











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
En México, las distancias geograficas y las formas en que se han integrado culturalmente los grupos humanos han dado lugar a relativos aislamientos geneticos al irse configurando, uniendo, relacionando o alejando, de forma que desde tiempos prehispanicos y desde las nuevas formaciones surgidas durante el virreinato, se fueron entretejiendo las actuales poblaciones. En los espacios geográficos y periodos en que se involucraron, los conjuntos poblacionales han variado, por lo que deben considerarse en funcion de sus respectivos contextos dados los costos de diversa indole que involucran las relaciones humanas.
Con esto en mente, nos propusimos analizar si la distribución geográfica genética corresponde a la que alude la idea de Mesoamérica, a partir de 3 191 muestras provenientes del norte, centro sur y sureste de México. Se utilizaron marcadores genéticos del ADN mitocondrial (ADNmt), los cuales permiten rastrear aspectos sobre los asentamientos que hubo en el pasado, así como los aislamientos y el flujo genético en las distintas regiones. Dichos procesos poblacionales afectaron de forma distinta a las poblaciones en cada lugar, por lo que no es posible precisar de la misma manera en todas las regiones cómo cambio la estructura demografica (cuantitativa y cualitativamente) despues de los muchos y complejos cambios tanto en tiempos prehispánicos como durante el Virreinato, incluidas las poblaciones en las zonas que fueron refugio para los ascendientes de los actuales hablantes de lenguas indigenas. Sin embargo, el seguimiento del linaje materno expresa eventos historicos y demograficos al conservar las variantes en el ADNmt que se han ido plasmando a través del tiempo en el recorrido geográfico de la humanidad, desde la separacion de una secuencia ancestral de África hasta llegar al continente americano; de manera que es posible determinar el linaje que comparten las agrupaciones humanas a nivel continental, regional, de subpoblaciones y de familias. Se trata de un sistema donde no se produce la recombinacion genética1 y sus variaciones pasan de una generacion a otra en bloque, por lo que resulta muy útil en la reconstruccion de filogenias.
ADN mitocondrial
Respecto a la naturaleza del ADN mitocondrial diremos que, como lo indica su nombre, se encuentra en las mitocondrias; éstas son organelos de la célula que la proveen de oxigeno y varian en cantidad (de cientos a miles) segun el tipo de tejido de que se trate. Es un ADN que se distingue del nuclear al ser un genoma circular de 16 569 pb2 de doble cadena de 37 genes (Anderson et al. 1981); la secuencia completa y su organizacion se describieron a partir de un individuo europeo, estableciéndose la secuencia de referencia conocida como “Cambridge”.3 Los estudios remontan su antigüedad a los 180 000 años (Brown 1980) y la separación del continente africano del resto del mundo entre 125 500 y 165 500 años (Cann, Stoneking y Wilson 1987; Vigilant et al. 1991; Chen et al. 1995). El alto grado de correlacion del ADNmt con la distribucion geográfica posibilita conocer o entender patrones de migracion y estructuras poblacionales, aunque ello signifique conocer solo parte de la historia de las poblaciones humanas, ya que se restringen a la herencia a través de las mujeres. Así, este sistema genetico es susceptible a los efectos de la deriva genetica al reducirse el tamaño de la poblacion debido a los cambios aleatorios en las frecuencias haplotipicas4 de una generacion a otra, lo que acelera la diferenciacion entre poblaciones. En los patrones de herencia de los linajes maternos, la diversidad genetica se expresa en funcion del comportamiento migratorio de las portadoras pues, segun los lugares, causas y periodos historicos, se han movido unos grupos mas que otros. En general, la literatura reporta que a lo largo de la historia lo han hecho mas los varones y a mayores distancias, como exploradores, comerciantes y soldados, a diferencia de las mujeres quienes se han movido a pequeña escala según los intercambios entre las distintas agrupaciones de acuerdo a cada cultura.
La nomenclatura de los haplogrupos mitocondriales se denominan con letras mayusculas de la A a la Z en un orden segun su descubrimiento: L corresponde al macro-haplogrupo ancestral africano, en tanto que A, B, C y D corresponden al continente americano (Torroni et al. 1993). Posteriormente se comprobo que C y D son derivados del haplogrupo M, mientras que A y B lo son de N, siendo M y N macro-haplogrupos que se desprendieron del L3 africano hacia el Cercano Oriente y Europa hace entre 55 000 y 90 000 años. Con los marcadores mitocondriales de los haplogrupos A, B, C y D y sus respectivos haplotipos se han realizado estimaciones respecto al momento en que entraron las primeras migraciones al continente americano, entre 11 000 y 40 000 años (Volodko et al. 2008; Achilli et al. 2008), y 15 000 y 20 000 respecto al linaje paterno a partir del ADN del cromosoma Y (Seielstad et al. 2003; Zegura et al. 2004; Kitchen et al. 2008).
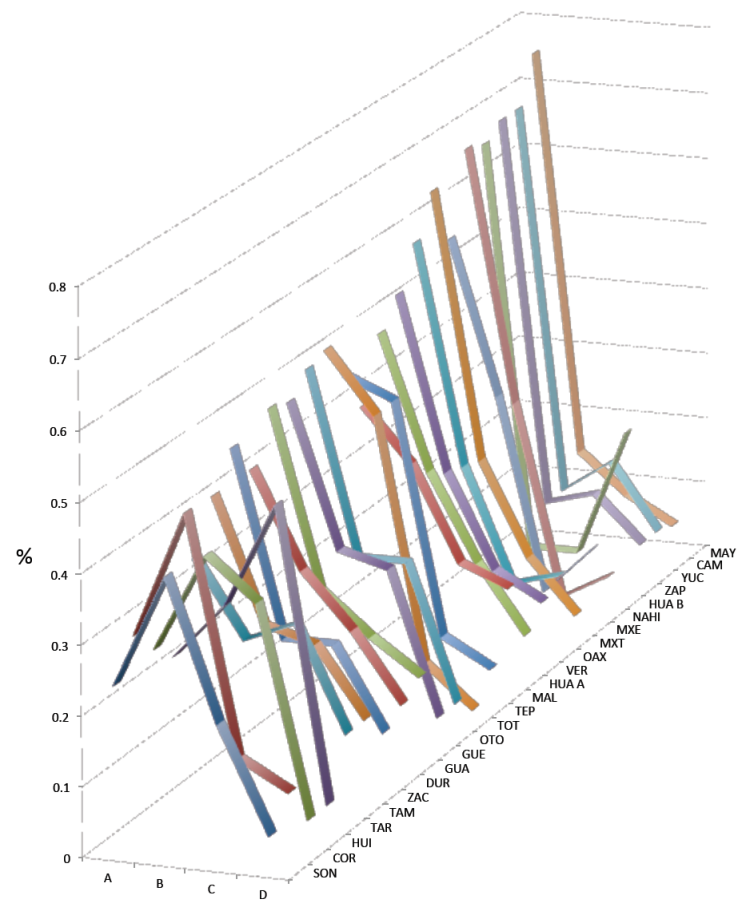
Las distancias geneticas y los procesos sociales tienen que ver con la separación de grupos tanto espacialmente como por la confrontación constante en funcion de intereses territoriales y recursos naturales y humanos. Formaciones poblacionales antiguas junto con sus propios patrones de cambio y dispersion fueron interrumpidos -unos más que otros- con la conquista española, apabullante en muchos aspectos pero no en cuanto a la herencia del linaje materno americano se refiere, el cual ha predominado hasta la actualidad en México. De la variedad de haplogrupos del ADN mitocondrial que hay en el mundo, encontramos 65% (González 2016: 65); de éste, 97% corresponde a fundadores americanos (50% A; 24% B; 16% C; 7% D); 1,8% europeos (haplogrupos R, HV, K, T, U, V, X) y 1,2% africanos (haplogrupo L) (Figura 2).

Figura 1 Los grupos de estudio. Las poblaciones urbanas se presentan en círculos grises, y las de poblaciones hablantes de lenguas indígenas en colores. El número de círculos corresponde al número de localidades.

Material y Métodos
Se analizaron 3 191 muestras contemporáneas de 16 estados de la República, incluyendo habitantes de 11 ciudades (1806 muestras) y 137 localidades de hablantes de 12 grupos hablantes de lenguas indígenas (1385 muestras) (Figura 1, Cuadro 1).5
| Grupos | Localidades | N |
|---|---|---|
| Huastecos (a) | San Luis Potosí y Veracruz: El Carrizal, Tampamolón Corona; Francisco Villa; San Vicente Tancuayalab; San Antonio Huitzquilico, Xilitla; Tatacuatla, Huehuetla; Oktzen, Tancanhuitz; Sagrada Familia, Tanquián de Escobedo; San Nicolás, Tanlajá; San Pedro de las Anonas, Aquismón; Tampacoy, Tamuín; Tampacoy, Tamuín; Unión Guadalupe, Aquismón. Chinampa de Gorostiza (cabecera municipal); Las Cruces, Chontla; La Esperanza, Tantoyuca; La Garrapata, Tantoyuca; El Mamey, Tantoyuca; El Sabino, Tantoyuca; San Francisco, Chontla Tancoco (cabecera municipal). | 36 |
| Huastecos (b) | San Luis Potosí: San Pedro de las Anonas, y Veracruz: el Mamey San Gabriel (Muñóz 2017). | 24 |
| Nahuas | Veracruz: Ixhuatlancillo | 69 |
| Zapotecos | Oaxaca: San Miguel Aloapam | 173 |
| Tepehuanes del Sur | Durango: La Guajolota, Mezquital | 220 |
| Mayas | Campeche: Calkiní | 226 |
| Mixtecos | Oaxaca: San Juan Mixtepec | 168 |
| Maltrata | Veracruz | 97 |
| Mixes | Oaxaca (Regiones Media, Alta y Baja): Asuncion, Rancho Tejas, San Cristobal Chichicaxtepec, San Francisco Jayacaxtepec, San Isidro Guayapan, San Jose El Paraiso, San Juan Bosco Chuxnaban, San Juan Cotzocon, San Juan Guichicovi, San Juan Mazatlan, San Juan Metaltepec, San Juan Otzolotepec, San Lucas Camotlan, San Miguel Quetzaltepec, San Pedro Ayasextepec, San Pedro Ocotepec, San Pedro y San Pablo Ayutla, Santiago Atitlan, Santiago Ixcuintepec, Santiago Zacatepec, Sta. Ma. Alotepec, Sta. Ma. Mixistlan, Sta. Ma. Nativitas Coatlan, Sta. Ma. Ouxmetacan, Sta. Ma. Puxmetacan, Sta. Ma. Matamoros, Sta. Ma. Tepantlali, Sta. Ma. Tiltepec, Sta. Ma. Tlahuitoltepec, Sto. Domingo Tepuxtepec, Tlaxcaltepec. | 88 |
| Oaxaca | Oaxaca | 183 |
| Mérida | Yucatán | 166 |
| Coras | Nayarit: Mesa del Nayar, Los Pinitos, Las Huertitas, El Colomo, Arroyo el Capomo, Jazmin del Manguito, Zoquipilla, Las Sillas, Arroyo del Fraile, Paso Cuate, Las Guayabas, Los Bules, Gavilanes, Los Zorrillos, San Diego del Naranjo, El Chalate Limon, Mesa del Canare, Dolores Nuevo, Jesus Maria, Las Ventanas, Pajaritos, El Saladito, El Sabino, Camarones. | 111 |
| Jalapa | Veracruz | 171 |
| Mayas | Campeche: Kalkiní | 182 |
| Chilpancingo | Guerrero | 178 |
| Guanajuato | Guanajuato | 174 |
| Durango | Durango | 161 |
| Zacatecas | Zacatecas | 170 |
| Hermosillo | Sonora | 164 |
| Totonacos | Puebla: San Juan Ozelonacaxtla y Caxhuacán | 30 |
| Otomíes | Hidalgo: Actopan, Chicuautla, Huichapan, Tezontepec de Aldama, Mixquiahuala, Tetepango, Tlahuelilpan, Atitalaquia, Tlaxcoapan, Atotonilco, Tepeji del Río, Tula de Allende, Alfajayucan, Tepetitlán, Chapantongo, Nopala, Tecozautla, San Salvador, Santiago de Anaya, El Cardonal, Ixmiquilpan, Tasquillo, Tepatepec (Juárez 2014). | 94 |
| Tamaulipas | Cd. Victoria | 160 |
| Huicholes | Nayarit: Gpe. Ocotan, Sta. Barbara, El Saucito, La Mojarra, Cienega Sta. Rosa, Sta. Rosa, Caliguey, Zapote, El Maguey, Trigalillo, Gavilanes, San Juan Peyotan, Atonalisco, El Gabino, La Guerra. | 79 |
| Total 24 | 137 | 3191 |
En el presente estudio, exponemos los resultados obtenidos con la técnica de PCR (reacción en cadena de la polimerasa) y cortes de fragmentos de ADN con las enzimas de restricción Hae, Hinc y Alu para el caso de los haplogrupos A, C y D respectivamente, y la deleción de 9 pb en la región intergenica COII y ARN-Lys para el haplogrupo B. Para la visualización de las bandas de ADN se utilizaron geles de poliacrilamida. En cuanto a los análisis de diversidad, los índices de diferenciación genética entre poblaciones se estimaron con las distancias Fst, estadístico F que Wright y Malécot desarrollaron para el estudio de distribución a partir de análisis de varianzas (Holsinger y Weir 2009). Fst estima la diferenciación por la reducción de la heterocigosis e indica la falta de flujo génico. Cuando es igual a cero no hay diferencia, y cuando es igual a uno, significa que las poblaciones son completamente diferentes; la estructura poblacional resulta de la diferencia entre las frecuencias de las subpoblaciones debido a las separaciones a lo largo del tiempo en relación con el tamaño efectivo de la población y el número de generaciones. Excoffier y colaboradores introdujeron AMOVA, donde el promedio cuadrado es una medida de la distancia evolutiva entre haplotipos. Para nuestro estudio se estimó la distancia Phist (prácticamente igual que Fst en el caso del estudio de haplogrupos)6 con el programa ARLEQUIN 3.0 (Excoffier 2005). Con la matriz de diferenciación de Phist, se generó un dendrograma de agrupamientos entre las poblaciones con el programa STATA/MP 14.2.
Mesoamérica
Puesto que en el pasado las poblaciones humanas tenian menos posibilidades de moverse rapida y masivamente, la division de los grupos estaba mas marcada en cuanto a la diversidad genetica -determinada por la densidad demografica, los aislamientos poblacionales y las distancias espaciales- y dependia de la cantidad y tipo de gente que se movia a distintas velocidades segun los itinerarios y/o asentamientos que las necesidades de cada region.
Seguramente hubo cambios notables en las frecuencias genéticas en los diferentes sitios en las fechas de que hay constancia respecto al desarrollo de la protoagricultura estimada entre los 9 000 y 6 000 años atrás (Cutler y Whitaker 1967; Flannery 1985; MacNeish y Eubanks 2000; Niederberger 1979; Whitaker, Cutler y MacNeish 1957). Pero, independientemente de cuanto y como aumento el numero de individuos y su asociación con la agricultura, es un hecho que hubo expansiones y desplazamientos entre los diferentes asentamientos, los cuales dieron lugar al flujo de genes y constantes cambios en las poblaciones; al parecer las redes de intercambio (de productos, ideas, personas, idiomas, etcetera) desde tiempos muy antiguos favorecieron estos procesos en las regiones centro-sur de Mexico.
Si vamos a hablar de una frontera mesoamericana, tenemos que considerar que los desplazamientos de la gente dependen de la accesibilidad en función de distancias geográficas, el tiempo de los recorridos, la cantidad de personas que se desplazan, los avances tecnológicos para salvar las distancias en cada época y las posibilidades de que se dé el intercambio genético (pues moverse y migrar -las personas, sus cosas, sus tradiciones, su lengua- no implica necesariamente la reproducción de los que se trasladan). Todo esto significa que podríamos encontrar una gran cantidad de formas de arreglos en los subconjuntos poblacionales genéticos entre los mesoamericanos, entre los mesoamericanos y los no mesoamericanos, y entre los no mesoamericanos, de manera que “lo” mesoamericano no llegara a perfilarse. Específicamente, respecto a la herencia y la temporalidad de la Mesoamérica prehispánica, no se sabe si la distribución de la variabilidad de una determinada población en un periodo dado conformaba una población conjunta de distintos orígenes, si convivía como grupo cultural pero separada en estratos (de distinto origen),7 o si su diversidad era semejante a otras regiones, sitios o poblaciones. Este punto hace -en tanto no se tengan muestreos genéticos amplios de restos arqueológicos- imposible definir una Mesoamérica en términos de herencia biológica. Sin embargo, podría suponerse que antes de los eventos posthispánicos, el flujo de genes estuvo condicionado, en principio, por las distancias y las dificultades geográficas respecto al comercio y las relaciones sociales. Habría que evaluar en qué medida en la Mesoamérica del Postclásico -que involucró grandes movimientos poblacionales cambiaron las estructuras genéticas del Clásico, y en qué espacios (mismos que para momentos de la conquista influyeron en las composiciones subsecuentes).
El desarrollo de los pueblos mesoamericanos, a decir de López y López ([1996] 2010), favorecido por la situación geográfica, produjo desplazamientos poblacionales hacia el norte, sur y ambas costas del centro del país, de manera que, a la llegada de los españoles, los mexicas tenían el dominio del Altiplano Central, abastecido con los productos de tributarios en un flujo de comunicación constante con las zonas que daban al océano Pacífico y las del Golfo de México. Las relaciones comerciales se daban de Michoacán a la península de Yucatán (García 2006), aunque para Fournier (2006), la compleja red de intercambio entre regiones hacia y desde Mesoamérica fue más de orden cultural, involucrándose solamente un sector minoritario de las poblaciones participantes en un intenso intercambio entre los distintos pueblos mesoamericanos.
Como concepto, Mesoamérica ha estado en debate desde hace ya varios lustros, sobre el cual nos referiremos dadas sus implicaciones espaciales para la comprensión de muchos aspectos de las agrupaciones genéticas. Los mesoamericanistas han propuesto teorías y metodologías distintas, por lo que su construcción ha dependido de intereses diversos, ideologías y momentos históricos particulares de distintos especialistas. En principio, la idea tiene que ver con un espacio y los grupos que en éste o su periferia se desarrollaron, y que como área cultural sirvió a la política populista del gobierno cardenista para unificar las culturas prehispánicas [de una buena parte] del territorio actual y como ideal de unidad nacional (Rodríguez 2000: 52-53apudJáuregui 2008). En la compilación de la obra del etnólogo alemán Konrad Theodor Preuss, Jáuregui y Neurath (1998: 34) recuperaron la prefiguración del concepto de Mesoamérica hecha por Preuss que posteriormente desarrollaría Kirchhoff en 1943, y al que Preuss se refiere como “círculo cultural mexicano” pero no como “un área con fronteras fijas y zonas de influencia claramente perfiladas [sino como...] una región poco definida donde se presentan ciertas relaciones culturales concretas que siempre tienen algo que ver con los antiguos mexicanos” (Preuss [1911] 1998: 404, 405). Retomaremos a Diez (2007) para señalar que en la teoría de las áreas culturales, los procesos de transformación se producen mediante la expansión de objetos, técnicas e ideas de zonas nucleares a otras más lejanas, y ello conforma la base del difusionismo de la transmisión cultural de la escuela de Viena, que a su vez es el exponente etnográfico de las corrientes filosóficas del idealismo alemán y sus seguidores estadounidenses de la escuela historicista de la primera mitad del siglo XX. En la arqueología fue determinante esta corriente, de manera que se utilizaron instrumentos analíticos espaciales y recursos cartográficos relacionados con la clasificación tipológica, distribución de objetos y rasgos, para explicar modelos expansivos de aculturación, identificación de áreas culturales, así como su evolución en el tiempo y en el espacio. Tenemos entonces la descripción de Kirchhoff ([1943] 1992) de Mesoamérica como una superárea cultural de cultivadores superiores con elementos característicos, con expansiones y retracciones en el espacio en las distintas épocas, unos más exclusivos que otros según su distribución en otras áreas. Los límites van, respecto al sur, más o menos desde la desembocadura del río Montagua hasta el golfo de Nicoya pasando por el lago de Nicaragua; y respecto al norte desde el río Pánuco hasta el Sinaloa, pasando por el Lerma, frontera poco precisa. La definición alude a la situación prehispánica al momento de la conquista, cuyo objeto fue aplicar un método histórico y lingüístico e identificar rasgos culturales (Kirchhoff ([1935, 1943] 1992: 6, 28-45). En su modelo, los movimientos migratorios quedan confinados al quedar las culturas de “cultivadores superiores” enmarcadas dentro de ciertos límites geográficos y ecológicos cuando se enfrentan a grupos cazadores recolectores y semiagricultores -en su mayor parte localizados en Aridoamérica y Oasis América- cuyo territorio desértico y árido abarca el noroeste, norte y noreste del México actual y suroeste de Estados Unidos, y cuya frontera en la parte meridional ha variado a través de los siglos (a la altura del río Pánuco al Sinaloa, pasando por el Lerma). Esta definición alude a la situación prehispánica al momento de la conquista. Según vemos en López Austin y López Luján ([1996] 2000: 15-17), en la superárea cultural de Mesoamérica existieron
[…] grupos humanos ligados por un conjunto complejo y heterogéneo de relaciones [… que] a lo largo de los milenios se establecieron entre sociedades que vivían en áreas contiguas [y] el resultado fueron tradiciones o historias compartidas. Fundamentalmente, las relaciones se generaron a partir de los intercambios constantes de bienes, de los desplazamientos transitorios o permanentes de grupos dentro de la superárea, de los intereses compartidos entre las élites que gobernaban las diferentes entidades políticas, del dominio de unas sociedades sobre otras, de las acciones bélicas tanto de alianza como de conflicto, etc. Más que como un conjunto de elementos inmutables en el tiempo y en el espacio, las tradiciones que caracterizaron la superárea cultural deben concebirse como una particular corriente de concepciones y prácticas en continua evolución multisecular y con notables particularidades regionales. Las sociedades de una misma superárea cultural podían diferir en nivel de desarrollo. Lo importante fue que las relaciones se constituyeron en forma estructural y permanente. En cambio, las meras relaciones comerciales o las simples copias de estilos artísticos entre las tres superáreas no bastaron para integrar a sus pueblos en una misma tradición. Por ejemplo, el intercambio entre Oasisamérica y Mesoamérica, aunque intenso, no uniformó los fundamentos socioculturales de ambas superáreas.
Con un enfoque distinto, López Aguilar (2000) señala que el enunciado teórico difusionista “Mesoamérica”, una vez definido como escenario geográfico -con fronteras y pertenencias o no pertenencias- se fue redefiniendo con agregados de evidencia empírica a favor, cargado de presupuestos teóricos provenientes de diferentes perspectivas, como si se tratara de una realidad de facto (por ejemplo véase la alusión que hace a Chapman (1990: 23) cuando ésta habla de una Mesoamérica uniforme que, aunque con diferentes trayectorias, “mantiene una identidad común”); el mismo López Aguilar encuentra otras inconsistencias entre las que mencionamos la ambigüedad de que no estén establecidas las condiciones necesarias y suficientes8 para que determinada cultura pertenezca al ámbito mesoamericano (porque Kirchhoff advierte que un elemento para determinada superárea no implica que se encuentre en todas las áreas que la componen).
Al considerar estos argumentos como referente para pasar a los datos obtenidos sobre la herencia biológica, no dejaremos de tomar en cuenta las palabras de Leroi-Gourhan ([1973] 1989: 271, 272, 301) quien dice que
[…] si nos ceñimos a la historia de cada uno de los distintos elementos de un grupo étnico (el tipo físico, la lengua, las tradiciones religiosas y las técnicas), nos percataremos de que están dotados de muy distintos ritmos y son divergentes, de manera que existe una discontinuidad en las representaciones sucesivas de su historia general. Con excepción de contados [casos] los conjuntos casi nunca son absolutamente concordantes. Sin embargo, ello no implica la discontinuidad de la evolución de los diversos elementos […]
Discusión y Resultados
Con lo expuesto, tomando en cuenta que la distribución y densidad poblacional representan eventos demográficos en correlación con las interacciones socioculturales entre y dentro de los distintos grupos sociales, y que dicha distribución se refleja en la estructura genética de las poblaciones, daremos cuenta de la distribución de los linajes del ADNmt que conciernen a los fundadores americanos y el área conocida como Mesoamérica. Es un hecho que la historia biológica difiere de la historia social por infinidad de factores intrínsecos a la naturaleza de cada uno de estos procesos, muchos de éstos determinados, a su vez, por los eventos culturales en tanto que dan lugar a intercambios o aislamientos genéticos que generan una distribución diferencial en los distintos espacios. La conformación genética de que daremos cuenta, nos remite no sólo a eventos presentes sino (como ya se mencionó) a los de un pasado relativamente reciente como sería el Virreinato, la época prehispánica y tiempos más remotos desde el poblamiento de América.
En principio, los resultados mostraron que las poblaciones, o grupos de ellas, en México se dan en un continuum de flujo en todo el pais, mayor entre las poblaciones urbanas como era de esperarse, con diferencias -cuando las hubo- a nivel de las grandes regiones (norte, centro, sur y sureste), y diferencias, también, dentro de las mismas (Figura 3).
En la Figura 4 se muestran los componentes principales de los 4 haplogrupos distribuidos entre los grupos de estudio. El elemento más grande de variación se encuentra en los haplogrupos A y C. El factor 1 explica 53% de la varianza, mientras que el factor 2, 26% (80.31% en total).

Figura 4 Componentes principales de los haplogrupos A, B, C, D del ADNmt en las muestras de estudio. Los 4 haplogrupos se distribuyen en cuadrantes.
En las Figuras 5 a 7 se desglosan las poblaciones según la similitud en las frecuencias de los distintos haplogrupos. La diferencia más notoria se da en el conjunto del noroeste conformado por Sonora, coras, huicholes y tarahumaras al presentar menor frecuencia en el haplogrupo A y mayor en B, separándose a su vez tarahumaras y huicholes por una mayor frecuencia en C (Figuras 3, 4 y 5). En las Figuras 4 y 6 se observan tepehuanes y Maltrata con una mayor frecuencia de A y B. El resto presentan semejanza de B y C: Tamaulipas (aunque con menos A), Zacatecas y Durango, Guanajuato y Guerrero, así como otomíes y totonacos donde aumenta ligeramente tanto B como C. En las Figuras 4 y 7 los grupos presentan el haplogrupo A en aumento, no mucho B, poco C y D; a este conjunto corresponden huastecos(a), Veracruz, Oaxaca y mixtecos, y mixes (en éstos abunda un poco más el haplogrupo A, y nahuas y huastecos(b) que se caracterizan por tener más B dentro de este conjunto. Finalmente, en las Figuras 4 y 8 se observa el conjunto mayas-Yucatán-Campeche con el mayor porcentaje de A, junto con los zapotecos donde, además, aumenta el D.

Figura 5 Frecuencias del ADNmt americano en grupos del noroeste. El conjunto noroeste conformado por Sonora, coras, huicholes y tarahumaras presentan menor frecuencia en el haplogrupo A y mayor en B, y se separa a su vez tarahumaras y huicholes por una mayor frecuencia en C.

Figura 6 Frecuencias del ADNmt americano en grupos del norte centro y este. Los tepehuanes y Maltrata presentan una mayor frecuencia de A y B. El resto se asemejan en los haplogrupos B y C: Zacatecas y Durango, Guanajuato y Guerrero, otomíes y totonacos donde aumenta ligeramente tanto B como C. Tamaulipas presenta menos A.

Figura 7 Frecuencias del ADNmt americano en grupos del centro este y sur. Los grupos del centro-este y sur presentan el haplogrupo A en aumento, no mucho B, poco C y D; huastecos(a), Veracruz, Oaxaca y mixtecos; en los mixes abunda un poco más el haplogrupo A y nahuas-huastecos(b) se caracterizan por tener más B.

Figura 8 Frecuencias del ADNmt americano en grupos del sureste. En el conjunto mayas-Yucatán-Campeche se presenta el mayor porcentaje de A, junto con los zapotecos donde, además, aumenta el D.
En la Figura 10 se muestran las cercanías genéticas de los distintos grupos representados por círculos, indicando por colores los que son semejantes, y con líneas grises se indica la división y subdivisiones de Mesoamérica. Los zapotecos parecen relacionarse más con el sureste que con mixtecos, mixes y Oaxaca; los dos grupos huastecos se alejan entre sí acercándose los huastecos(a) a Veracruz y Oaxaca, en tanto los huastecos(b), a nahuas de Ixhuatlancillo; los tepehuanes se asemejan a Maltrata; se relacionan las poblaciones urbanas de Durango, Zacatecas, Guanajuato y Guerrero; en tanto que los coras y huicholes en vez de acercarse entre sí, se acercan a Sonora los primeros y a los tarahumaras los segundos.

Figura 9 Semejanzas genéticas y distribución geográfica en los grupos de estudio. Agrupamientos por distancias genéticas Phist. Se observan dos grandes ramas en un rango entre 0 y 0.19. El sureste (Yucatán, Campeche y mayas) junto con zapotecos se separa del resto de los grupos, éstos se subdividen. En general, se presenta asociación entre las distancias genéticas y las geográficas pero las subdivisiones y distancias concretas no necesariamente corresponden a las de Mesoamérica.

Figura 10 Semejanzas genéticas y distribución geográfica en los grupos de estudio. Los grupos se representan en círculos. Los mismos colores indican cercanía genética. La división y subdivisiones de Mesoamérica se indican con líneas grises.
Como se ve, no hay un patrón que coincida completamente con los límites mesoamericanos, pues se presentan (o no) distancias dentro de la propia Mesoamérica. Esto no implica la ausencia de concordancia entre las grandes distancias geográficas y las genéticas -el conjunto de la gran región norteña se aleja de los grupos del sureste. Sería prudente, entonces, hablar de regiones y poblaciones o grupos, explicando el contexto cultural y demográfico específicos en sus respectivos tiempos y espacios.
Para López Austin (1989), los procesos que involucran creencias, espacios geográficos y expresiones de la cultura material, no son ajenos o excluyentes entre sí, y la mesoamericana correspondería a una unidad religiosa en el tiempo y en el espacio que protege valores, creencias, prácticas y representaciones fundamentales con importantes diferencias regionales y temporales. A manera de conjunto estructurado de un microsistema conceptual, dicha unidad -que el autor llama núcleo duro- engloba a todos los demás sistemas (sociales), los ordena y los ubica. Dichas representaciones destacan la geometría del universo, la oposición de contrarios que segmenta el cosmos para explicar su diversidad, orden y movimiento, y la división dual armonizada en un estado de equilibrio (ibidem: 11). Explorando cuánto de esta persistencia cultural conlleva procesos sociales directamente relacionados con eventos que inciden en la reproducción y, por lo tanto, en la conformación genética de los pueblos, cabría pensar que: a) la agricultura -como eje que ha condicionado tanto las creencias como modos de vida y subsistencia “mesoamericanos”- fue un factor determinante en los aspectos demográficos y, con ellos, en la conformación genética de sus poblaciones, o b) que en términos de genética poblacional no haya (ni hubo) una correspondencia con “Mesoamérica”. En ésta, resulta inexacto respecto al análisis de genética poblacional, precisar la cantidad de individuos que hubo en el pasado, los flujos migratorios de los diversos grupos en los distintos momentos, así como las contracciones y expansiones poblacionales, pues tanto el flujo como la deriva genéticos tienen que ver con muchos “límites” que no son los mesoamericanos o que se presentan o no dentro de la misma Mesoamérica. Los resultados, entonces, deben verse como un complejo de eventos: la presencia similar de frecuencias de los 4 haplogrupos del ADNmt en todos los grupos de las distintas regiones de México hace pensar en establecimientos humanos desde tiempos muy antiguos en el territorio, pues con todo y la intensidad de los cambios demográficos posthispánicos, ha permanecido. Por otro lado, la agricultura intensiva y el Estado implicaron contracciones poblacionales que generaron la diferencia en la distribución de la diversidad genética. Sea mucho o poco lo diferentes que fueron respecto al pasado, la distribución genética actual tiene que ver con el establecimiento de vínculos de dependencia y tributación post-hispánicos entre españoles y las culturas agrícolas, caracterizadas por estar densamente pobladas y contar con estructuras sociopolíticas fuertemente jerarquizadas, a diferencia de las norteñas cuyos parámetros socioeconómicos y culturales no permitieron dichas relaciones. Éstos eran cazadores, recolectores, pescadores, semiagricultores y agricultores que se dispersaban según el aprovechamiento de recursos y las temporadas de caza y recolección, sin las estructuras políticas centralizadas del centro-sur y sur de México (como la mexica). Así, cuando se dio la conquista española se dieron grandes acomodos poblacionales, muchos radicales, pero también gran parte estuvieron relacionados con los acomodos previos, al tiempo que se hicieron y rehicieron otros, en una lucha desigual que repercutio en la sobrevivencia y la reproduccion de personas y grupos.
En la Figura 9 se presentan los agrupamientos de las distancias genéticas Phist. La división en dos grandes ramas se da en un rango entre 0 y 0.19. El sureste (Yucatán, Campeche y mayas) junto con zapotecos se separa del resto de los grupos. Éstos se subdividen y puede observarse una asociación entre las distancias genéticas y las geográficas grosso modo (sureste, centro, norte), pero las subdivisiones y distancias concretas no necesariamente corresponden a las de Mesoamérica.
A manera de comentarios finales, podemos señalar que:
no se puede aseverar que la actual distribución genética nativa americana de los mexicanos corresponda a la que hubo en el pasado prehispánico,
siempre debe tenerse en cuenta que hubo un gran cuello de botella después de la Conquista que disminuyó (y cambió) la distribución de la variabilidad dada la gran mortandad que redujo drásticamente el número de nativos (en el Altiplano Central se calculó entre 4 y 25 millones de personas según los “minimalistas” o “maximalistas” respectivamente (Borah y Cook 1971; Gerhard 1986; Rosenblat [1935] 1954; Mendizábal 1946; Kubler 1942) con una recuperación demográfica hacia 1620-1650 (con excepciones locales y grandes retrocesos durante las epidemias de 1692, 1697 y 1736-1739), y cambios irreversibles en las estructuras poblacionales,
los numerosos y masivos desplazamientos pusieron a varios grupos en contacto, muchos de los cuales estaban separados o eran enemigos previamente (Ouweneel y Hoekstra 1998; Blanton et al. 1996; Cramaussel 2004),
hubo una extinción generalizada de lenguas (Moctezuma 1991: 127; Reff 1991) y/o grupos enteros, pero los hablantes de dichas lenguas al parecer se asimilaron a grupos hablantes de otras lenguas y su herencia genética ahora es parte de grupos a los que sus antecesores no pertenecían.
muchos eventos se combinaron de diferente modo según la densidad poblacional, el aislamiento y las formas de interacción entre las poblaciones de las distintas grandes regiones del país,
no se puede hablar de los mismos patrones de desarrollo demográfico en todo el territorio dadas las diferencias en el interior de los grupos -tanto de hablantes de lenguas indígenas como de los nuevos grupos que se fueron generando entre los grupos y sus estratos sociales-. Tales diferencias dependieron de las limitaciones geográficas para el despliegue económico, las dinámicas demográficas, la cultura y sus cambios en los diversos grupos indígenas también cambiantes, así como del dominio territorial de los conquistadores.
Aunque pareciera que todos estos eventos impiden percibir si hubo o no una frontera genética mesoamericana -a causa del cúmulo de tantos acontecimientos y la imposibilidad de hacer precisiones respecto a cuáles, cuántos y en qué momento ocurrieron- el dato genético no deja de reflejar el origen de los grupos humanos y sus desplazamientos. Efectivamente se observan diferencias a nivel de grandes regiones, pero las distancias biológicas tienen más que ver con la densidad poblacional y ciertas formas de dinamismo que con la separación geográfica. La forma de lidiar de cada grupo con las adversidades frente a conquistas, formas de control, distribución de recursos (materiales y humanos) o preferencias en las formas de interactuar (confrontar, comerciar, someterse, etc.) han sido innumerables en todas partes, y de ello han resultado las disminuciones poblacionales, los aislamientos, los maridajes entre poblaciones distantes, las prohibiciones entre poblaciones vecinas, etc. Por supuesto las distancias geográficas y las dificultades que cada población ha tenido que enfrentar tanto antes como después de la conquista española han determinado parte de lo que ahora se presenta en las distribuciones genéticas, por lo que la interpretación de éstas es mucho más complejo que la concordancia directa lengua - etnia - cultura material, lo mesoamericano o lo que excluye.
Por ello, es necesario no sólo ampliar en cantidad y calidad los estudios de ADN mitocondrial (y por supuesto de otros marcadores genéticos) tanto de materiales antiguos como contemporáneos (estos últimos próximos a homogeneizarse como consecuencia de la multiplicación de migraciones actualmente), sino acotar y ampliar los estudios etno-históricos particulares de cada región y sus poblaciones de manera interdisciplinaria.

