 2 | 1.09 | 0.45 | 0.468 | 0.088 |
| Imax/Imin | 52 | 1.29 | 0.18 | 2 | 1.41 | 0.12 | 0.348 |
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; s=Desv. Tipica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.
De manera particular, es interesante señalar que es más elevado el valor de la magnitud de torsión en los masculinos que en los femeninos, lo cual nos indica que estuvieron sujetos a una mayor actividad física. Sin embargo, los femeninos presentan una movilidad mayor que los masculinos (F=0.088, p<0.001), con una forma de la sección transversal más redondeada entre los masculinos y más elipsoidal entre los femeninos. Los valores de tamaño son homogéneos, si bien la falta de diferencias podría deberse a la sub-representación de fémures femeninos. No obstante, desde la perspectiva de los parámetros biomecánicos, la historia es otra: hombres y mujeres responden de manera diferencial en las regiones costeras, a pesar de no presentar dimorfismo sexual acentuado. Esta aseveración es consistente con el ambiente socioeconómico que se vivía al final del Clásico y principios del Posclásico: cuando en el primero se produjo el colapso de los grupos en las zonas altas del sur de la península de Yucatán y del Petén guatemalteco, aumentó la actividad de trabajo físico y se redistribuyó la población hacia la costa.
Según Wescott (2004), la movilidad producida por migración es un causal próximo que influye en las diferencias en el dimorfismo sexual. A este dimorfismo sexual va aparejado una alimentación basada en el monocultivo y en la pesca, donde los hombres y mujeres tienen que producir de manera similar e intensiva.
Así, el diseño experimental aplicado a los datos demostró que, respecto a los periodos Clásico y Posclásico, las variables biomecánicas entre hombres y mujeres, medidas a través de las áreas o primeros momentos de inercia y referidas al tamaño absoluto y relativo de la sección transversal del fémur, fueron consistentes con el entorno sociopolítico más que con el entorno físico, porque la costa de la porción norte de la península de Yucatán no es un medio geográfico accidentado, sino relativamente plano y homogéneo. También lo fueron aquellas referidas a la resistencia y a la fractura, que miden la distribución de la masa ósea en el hueso; éstas nos proporcionan inferencias directas sobre la actividad física. Por último, fueron consistentes también las variables que están relacionadas con la forma de la sección transversal y que nos remiten al nivel de movilidad de los individuos.
Ahora, veamos cuál fue el comportamiento de estas mismas variables, pero en la planicie (Cuadro 5). Se aprecia que los parámetros biomecánicos son más estables y homogéneos, solo se obtuvo una diferencia significativa en el índice de movilidad, que involucra los planos máximo y mínimo, siendo ligeramente mayor la movilidad entre las mujeres que entre los hombres, patrón parecido a la muestra de la costa.
Cuadro 5 Prueba T de los parámetros biomecánicos, Mayas, Planicie
| Masculinos | Femeninos | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | n | m | s | n | m | s | p (t)* | p(F)* |
| AT | 44 | 704.30 | 121.13 | 9 | 655.50 | 144.57 | 0.291 | 0.413 |
| AC | 44 | 503.01 | 82.46 | 9 | 476.12 | 102.74 | 0.396 | 0.255 |
| AM | 44 | 201.28 | 49.51 | 9 | 179.38 | 45.67 | 0.227 | 0.676 |
| Ix | 44 | 20988.93 | 7406.95 | 9 | 19537.90 | 7607.23 | 0.596 | 0.724 |
| Iy | 44 | 21409.18 | 7607.28 | 9 | 18996.74 | 9286.41 | 0.407 | 0.330 |
| Imax | 44 | 23598.82 | 8345.69 | 9 | 21437.56 | 8773.20 | 0.486 | 0.578 |
| Imin | 44 | 18797.04 | 6112.32 | 9 | 17097.08 | 7893.79 | 0.473 | 0.150 |
| J | 44 | 42398.11 | 14097.18 | 9 | 38534.64 | 16437.83 | 0.469 | 0.391 |
| Ix/Iy | 44 | 1.00 | 0.22 | 9 | 1.11 | 0.33 | 0.207 | 0.228 |
| Imax/Imin | 44 | 1.25 | 0.18 | 9 | 1.33 | 0.32 | 0.344 | 0.068 |
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; S=Desv. Típica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.
En lo particular, se muestra un patrón dimórfico de los parámetros biomecánicos similar entre las dos muestras. En la planicie, a diferencia de la costa, las actividades están basadas en el monocultivo y en una actividad intensa constructiva, se presume que, tanto hombres como mujeres estuvieron sujetos a una serie de actividades físicas parecidas o actividades que involucraban el uso de las extremidades de manera similar; de allí la expresión semejante entre estos parámetros. Sin embargo, y de nueva cuenta, la movilidad fue la variable que presentó un valor mayor entre las mujeres que entre los hombres, al igual que en la costa.
Con base en lo arriba descrito, inferimos una mayor resistencia ósea entre la gente de la planicie, probablemente relacionada con una mayor demanda física sobre estos individuos que afectó de manera similar a ambos grupos. Sin embargo, los niveles de movilidad fueron constantes y no presentan diferencias significativas. Probablemente fueron afectados igualmente por caminatas intensas en terrenos irregulares, en individuos que no tuvieron otra forma de desplazarse mas que utilizando sus piernas. Aquí habría que considerar otras causales que podrían explicar las diferencias en la densidad ósea, por ejemplo, se ha visto una pérdida de masa ósea en relación con la edad, sobre todo en mujeres adultas maduras a causa de la maternidad, la disminución hormonal que acompaña la menopausia y las deficiencias metabólicas; en hombres jóvenes, la menor densidad ósea podría ser un indicativo de deficiencias nutricionales; también pueden existir factores genéticos vinculados con la disminución de la densidad ósea. De manera general, se ha asociado a la inactividad física y a déficits dietarios y nutricionales para ambos sexos (Suby et al. 2013).
En resumen, podemos afirmar dentro de los límites de la confiabilidad estadística que los individuos que vivieron en la costa y en la planicie son consistentes con los causales próximos, tales como la agricultura y construcción que se intensificaron en los dos periodos, a pesar de la redistribución de los centros de poder que fungieron como causales finales, lo cual provocó un mayor incremento de actividad física. Los masculinos de la costa presentan una mayor asociación significativa entre variables biomecánicas, resultado de causales finales relacionados con la redistribución, concentración y migración de población, que ocurrió en la transición del Clásico al Posclásico, aunados a una intensificación de las estrategias de subsistencias en zonas costeras.
Variabilidad intragrupal (Zapotecos de Monte Albán y Mendrugo)
Los resultados de la comparación de las medias para muestras independientes nos indican que las únicas variables con diferencias significativas fueron el área medular (F=3.778, p<0.063<0.01), con una media de 174 cm2 en los fémures de Monte Albán y de 164 cm2 en los fémures del mendrugo. Y en el índice de movilidad (F=4.392, p<0.046<0.01) en masculinos, y en femeninos solo tuvimos un caso, insuficiente para el cálculo comparativo. Las diferencias son mínimas, por ello decidimos incluirlo en el total de la muestra de los fémures de los valles centrales, con las reservas del caso, puesto que estamos diferenciando a estos grupos por características fenotípicas morfológicas sujetas a modificaciones medioambientales y, por tanto, no estables ni heredables. De esta manera hemos obtenido un total de 38 fémures.
Asimetría bilateral, independiente de sexo y cronología.
Según lo reportado por la literatura acerca de la asimetría corporal, la cual es influida por factores genéticos, medioambientales o de estilos de vida (Trinkaus et al. 1994; Churchill y Formicola 1997; Trinkaus 2000; Auerbach 2006), podríamos tener valores biomecánicos similares o diferentes en la misma persona o en el mismo esqueleto. Entonces, para discriminar este efecto, en primer lugar se calcularon los estadísticos medios de los parámetros biomecánicos, tanto de los fémures derechos como izquierdos (Cuadro 6), con la finalidad de apreciar la variabilidad intragrupal y posteriormente realizar las comparaciones pertinentes con otros grupos. Por consiguiente, usaremos los fémures de masculinos derechos (N=12) e izquierdos (N=16), y femeninos derechos (N=9) e izquierdos (N=5), para analizar la asimetría bilateral. Los resultados se presentan en el Cuadro 6, que muestra la aplicación del análisis de la varianza para muestras independientes (p<0.05, dos colas).
Cuadro 6 Prueba F de Análisis de la varianza para muestras independientes; Según lado, independiente de sexo y cronología; Asimetría bilateral de los parámetros biomecánicos; Zapotecos (N=37)
| Derechos | Izquierdos | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | n | m | s | n | m | s | p(F)* | p (t)* |
| AT | 17 | 610.35 | 166.47 | 20 | 611.14 | 140.86 | ||
| AC | 17 | 448.31 | 124.31 | 20 | 458.64 | 106.73 | ||
| AM | 17 | 162.03 | 46.72 | 20 | 152.50 | 41.71 | ||
| Ix | 17 | 17421.48 | 9640.52 | 20 | 19136.00 | 9360.86 | ||
| Iy | 17 | 17547.52 | 9859.15 | 20 | 16898.45 | 7572.92 | ||
| Imax | 17 | 19543.68 | 11074.53 | 20 | 20304.73 | 9310.74 | ||
| Imin | 17 | 15425.33 | 8322.24 | 20 | 15729.71 | 7487.32 | ||
| J | 17 | 34969.01 | 19345.37 | 20 | 36034.44 | 16448.58 | ||
| Ix/Iy | 17 | 1.01 | 0.17 | 20 | 1.14 | 0.24 | ||
| Imax/Imin | 17 | 1.25 | 0.14 | 20 | 1.31 | 0.23 | 0.023 |
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; S=Desv. Tipica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.
Como bien se aprecia, solamente hubo diferencias significativas en la magnitud del índice de movilidad que incluye los planos máximos y mínimos (F=5.66, p<0.023<0.05), con una forma ligeramente mayor del lado izquierdo debido quizá a un esfuerzo diferente durante el desplazamiento. En general, los parámetros biomecánicos no reflejan mayores diferencias entre los fémures derechos e izquierdos, la afectación debida a la asimetría bilateral fue insignificante, indicándonos una actividad física que influía equilibradamente e involucraba a los dos segmentos inferiores.
Dimorfismo sexual y parámetros biomecánicos
El sexo de un individuo está influido genéticamente, pero a nivel fenotípico intervienen multitud de factores que provienen del medioambiente social y físico, que dan lugar a una amplia variabilidad. El término dimorfismo sexual se refiere al grado en el que se manifiestan fenotípicamente las diferencias sexuales como resultado del rol fisiológico en la reproducción, adaptación al medio, configuración biológica, nutrición, hormonal, envejecimiento, la actividad física, entre otros factores. Para el cálculo del dimorfismo sexual se utilizan los valores de tamaño y forma, es decir, la talla y masa corporal. En esta investigación solo incluiremos el análisis de los parámetros biomecánicos, tanto los primeros y los segundos momentos de inercia, como los derivados para el cálculo de la movilidad.
Para este propósito calculamos los valores medios, según lado y sexo, independiente de la cronología, en el Cuadro 7 se muestran las comparaciones aplicando la prueba t de Student en fémures derechos.
Cuadro 7 Prueba T para muestras independientes; Según sexo y lado, independiente de cronología; Parámetros biomecánicos de fémures derechos; Zapotecos (Monte Albán y Mendrugo, N=17)
| Masculinos | Femeninos | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | n | m | s | n | m | s | p (t)* | p(F)* |
| AT | 12 | 667.72 | 152.72 | 5 | 472.65 | 114.60 | 0.02 | |
| AC | 12 | 488.05 | 119.73 | 5 | 352.95 | 79.27 | 0.04 | |
| AM | 12 | 179.67 | 38.78 | 5 | 119.70 | 37.70 | 0.01 | |
| Ix | 12 | 20337.28 | 9917.09 | 5 | 10423.57 | 3818.48 | 0.01 | |
| Iy | 12 | 20471.55 | 9970.48 | 5 | 10529.86 | 5312.55 | ||
| Imax | 12 | 22806.81 | 11420.28 | 5 | 11712.18 | 4827.93 | 0.06 | |
| Imin | 12 | 18002.02 | 8354.82 | 5 | 9241.26 | 4166.40 | ||
| J | 12 | 40808.83 | 19715.11 | 5 | 20953.43 | 8957.21 | ||
| Ix/Iy | 12 | 1.00 | 0.13 | 5 | 1.06 | 0.27 | ||
| Imax/Imin | 12 | 1.24 | 0.14 | 5 | 1.28 | 0.15 | ||
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; S=Desv. Tipica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.
Las diferencias entre masculinos y femeninos dentro del grupo de los zapotecos son evidentes. Los primeros momentos de inercia (AT, AC y AM), como primera aproximación en la comprensión de la actividad física, que tienen que ver con la resistencia del fémur a fracturarse y/o cizallarse, presentan diferencias significativas (p<0.05) en las áreas corticales y medulares, con mayor valor los fémures masculinos que los femeninos.
De la misma forma, los segundos momentos de inercia que muestran la composición interna del fémur para amortiguar fracturas y cizallamientos en los planos (x, y) muestran diferencias significativas. La magnitud del ángulo de torsión o momento polar de inercia presentó valores mayores en masculinos que en femeninos, congruente con una mayor actividad física. Los valores de movilidad no tuvieron diferencias significativas, hombres y mujeres tuvieron la misma capacidad de desplazamiento.
En los fémures izquierdos (Cuadro 8) podemos apreciar diferencias significativas entre masculinos y femeninos sin diferencias en la movilidad. Son muy similares a los fémures derechos, en ambos se presenta una heterogeneidad biomecánica, hay un cierto desequilibrio entre los sexos de este grupo. Los masculinos presentan una mayor actividad física. Queda pendiente el análisis de los húmeros y tibias de este grupo, para consolidar esta aseveración e inferir con mayor precisión el rol de actividades en los sexos.
Cuadro 8 Prueba T para muestras independientes; Según sexo y lado, independiente de cronología; Parámetros biomecánicos de fémures izquierdos; Zapotecos (Monte Albán y Mendrugo, N=20)
| Masculinos | Femeninos | ||||||
|---|---|---|---|---|---|---|---|
| Variable | n | m | s | n | m | s | p (t)* |
| AT | 15 | 659.40 | 122.60 | 5 | 466.35 | 81.81 | 0.00 |
| AC | 15 | 493.25 | 96.31 | 5 | 354.81 | 60.62 | 0.01 |
| AM | 15 | 166.15 | 36.92 | 5 | 111.54 | 26.39 | 0.01 |
| Ix | 15 | 21983.15 | 8943.83 | 5 | 10594.53 | 3828.24 | 0.01 |
| Iy | 15 | 19081.19 | 7282.38 | 5 | 10350.21 | 3915.03 | 0.02 |
| Imax | 15 | 23117.49 | 8853.95 | 5 | 11866.45 | 4327.73 | 0.02 |
| Imin | 15 | 17946.85 | 7231.05 | 5 | 9078.29 | 3088.81 | 0.02 |
| J | 15 | 41064.34 | 15610.34 | 5 | 20944.74 | 7262.99 | 0.01 |
| Ix/Iy | 15 | 1.17 | 0.23 | 5 | 1.05 | 0.26 | |
| Imax/Imin | 15 | 1.31 | 0.24 | 5 | 1.31 | 0.23 | |
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; S=Desv. Tipica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.
Como pudimos constatar, en ambos grupos están actuando similarmente los factores de estrés biomecánico y es mayor la similitud entre los que vivieron en la costa yucateca y los que vivieron en la planicie. Es congruente porque, en primer lugar, ambos grupos están sometidos a similares exigencias por la construcción arquitectónica y la agricultura. La intensidad de estos factores es diferente en calidad, cantidad, exposición, etcétera; pero con la misma dirección e intensidad, al final la respuesta ósea es parecida.
Mientras el medioambiente social es heterogéneo porque se realizan diversas tareas premeditadas y estandarizadas que tienen como finalidad satisfacer las necesidades de supervivencia, el sistema óseo responde de manera autoorganizada debido a una remodelación y estructuración constante (Sepúlveda 2017) con una serie de grados de libertad ligados al movimiento, esfuerzos y tensiones permanentes. Este proceso se da bajo la denominada Cadena Causal Biomecánica que se ajusta constantemente según el modelo del Mecanostato. Y que, de acuerdo con esta investigación, hemos planteado que el ambiente sociopolítico tiene una mayor influencia en las modificaciones en las respuestas biomecánicas de la estructura corporal, que el entorno físico.
Según la literatura citada (Ruff 1981; Bridges 1989; Wescott 2008), los valores de los parámetros biomecánicos que hemos obtenido en ambos grupos deberían corresponder a individuos que se desplazan en entornos montañosos o escabrosos; si bien esto sí ocurre en otros grupos, no es nuestro caso. De modo que recurrimos a otros factores que operan a escalas temporales cortas y que modifican a nivel ontogenético la plasticidad humana, produciendo con esta aptitud microadaptaciones fenotípicas que les permiten entrar en sintonía con su entorno social.
Variación intergrupal
Otro aspecto central en esta investigación que decidimos explorar es el comportamiento de las variables biomecánicas entre los mayas que habitaron en la costa y planicie, y los zapotecos que habitaron en los valles centrales. Los resultados de los parámetros biomecánicos se muestran en el Cuadro 9.
Cuadro 9 Comparación de los valores medios de los parámetros biomecánicos; Maya, Costa (N=51) y Zapotecos, valles centrales de Oaxaca (N=27) Masculinos
| Costa | Valle | ||||||
|---|---|---|---|---|---|---|---|
| Variable | n | m | s | n | m | s | p (t)* |
| AT | 51 | 704.59 | 117.41 | 27 | 663.1 | 134.09 | |
| AC | 51 | 514.53 | 84.35 | 27 | 490.94 | 105.2 | |
| AM | 51 | 190.05 | 40.1 | 27 | 172.16 | 37.65 | 0.059 |
| Ix | 51 | 21563.84 | 7131.15 | 27 | 21251.66 | 9239.93 | |
| Iy | 51 | 22663.12 | 7810.03 | 27 | 19699.13 | 8432.69 | |
| Imax | 51 | 24663.76 | 7765.28 | 27 | 22979.41 | 9869.9 | |
| Imin | 51 | 19547.15 | 6880.74 | 27 | 17971.37 | 7595.26 | |
| J | 51 | 44210.9 | 14412.77 | 27 | 40950.78 | 17195.19 | |
| Ix/Ix | 50 | 0.98 | 0.2 | 27 | 1.09 | 0.21 | 0.022 |
| Imax/Imin | 50 | 1.29 | 0.19 | 27 | 1.28 | 0.2 | |
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; S=Desv. Típica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.
Al comparar los valores medios entre fémures de individuos masculinos mayas que vivieron en la costa y zapotecos que vivieron en los valles centrales de Oaxaca, para muestras independientes en prueba t de dos colas y prueba F en la que se asume la igualdad de varianzas o prueba de Levene, encontramos un alto nivel de significancia.
Los primeros momentos de inercia am presentaron un nivel de significancia (t=1.914, p=0.059<0.05), con un valor del área medular mayor entre los mayas de la costa que entre los zapotecos de los valles centrales, esto significa una mayor resistencia a la fractura entre los mayas masculinos de la costa. El índice de movilidad en el eje principal del fémur (t=2.346, p=0.022<0.05), presentó valores absolutos bajos entre los mayas de la costa mientras que los índices máximo y mínimo de movilidad presentan valores altos, con una forma de la diáfisis más elipsoidal, propios de grupos agricultores. Entonces podemos afirmar que, entre masculinos mayas de la costa y zapotecos de los valles centrales, no hubo mayores diferencias en el resto de las variables; podríamos decir que en ambos grupos hay una homogeneidad biomecánica y morfológica, probablemente como resultado de la misma intensidad de los estresores biomecánicos en ambientes geográficos diferentes, puesto que los primeros vivieron sobre una topografía plana sin mayores relieves y los segundos en áreas de mayor altitud, en un valle con una topografía escabrosa.
Para el caso de femeninos, debido al reducido número de fémures y para evitar el error de muestreo Tipo II, aplicamos en lugar de la prueba de Student, la prueba de Fischer (F) que considera las varianzas de ambas muestras en lugar de N. Los resultados se muestran en el Cuadro 10, Figura 7.
Cuadro 10 Comparación de los valores medios de los parámetros biomecánicos; Mayas, Costa (N=2) y Zapotecos de los valles centrales de Oaxaca (N=10) Femeninos
| Costa | Valle | ||||||
| Variable | n | m | s | n | m | s | p (F)* |
| AT | 2 | 501.42 | 123.82 | 10 | 469.50 | 93.93 | |
| AC | 2 | 358.80 | 87.65 | 10 | 353.88 | 66.53 | 0.092 |
| AM | 2 | 142.62 | 36.18 | 10 | 115.62 | 30.98 | |
| Ix | 2 | 10496.74 | 2732.32 | 10 | 10509.05 | 3605.83 | 0.005 |
| Iy | 2 | 11113.47 | 7108.55 | 10 | 10440.04 | 4400.54 | |
| Imax | 2 | 12540.41 | 5323.13 | 10 | 11789.32 | 4323.22 | |
| Imin | 2 | 9069.78 | 4517.75 | 10 | 9159.77 | 3458.72 | |
| J | 2 | 21610.19 | 9840.88 | 10 | 20949.09 | 7687.87 | |
| Ix/IY | 2 | 1.09 | 0.45 | 10 | 1.05 | 0.25 | |
| Imax/Imin | 2 | 1.41 | 0.12 | 10 | 1.30 | 0.18 | |
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; S=Desv. Tipica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.
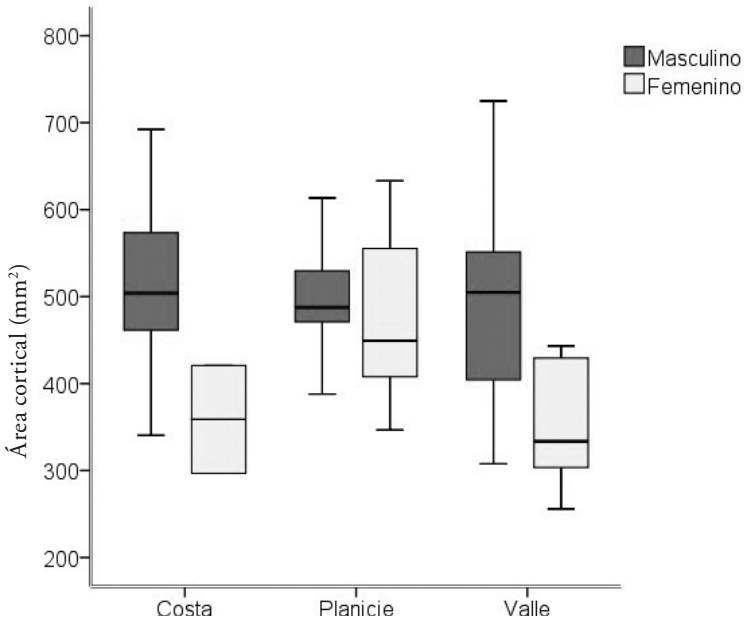
Solo se presentaron diferencias significativas en el área cortical y en el momento de inercia sobre el plano (y) o plano AP, con ligeras diferencias en los valores absolutos entre los de la costa yucateca y los de los valles centrales. El resto de las variables presentan nulas diferencias importantes y, por ende, similares respuestas al estrés biomecánico. Si bien el momento de inercia presenta un valor ligeramente mayor entre los de los valles que entre los de la costa, significando una mayor resistencia a la fractura en este plano.
Ahora prosigamos con la comparación de los valores medios de los parámetros biomecánicos entre mayas-yucatecos de la planicie y zapotecos de los valles centrales de Oaxaca, masculinos y femeninos. Los resultados para masculinos se muestran en el Cuadro 11, Figura 8.
Cuadro 11 Comparación de los valores medios de los parámetros biomecánicos; Mayas, Planicie (N= 44) y Zapotecos de los valles centrales de Oaxaca (N=27) Masculinos
| Planicie | Valle | ||||||
|---|---|---|---|---|---|---|---|
| Variable | n | m | s | n | m | s | p (t)* |
| AT | 44 | 703.54 | 93.40 | 27 | 663.10 | 134.09 | |
| AC | 44 | 508.56 | 64.66 | 27 | 490.94 | 105.20 | |
| AM | 44 | 194.98 | 36.03 | 27 | 172.16 | 37.65 | 0.013 |
| Ix | 44 | 22635.67 | 5980.55 | 27 | 21251.66 | 9239.93 | 0.016 |
| Iy | 44 | 20939.19 | 6474.74 | 27 | 19699.13 | 8432.69 | 0.063 |
| Imax | 44 | 24297.58 | 6736.23 | 27 | 22979.41 | 9869.90 | 0.054 |
| Imin | 44 | 19292.65 | 5069.75 | 27 | 17971.37 | 7595.26 | 0.012 |
| J | 44 | 43590.23 | 11397.68 | 27 | 40950.78 | 17195.19 | 0.027 |
| Ix/Iy | 44 | 1.11 | 0.22 | 27 | 1.09 | 0.21 | |
| Imax/Imin | 44 | 1.26 | 0.17 | 27 | 1.28 | 0.20 | |
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; S=Desv. Tipica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.
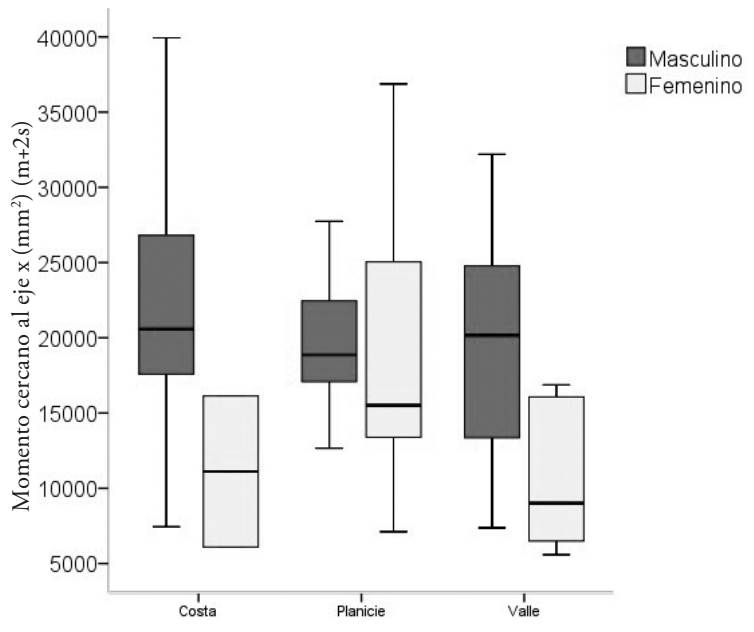
Figura 8 Momento cercano al eje x. Masculinos y femeninos de la costa y planicie de Yucatán, y de Monte Alban.
Las mayores diferencias se presentaron en los primeros y segundos momentos de inercia, validados por las pruebas t y F respectivamente. Los mayas siguen repuntando con valores altos de at, ac, am, Ix, Iy, Imax, Imin, y J. Los mayas masculinos de la planicie yucateca presentan mayores valores en resistencia a la fractura, con mayores reforzamientos en los planos x e y, tanto en los cercanos al eje como en los máximos y mínimos. No hubo diferencias en la movilidad, ambos grupos presentan valores elevados del índice de Nordin. Esto demuestra una mayor actividad física entre los mayas masculinos de las planicies yucatecas que entre los zapotecas de los valles centrales, a pesar de que son más robustos los zapotecos de los valles centrales.
En resumen, podemos afirmar, al comparar los grupos de variables de los primeros y segundos momentos de inercia, que existen más diferencias entre los masculinos que vivieron en la planicie yucateca y los zapotecos de los valles centrales y menos diferencia con los mayas de la costa, pero ambos tienen valores elevados en los PMI y en los SMI, despuntando el valor de la torca que está estrechamente relacionado con los procesos de carga física. ¿Acaso estos valores elevados de los parámetros biomecánicos podrían deberse a las exigencias en el trabajo físico que se dieron entre mayas y zapotecos al final del Clásico y principios del Posclásico, más que al entorno geográfico? En el primer caso sabemos (Arias 2012) que, durante el Clásico, el nivel de exigencia física aumentó debido al incremento en la construcción de obras civiles (lapidaria, caminos, estructuras habitacionales, etcétera) y que bien podría ser éste el caso entre los zapotecos, con ligeras diferencias determinadas por la intensidad, tasa y ritmo de los estresores biomecánicos en el fémur. Sin embargo, en los masculinos hubo ganancia de peso debido al consumo del maíz, propio de grupos dedicados a la agricultura. La respuesta de los fémures a la movilidad en ambos grupos fue similar, no presentan diferencias importantes.
Y en el caso de los femeninos, con las reservas del caso, se muestran en el Cuadro 12, Figura 9 las diferencias significativas entre la mayoría de las variables, que comprenden a los PMI y a los SMI, con excepción del índice de movilidad. Como en el caso masculino, los femeninos presentan valores mayores entre los individuos de la planicie que entre los de los valles centrales. De nueva cuenta llaman la atención los resultados de los parámetros biomecánicos entre los femeninos que, al igual que entre los masculinos, presentan valores elevados. Despunta el valor de la torca, la cual fue elevada en ambos sexos.
Cuadro 12 Comparación de los valores medios de los parámetros biomecánicos; Mayas, Planicie (N=10) y Zapotecos de los valles centrales de Oaxaca (N=10) Femeninos
| Planicie | Valle | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | n | m | s | n | m | s | p (t)* | p(F)* |
| AT | 10 | 653.94 | 136.39 | 10 | 469.50 | 93.93 | 0.002 | |
| AC | 10 | 474.86 | 96.94 | 10 | 353.88 | 66.53 | 0.004 | |
| AM | 10 | 179.08 | 43.07 | 10 | 115.62 | 30.98 | 0.001 | |
| Ix | 10 | 19661.86 | 7182.87 | 10 | 10509.05 | 3605.83 | 0.002 | 0.06 |
| Iy | 10 | 18682.52 | 8811.52 | 10 | 10440.04 | 4400.54 | 0.016 | 0.084 |
| Imax | 10 | 21302.21 | 8282.52 | 10 | 11789.32 | 4323.22 | 0.005 | |
| Imin | 10 | 17048.37 | 7443.93 | 10 | 9159.77 | 3458.72 | 0.007 | 0.032 |
| J | 10 | 38350.58 | 15508.66 | 10 | 20949.09 | 7687.87 | 0.005 | 0.069 |
| Ix/Iy | 10 | 1.13 | 0.32 | 10 | 1.05 | 0.25 | ||
| Imax/Imax | 10 | 1.31 | 0.30 | 10 | 1.30 | 0.18 | ||
AT en mm2; AC en mm2; AM en mm2; Iy en mm4; Imax en mm4; Imin en mm4; J en mm4; n= submuestra; Ix/Iy, Imax/Imin=índices de movilidad; S=Desv. Tipica
* diferencias significativas, p> 0.05
Fuente: Datos propios 2020.

Figura 9 Momento polar de inercia o Torca. Masculinos y femeninos de la costa y planicie de Yucatán, y de Monte Albán.
Esta congruencia de similitudes no es casual, pensamos que tanto hombres como mujeres estaban sometidos a exigencias de trabajo con intensidades diferentes, pero que al final presentan las mismas respuestas biomecánicas en espacios geográficos diferentes. La movilidad entre mujeres zapotecas es similar a las de las mujeres de la planicie yucateca y es muy diferente a lo encontrado por otros autores (Macintosh et al. 2014; Stock et al. 2016) que señalan que las respuestas mecánicas entre hombres son más sensibles a entornos sociopolíticos y las de las mujeres a los entornos geográficos; aquí son similares. El entorno sociopolítico es determinante en la respuesta ósea al estrés biomecánico entre las mujeres mayas y zapotecas.
Presentamos los Cuadros 13 y 14 con el propósito de hacer una síntesis de los resultados hasta aquí obtenidos de las probabilidades (t, F) de los valores medios entre grupos, según geografía y sexo e independiente del tiempo, tanto a nivel intragrupal como intergrupal y para mostrarlo más claramente.
Cuadro 13 Resumen de las probabilidades de significancia de los parámetros biomecánicos; Según geografía y sexo, independiente de la cronología
| Zapotecos | Mayas | ||
|---|---|---|---|
| Variables | Masculinos/ Femeninos | Masculinos/ Femeninos | Masculinos/ Femeninos |
| PMI | Monte Alban/ Mendrugo | Costa | Planicie |
| AT | 0.02** | 0.02** | |
| AC | 0.04** | 0.01** | |
| AM | 0.01** | ||
| SMI | |||
| Ix | 0.01** | 0.03** | |
| Iy | 0.04** | ||
| Imax | 0.06** | 0.03** | |
| Imin | 0.03** | ||
| J | 0.03** | ||
| Movilidad | |||
| Ix/Iy | |||
| Imax/Imin | [0.06**] | ||
PMI=Primeros momentos de inercia. SMI=Segundos momentos de inercia.
Sin corchetes, probabilidad de la T de Student. Con corchetes, F de Fisher o varianza.
** Altamente significativa p<0.05
Fuente: Datos propios 2020.
Cuadro 14 Resumen de las probabilidades de significancia de los parámetros biomecánicos; Según geografía y sexo, independiente de la cronología; Mayas y Zapotecos de los Valles Centrales, variación intragrupal
| Variables | Masculinos | Femeninos | Masculinos | Femeninos |
|---|---|---|---|---|
| PMI | Costa/Valle | Costa/Valle | Planicie/Valle | Planicie/Valle |
| AT | [0.03**] | 0.00** | ||
| AC | [0.09**] | [0.00**] | 0.00** | |
| AM | 0.05** | 0.01** | 0.00** | |
| SMI | ||||
| Ix | [0.00**] | 0.01** | 0.00** [0.06**] | |
| Iy | 0.06** | 0.01** [0.08**] | ||
| Imax | 0.05** | 0.00** | ||
| Imin | 0.01** | 0.00**[0.03**] | ||
| J | 0.02** | 0.00** [0.06**] | ||
| Movilidad | ||||
| Ix/Ix | 0.02** | |||
| Imax/Imin |
PMI=Primeros momentos de inercia. SMI=Segundos momentos de inercia.
Sin corchetes, probabilidad de la T de Student. Con corchetes, F de Fisher o varianza. ** Altamente significativa p<0.05
Fuente: Datos propios 2020.
La variación intragrupal de los parámetros biomecánicos entre los zapotecos, masculinos y femeninos, presentó diferencias altamente significativas en los primeros momentos de inercia y en los segundos momentos de inercia solo en el plano (y), cercano al eje central y al máximo. Sin embargo, en los índices de movilidad no hubo diferencias importantes, son homogéneos.
Es interesante que, entre los mayas de la costa, masculinos y femeninos, fueron mayores las diferencias en PMI y SMI y de la robustez, siendo mayores los valores en masculinos que en femeninos. Los valores son congruentes con los de los zapotecos, propios de sociedades agrícolas donde el maíz es el alimento primordial, amén de otros productos provenientes del ecosistema marino.
En el Cuadro 14 se muestra que los mayas de la costa y los zapotecos de los valles centrales presentan menos probabilidades significativas en los PMI y SMI, con excepción de los femeninos de la costa y de los valles en el ac y en el plano (y=Ix) y un valor mayor del am en masculinos de la costa. Lo anterior significa un mayor espesor de la cortical y una mayor resistencia a la fractura entre los mayas de la costa. El índice de movilidad cercano al eje presentó diferencias entre masculinos de la costa y entre los de valles centrales, lo cual significa una mayor movilidad entre los de la costa. No siendo así para las mujeres.
Para el caso de los mayas que vivieron en la planicie y los zapotecos de los valles centrales, vemos que existen diferencias significativas en la mayoría de las variables, tanto en los PMI (Figura 8) y SMI (Figura 9), la excepción fue en el índice de movilidad que no presentó diferencias. Ambos grupos, independiente del sexo, tuvieron una gran movilidad, a pesar de tipificarlas como sociedades sedentarias agrícolas, aquí se invocan causales proximales como el transporte de mercancías, agua, materiales de construcción, etcétera, y causales finales como el tipo de organización social basado en la centralización y segmentación de la sociedad prehispánica yucateca en los tiempos clásicos donde se requería mucha mano de obra, quizá durante varias generaciones. Al respecto Abrams y Bolland (1999) plantean las maneras de analizar el trabajo y la organización requerida para la construcción de las pirámides, desde la perspectiva de la energética arquitectónica. Calculan el costo aproximado de cada edificio en términos de trabajo humano, medido en días-hombres en el área maya. Lo anterior, y sin ser conclusiones de ellos, pero sí de nosotros, pudo haber tenido un costo biológico en las poblaciones que podemos observar en las respuestas biomecánicas corporales de hombres y mujeres.
Contrastamos nuestros resultados con los obtenidos entre indígenas americanos del sudoeste americano, Cazadores-Recolectores (CR) y Horticultores (Ho) (Bridges et al. 2000), donde se han encontrado respuestas similares. La intensa carga de trabajo asociada con actividades de subsistencias puede explicar muchas de las diferencias en las propiedades de la sección transversal del fémur; en estos últimos y durante la transición se ha hallado que la resistencia de los huesos largos se incrementa tanto en hombres como en mujeres.
Por otro lado, Ruff et al. (1999) han observado que aquellos grupos que viven en terrenos montañosos y escabrosos, independiente de las estrategias de subsistencias, tienen los miembros inferiores más fuertes que aquellos grupos que viven en terrenos menos escabrosos (por ejemplo, regiones costeras y planicies). Se argumenta que, mientras la forma del fémur puede corresponder a actividades de subsistencias, el relieve del terreno podría influir en su tamaño y robustez (Ruff 2000). Pearson (2000) encuentra que los huesos largos de las poblaciones que viven en ambientes fríos son generalmente más robustos que aquellas poblaciones que viven en climas calientes, por esta razón, él señala que los CR tienden a ser más robustos que los AG en climas fríos, mientras que un patrón inverso se observa en climas calientes.
Al investigar las diferencias en la forma a la mitad del fémur, su robustez, dimorfismo sexual y medidas externas entre grupos prehistóricos e históricos de Norteamérica, con diferentes estrategias de subsistencias y niveles de movilidad, Wescott (2004, 2006, 2008) encuentra que las variaciones significativas no son consistentes con la estructura del fémur en hombres y mujeres; sin embargo, el dimorfismo sexual sí es consistente con una gran movilidad, y añade que los masculinos tienden a mostrar modificaciones en el fémur según sus prácticas de subsistencia, mientras que la forma del fémur en las mujeres está más asociada a la región geográfica. En general, plantea un patrón general de variación intraespecífica, en donde la variabilidad morfológica a nivel geográfico y entre grupos está más asociada con diferencias inducidas por el clima, que aquellas debidas al relieve del terreno.
Contrasta con lo que nosotros hemos encontrado entre los mayas, donde el relieve del terreno no es significativo para inducir modificaciones importantes en la morfología ósea; entonces la modificación ósea sí puede referirse hipotéticamente a factores provenientes del entorno sociopolítico, más que al clima o al relieve topográfico.
Por último, al estudiar la transición de una dieta de CR a una agrícola, durante el Neolítico en Çatalhöyük o Catal Huyuk (Turquía) (7100-5950 a.P), Spencer et al. (2019) encuentran cambios importantes en la calidad y estilos de vida, como prueba del costo biológico. Este estudio revela costos crecientes para los miembros del asentamiento, incluida la exposición elevada a enfermedades y demandas laborales en respuesta a la dependencia y producción comunitaria de carbohidratos de plantas domesticadas, población en crecimiento en tamaño y densidad que provocó una fertilidad elevada y tensiones crecientes debido a una mayor carga de trabajo y a una mayor movilidad requerida para pastoreo de caprinos.
Conclusiones
Con las reservas del caso, y tomando como base el número total de fémures, así como su estado de conservación que en general fue bueno, podemos proponer algunas líneas que dibujan nuestras conclusiones. Una de las primeras premisas al realizar esta investigación fue pensar que la situación geográfica jugó un papel sumamente importante en los dos grupos de estudio, como ya ha sido señalado por la literatura internacional, pues provienen de diferentes entornos naturales, donde variables como la topografía del terreno pudieran influir en la respuesta biomecánica de los individuos. De origen, la situación geográfica juega un papel sumamente importante en estos dos grupos prehispánicos, debido al esfuerzo realizado en terrenos escabrosos por parte de Zapotecos, por ejemplo, si bien se aprecia que la topografía de Monte Albán estaba conformada por pendientes muy inclinadas en un terreno modificado intencionalmente. Aun así, es muy contrastante con la planicie o costa de la península de Yucatán, donde no existen mayores elevaciones que la sierrita de Ticul al sur y que colinda con el estado de Campeche, con poca elevación y pendientes moderadas; lo que podría marcar una diferencia física reflejada en una mayor resistencia ósea por parte de este grupo y que no pudimos apreciar en este análisis.
Es necesario señalar que la movilidad y actividad es mayor en hombres zapotecas que en mujeres, posiblemente por el gran nivel de exigencia en trabajo físico que implicaba la construcción de esta gran urbe edificada en la parte alta de un conjunto de cerros cuya altura máxima alcanza 400 m sobre el nivel del valle, así como la obtención de recursos naturales bajo forma de explotación agrícola intensiva y el agua de los ríos, la cual era transportada cuesta arriba en vasijas desde el río Atoyac en el piso del valle. Además, cabe aclarar que los fémures de los zapotecos, al igual que el de los mayas aquí analizados provienen de individuos de estatus social bajo, por tanto, cabríamos suponer que tuvieron una mayor exigencia en la construcción de Monte Albán, y de igual forma entre los mayas de Yucatán, a diferencia de los que habitaron en templos y residencias de elite y, que a final de cuentas, también se desplazaban para realizar sus actividades, pero no con la misma intensidad desarrollada por la mayoría de la población, pues no tenían más alternativa.
Los resultados expuestos nos llevan a reformular esto: más que el medioambiente, es necesario buscar otra explicación que podría venir del entorno sociopolítico o de otra condición que influyó en la estructura corporal y, particularmente, en los valores biomecánicos. Consideramos se produjo una sinergia en forma de Cadenas de Causalidades Biomecánicas en una constante interacción bidireccional y de retroalimentación positiva que moduló y determinó la forma y tamaño de los fémures en ambos grupos.
De aquí que concluyamos tentativamente, a reserva de analizar e incorporar los segmentos óseos superiores y aumentar el tamaño de nuestras muestras de ambos sexos, sobretodo de femeninos, que esta homogeneidad biomecánica encontrada tal vez no se exprese de igual manera en las extremidades superiores. Es posible que en este segmento se reflejen con mayor precisión las diferencias por sexo, sobre todo en la muestra de la costa yucateca y que, por tanto, sean las exigencias de la estratificación y desigualdad social las que estén impactando al sistema óseo, en este caso los fémures.
Por consiguiente, el ver que las dos muestras se comportan de manera similar en cuanto a los datos biomecánicos referidos, nos lleva a pensar en un sistema de producción variable y común que permeaba en una gran parte la organización social mesoamericana. Por último, como estudio siguiente y para consolidar nuestra hipótesis, proponemos, además de aumentar el tamaño de nuestras muestras, agregar a las variables biológicas y del paisaje geográfico ya consideradas, las Pendientes y Coeficientes de fricción, del Análisis del Vecino más Próximo para hacer la delimitación territorial y los Polígonos de Voronoi y Superficies de Costo de Desplazamiento.

