











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introducción
Desde mediados del siglo pasado, el estudio de múltiples fenómenos de interés para la biología se ha visto enriquecido por el creciente éxito que ha tenido el enfoque de los sistemas complejos (Martínez-Mekler 2000; Martínez-Mekler y Cocho 1999; Flores y Mekler 2011; Cocho 2017), donde el comportamiento de sistemas multifactoriales de muchos componentes fuertemente enlazados, con dinámicas y correlaciones a varias escalas de resolución espacial y temporal, da lugar a la emergencia de propiedades globales. En biología lo anterior es la norma. La disponibilidad de una cantidad de información previamente insospechada a varios niveles de resolución requiere de los desarrollos de herramientas de análisis de datos, modelación matemática y simulación, utilizadas para los sistemas complejos, muchas de ellas provenientes del estudio de los sistemas dinámicos y de la física estadística. Con este enfoque se busca incorporar ensayos experimentales puntuales, a manera de ir colectando piezas que eventualmente constituirán un “rompecabezas” completo que describa apropiadamente algún proceso biológico de interés. Siempre ha quedado manifiesta la importancia y necesidad de pasar de colectar a armar dichas piezas de conocimiento empírico, para darles un sentido integrativo en el contexto de un sistema completo. Este es el propósito de la biología de sistemas.
En este contexto ha sido de interés el estudio transdisciplinario de redes formadas por biomoléculas y sus interacciones, las cuales subyacen en la regulación de una variedad de procesos moleculares que sostienen el funcionamiento de la célula, como, por ejemplo, redes genéticas, metabólicas, de señalización bioquímica, entre otras. Las investigaciones se han centrado en caracterizar tres propiedades fundamentales de las redes: estructura, dinámica y función. El trabajo de esta índole pertenece a lo que se ha denominado biología cuantitativa, la cual plantea la deducción de principios cuantitativos que puedan dar lugar a la construcción de modelos predictivos de fenómenos biológicos.
En este artículo se abordan cuestiones pertinentes a la fecundación, principalmente el estudio de la fertilización externa del erizo de mar, con miras a entender los mecanismos que permiten a los espermatozoides localizar el óvulo. Dicha capacidad está regulada por una red de señalización que genera fluctuaciones de concentración de calcio en los flagelos, la cual es disparada por compuestos provenientes del entorno del huevo. Se busca un entendimiento de los procesos biomoleculares involucrados en el funcionamiento de esta red, esto mediante un modelo dinámico discreto de redes lógicas. Una descripción de algunos de los avances obtenidos bajo este programa de investigación se encuentra en el capítulo “Modelaje de la vía de señalización de calcio relacionada con la motilidad de espermatozoides” de la XVIII Escuela de Verano en Física (Espinal et al. 2011).
El enfoque de la biología de sistemas pone además de manifiesto propiedades globales de la dinámica de la red de señalización tales como su estabilidad, robustez, redundancia y criticalidad. Aquí se muestra que la dinámica se encuentra en un régimen crítico que comparte propiedades con los fenómenos críticos estudiados en la física y que ocurren en los sistemas dinámicos. El estudio abre la posibilidad de comentar sobre la complejidad en un sentido más amplio.
En la próxima sección se presentan las bases fisiológicas de nuestro objeto de estudio para pasar, en la sección 3, a la construcción de modelos en redes y mostrar en la sección 4, la criticalidad del sistema. La sección 5 presenta una discusión sobre aspectos generales de criticalidad en varios contextos. En la sección 6 se comentan rasgos encaminados a tipificar la complejidad en un sentido amplio, sin por ello dejar de hacer referencia al estudio de la fecundación. La última sección recapitula sobre la situación presente del estudio de fenómenos complejos con miras al futuro.
2. Fertilización del erizo de mar como caso de estudio
Uno de los mecanismos adaptativos esenciales que caracterizan y ayudan a la subsistencia de gran parte de los organismos es la capacidad de reaccionar ante un estímulo externo, ya sea acercándose hacia o alejándose de este, en donde el beneficio de dicho desplazamiento puede tener diferentes significados de acuerdo con el contexto, como lo puede ser la búsqueda de comida, la relocalización de algún tipo de células con funciones reparadoras hacia un sitio de lesión, o el encuentro con otra célula para reproducirse con ella. La respuesta sensomotriz que permite a una célula desplazarse con un movimiento dirigido hacia (o lejos de) la fuente de un gradiente de moléculas es conocida como quimiotaxis. El erizo de mar es un organismo que ha sido empleado por más de un siglo como referencia para el estudio de la quimiotaxis. Entender los elementos de la fisiología de sus espermatozoides relacionados con su motilidad sigue siendo un reto. Este tipo particular de célula se vale de una estructura subcelular conocida como flagelo para desplazarse describiendo trayectorias helicoidales en el medio. Dichas trayectorias son moduladas por moléculas de un polipéptido pequeño conocido como speract, las cuales son liberadas desde el recubrimiento del óvulo, difundiéndose para formar un gradiente alrededor de este último, y sirviendo como una señal de localización que el espermatozoide debe interpretar. La unión de dichas moléculas con receptores específicos localizados en el flagelo del espermatozoide desencadena una serie de reacciones bioquímicas y electroquímicas al interior de este (Figura. 1) que, en conjunto, generan fluctuaciones en la concentración intraflagelar de iones calcio ([Ca2+]i), las cuales constituyen una señal capaz de inducir cambios directamente en la curvatura del flagelo y por consecuencia en el nado. De esta forma, el fenómeno de quimiotaxis se puede entender como el resultado de la interacción coordinada entre variables externas e internas, relacionadas, respectivamente, con los gradientes de la concentración de estímulo explorados por la trayectoria del espermatozoide y el conjunto de eventos moleculares que suceden al interior del flagelo disparados por el estímulo externo (Darszon et al. 2008).

Nota: Las moléculas participantes se encuentran en el flagelo del espermatozoide e involucran principalmente receptores específicos de speract, enzimas (GC, AC) que sintetizan segundos mensajeros (cGMP, cAMP), canales iónicos y transportadores que permiten el flujo regulado de potasio (K+), sodio (Na+), cloro (Cl-) y calcio (Ca2+), los cuales, a su vez, generan cambios en el potencial de membrana (v) y pH intracelular. La tira gris representa la membrana celular con canales e intercambiadores iónicos señalados.
Fuente: Espinal-Enríquez et al. (2017: 4) .
Figura 1 Red de señalización activada por speract.
3. Modelación mediante redes lógicas discretas
Dada la naturaleza no lineal de la mayor parte de las interacciones que integran la red de señalización y la acumulación gradual de mediciones experimentales sobre propiedades puntuales relacionadas con los eventos moleculares que subyacen a la quimiotaxis, ha sido necesario recurrir a modelos teóricos para avanzar significativamente en la construcción de un panorama completo que integre las observaciones empíricas en una imagen coherente y que al mismo tiempo proporcione una idea clara de cúales son las propiedades esenciales del sistema de señalización visto como un todo.
En las referencias (Espinal-Enríquez et al. 2011, 2014 y 2017) desarrollamos modelos discretos, lógicos, con el propósito de ayudar a entender la emergencia de oscilaciones de calcio, por medio de su vía de señalización, al regular la motilidad del espermatozoide y llevar a cabo predicciones que han sido comprobadas experimentalmente por nuestro grupo. El estudio permite esclarecer el papel específico que desempeñan ciertos nodos en el contexto de la red completa. Este tipo de modelos son una extensión del formalismo booleano propuesto por primera vez por Stuart Kauffman (1969) para estudiar la dinámica de redes de regulación genética. En la propuesta inicial de Kauffman, mediante una aproximación de grano grueso, los niveles de expresión de cada uno de los genes eran representados por nodos binarios, mientras que las interacciones entre genes lo eran mediante enlaces entre nodos. Este formalismo implica la discretización tanto de los valores que los nodos pueden adquirir como de las unidades de tiempo en que suceden los cambios de estado. Si bien el formalismo de Kauffman no fue desarrollado para algún sistema biológico en particular sino que fue planteado como un modelo generalizable, ha resultado ser útil a la hora de adaptarlo a casos específicos en la biología, tales como la diferenciación celular en el desarrollo floral (Mendoza et al. 1998), diferenciación celular del sistema inmune (Martínez Sánchez et al. 2015) y el ciclo celular de la levadura (Davidichi y Bornholdt 2013), entre otros. Una presentación detallada, pedagógica al respecto se encuentra en (Aldana 2012).
Para la modelación más reciente de la red de señalización del espermatozoide en términos de una red lógica como las descritas en el párrafo anterior, se consideraron 20 entidades moleculares (Figura 2) representadas mediante 18 nodos binarios y 2 nodos ternarios, es decir, que puedan cambiar entre 2 o 3 estados posibles según la naturaleza fisiológica particular del nodo a ser modelado.

Nota: Las interacciones activadoras, represoras y duales están coloreadas, respectivamente, con negro, rojo y amarillo.
Fuente: Espinal-Enríquez et al. (2017: 5) .
Figura 2 Abstracción de la red de señalización bioquímica mediante una red lógica discreta.
Denotando el estado de un nodo i a un tiempo t como σi(t), la configuración a un tiempo t de la red completa queda expresada por un vector Σ(t) con longitud N = 20:
La evolución de cada nodo sucede en tiempos de máquina y el valor que adquiere al siguiente instante depende del estado actual de sus respectivos nodos reguladores. Bajo la suposición de que los tiempos característicos de transición son similares para todos los nodos, estos son actualizados síncronamente en cada paso de tiempo. Así, un nodo dado σi tendrá asociado un conjunto de k reguladores (σi1, σi2, …,σik ) y la función de actualización Fi toma como entrada la configuración de reguladores a un tiempo t:
Las funciones de actualización propias de cada nodo son construidas como tablas de verdad que contienen toda la combinatoria posible de estados de sus reguladores y un valor de salida asignado a cada una de las frases regulatorias, tomando en cuenta observaciones experimentales o hipótesis basadas en conocimiento previo cuando hay información faltante. Establecer estas reglas fue un ejercicio interdisciplinario que requirió de un intercambio teórico-experimental a fondo.
Dada una condición inicial Σ(0), la dinámica de la red completa eventualmente alcanza un patrón periódico, después de transcurrido un número finito de pasos, es decir, la configuración del sistema llega a un estado que se repite cada cierto número de pasos de tiempo. Este patrón periódico se define como atractor del sistema, y el número de pasos de tiempo entre configuraciones que se repiten define el periodo del atractor; mientras que las trayectorias dadas por la sucesión de configuraciones que preceden al atractor reciben el nombre de transitorios. El conjunto de condiciones iniciales que llegan a un atractor constituye la cuenca de atracción, y el conjunto de atractores con sus respectivas cuencas forma el paisaje de atracción del sistema. En particular, para el nodo del calcio (dCa) se obtiene un patrón oscilatorio que puede asociarse con el tren de oscilaciones típicamente observado experimentalmente en el flagelo de espermatozoides (Wood et al. 2003). La evolución de la red para una condición inicial basal se muestra en la Figura 3.
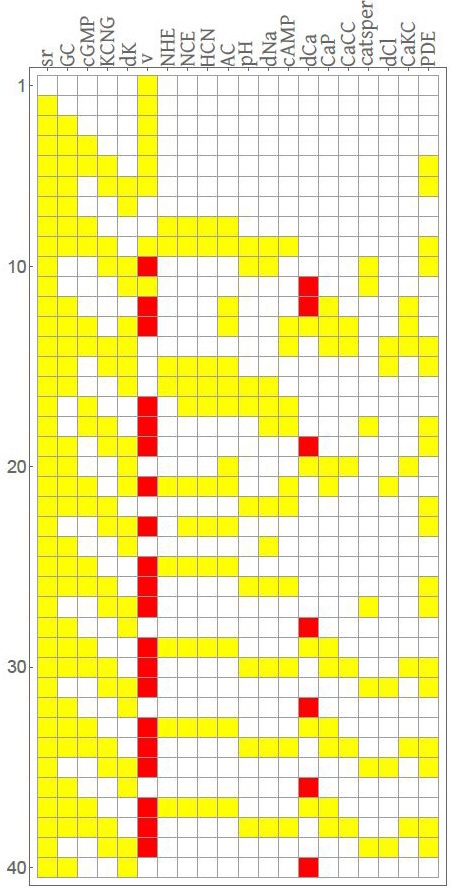
Nota: En esta gráfica, la configuración en el tiempo t = 0 corresponde al renglón superior, y los subsecuentes renglones corresponden a la evolución del sistema en cada paso de tiempo hasta llegar al atractor. Los colores blanco, amarillo y rojo corresponden, respectivamente, a los estados 0,1 y 2.
Fuente: Priego Espinosa et al. (2017: 148) .
Figura 3 Simulación para una condición inicial en la cual todos los nodos están en estado basal.
A partir de este tipo de modelos se ha logrado (Espinal-Enríquez et al. 2011, 2014 y 2017) un mejor entendimiento del funcionamiento de los canales iónicos luego de haber explorado in silico la activación o bloqueo constitutivo de algunos de ellos, seguido de su corroboración experimental. Uno de los casos más complejos explorados fue la caracterización del mecanismo de acción del ácido niflúmico (Espinal-Enríquez et al. 2014) sobre la dinámica de la señal de calcio, ya que esta droga tiene la peculiaridad de poder actuar sobre al menos 3 tipos de canales, ya sea inhibiéndolos o activándolos. En otro artículo (Espinal-Enríquez et al. 2017), gracias a la consideración de variantes de la red abordada inicialmente y el estudio por medio de simulaciones de diversos escenarios de interacción, al buscar consistencia con resultados experimentales observados, se pudo predecir que CatSper es el canal de calcio determinante. Haber hecho una predicción de esta relevancia por medio de un modelo matemático, pone de manifiesto los alcances que pueden tener estudios teóricos en la biología. El trabajo va más allá de proveer un mejor entendimiento y corroborar experimentos previos. La teoría plantea predicciones cuantitativas y cualitativas, y se convierte en un motor para el desarrollo de nuevos experimentos. Por supuesto, la palabra final la tendrán los experimentos. En este tenor, a largo plazo se espera que este tipo de estudios, con su extensión a mamíferos, pueda contribuir a la elaboración de fármacos que alteren la fertilidad ya sea aumentándola o disminuyéndola, por ejemplo, que contribuya al desarrollo de anticonceptivos masculinos.
4. Régimen dinámico crítico
El funcionamiento y la evolución de los seres vivos presupone implícitamente que la forma en que estos contienden ante cambios internos o ambientales depende de mecanismos suficientemente robustos como para que no se colapsen fácilmente y al mismo tiempo suficientemente flexibles como para codificar un repertorio de respuestas con ventajas adaptativas para diferentes escenarios. Si bien pareciera difícil imaginar un sistema en donde coexistan estas dos propiedades aparentemente opuestas: robustez y adaptabilidad, se ha visto que las redes que operan en un régimen dinámico crítico con respecto a la propagación de perturbaciones dan cabida a la optimización de ambas capacidades (Flores y Martínez-Mekler 2011; Aldana 2012).
Derrida propuso un cálculo que proporciona una medida de cómo perturbaciones sobre condiciones iniciales producen alteraciones en la configuración de una red que se propagan en el tiempo (Derrida y Pomeau 1986). Cuando t → ∞, estas perturbaciones tienen tres posibles destinos: se propagan a toda la red (régimen caótico), mueren eventualmente (régimen ordenado) o llegan a un escenario intermedio donde se constriñen a una proporción de nodos semejante al tamaño de la perturbación inicial con comportamientos estadísticos similares. A este último régimen se le ha denominado crítico (Derrida y Pomeau 1986). El origen del término proviene de los fenómenos críticos estudiados en la física, asociados con transiciones de fase en las que se presentan correlaciones a todas escalas. La siguiente sección presenta una discusión sobre criticalidad.
Dada una configuración inicial Σ(0), definimos una perturbación de tamaño h como aquella que genera una nueva configuración Σ’(0) que difiere de la original en el valor de h nodos, decimos entonces que la nueva configuración está a una distancia de Hamming h de la incial. Al normalizar esta perturbación por el tamaño de la red (H = h/N), podemos tener una medida que expresa el porcentaje en que fue alterada una configuración dada.
El mapeo de Derrida M(H(t)) = 〈H(t + 1)〉, que relaciona cómo cambian en promedio las alteraciones inducidas por una perturbación H realizada al tiempo t después un paso de evolución, puede indicarnos el régimen dinámico en el cual opera una red discreta. En la práctica, para calcular numéricamente este mapeo, se muestrea un conjunto de condiciones iniciales del espacio de configuraciones sobre las cuales se hacen incidir perturbaciones que abarcan todos los tamaños posibles, deja evolucionar a los pares Σ(0) y Σ’(0) resultantes y se mide el tamaño promedio de la perturbación luego de ese paso de evolución. Es importante enfatizar que se trata de una propiedad promedio de la evolución de todo el sistema y no de la de una condición inicial particular.
Para tiempo continuo la susceptibilidad de una red lógica discreta está definida por la pendiente al origen del mapeo:
donde S determina el régimen dinámico en que opera la red: ordenado (S < 1), desordenado (S > 1) o crítico (S ≈ 1). Para tiempos discretos normalizados:
Al calcular el mapeo de Derrida para nuestra red, resulta que el valor de la suceptibilidad es muy cercana a 1, lo cual indica que su dinámica opera en el régimen crítico (Figura 4a).

Nota: Con propósitos de referencia, se grafica una recta identidad con un patrón de colores rojo y azul. (4a) Red completa. Obsérvese que el valor de susceptibilidad S es cercano a 1, por lo cual esta red opera en el régimen crítico. (4b) Redes resultantes de knock-outs individuales. Entre paréntesis se encuentran los correspondientes valores de susceptibilidad. Nótese que, al igual que la red completa, las curvas se aproximan a la vecindad del origen con una pendiente cercana a 1, con excepción de dos casos: el calcio (dCa) y el voltaje (v), cuyas curvas son menos tangentes a la recta identidad y tienden hacia el régimen ordenado.
Fuente: Priego Espinosa et al. (2017: 151) .
Figura 4 Mapeos de Derrida calculados sobre 500,000 condiciones iniciales aleatorias.
En el cálculo anterior, la criticalidad está relacionada con la invariancia a lo largo del tiempo de la representación de las alteraciones inducidas por una perturbación en condiciones iniciales. Cabe preguntarse qué tan robusto es este comportamiento y qué tan sensible es a cambios en la estructura y definición de la red. A nivel estructural, consideremos la respuesta del comportamiento de la red ante la supresión de un nodo. Sucede que al llevar a cabo esta supresión (conocida en la literatura como knock-out de un componente de la red) el comportamiento crítico de la red resultante se preserva.
En la Figura 4b se muestra que ante la supresión de nodos (uno a la vez), se conserva la pendiente de uno en la curva de Derrida (S≈1), lo cual, recordemos, es indicativo de un régimen dinámico crítico. Por ende, hemos mostrado que la criticalidad es una propiedad robusta ante modificaciones estructurales. En el lenguaje de los sistemas dinámicos a este comportamiento se le conoce como estabilidad estructural. Para la biología esta propiedad es altamente significativa en términos evolutivos. Cabe comentar que con la eliminación de los nodos dCa y v, se obtienen las pendientes S que menos se aproximan a uno (Figura 4b), esto puede estar ligado a que son los que tienen más conexiones salientes y entrantes (Figura 4b), y que guardan un significado central desde un punto de vista fisiológico (Darszon et al. 2008). Consecuentemente, su ausencia constituye una perturbación mayor a las demás del sistema.
Dada la estrecha relación entre la vía de señalización que estudiamos y el nado del espermatozoide, los comentarios previos sobre robustez y capacidad evolutiva en régimen crítico se aplican directamente sobre el encuentro de los gametos. La criticalidad provee elementos para explicar por qué dos especies de erizo de mar que viven en distintos entornos (orillas del mar y agua profunda) mantienen la misma vía de señalización (robustez) pero manifiestan marcadas diferencias en grado de sensibilidad quimotáctica (adaptabilidad). Otra ventaja evolutiva de la criticalidad es que al optimizar transferencia de infor mación aumenta la eficiencia de la respuesta del nado al medio, lo cual es importante dado que los espermatozoides en el mar tiene una expectativa de vida corta.
5. Consideraciones sobre la criticalidad
En un inicio, el término criticalidad se acuñó en la física para el estudio de transiciones entre fases termodinámicas, posteriormente se adoptó en el contexto del comportamiento de sistemas dinámicos con transiciones entre fases dinámicas. El mapeo de Derrida, aunque se encuentra ligado a fases dinámicas, tiene características propias. La relación entre las diversas acepciones de la criticalidad aún no ha sido descifrada por completo y es tema de investigación. Las transiciones de fase han sido motivo de intenso estudio dentro de la física: transiciones hielo/líquido, líquido/gas, ferro/paramagneto, transición superfluida o superconductura son algunos de los ejemplos. Entre la variedad de las posibles transiciones de fase se distinguen los fenómenos críticos, los cuales caracterizaremos aquí en términos de la transición ferro/para-magnética. Los metales ferromagnéticos, al ser expuestos a un campo magnético generado externamente, poseen un rango de temperaturas a las cuales se magnetizan, esto es, mantienen una magnetización en ausencia del estímulo externo, se comportan como imanes. Resulta que al ir aumentando la temperatura, la magnetización decrece, hasta desaparecer a una temperatura que denotaremos por Tc; a partir de ella, al no retener magnetización inducida, el material se comporta como paramagneto. La magnetización es la densidad de momentos dipolares magnéticos, si consideramos un modelo simplificado bidimensional en que los momentos adquieren solo uno de dos valores (+/-), con interacciones de corto alcance (modelo de Ising), lo que ocurre es que debajo de Tc se tienen dominios magnéticos dominantes, ya sea positivos o negativos (uno de los dos), intercalados con zonas del signo opuesto. Cuando nos encontramos por arriba de Tc los dominios se van desintegrando y se presenta una distribución aleatoria de cúmulos muy pequeños de espines alineados que van desapareciendo al incrementarse la temperatura. En cambio, muy cerca de la temperatura crítica se tienen dominios compuestos de dominios, compuestos de dominios, en forma iterativa, dando lugar a una estructura sumamente compleja. Estas estructuras se presentan a todos tamaños, y se preservan ante cambios en la resolución de observación, es decir, hay una invariancia de escala, todo el sistema está estadísticamente correlacionado a todas las escalas (Bruce y Wallace 1989). Las interacciones son de corto alcance, pero las correlaciones se presentan en todos los rangos. Para mayor claridad, decimos que dos sitios están correlacionados si la variación de uno de ellos es perceptible a nivel estadístico por el otro. O sea, las variables no son estadísticamente independientes. La presencia de este sin fin de correlaciones a la temperatura crítica Tc da lugar a singularidades en cantidades termodinámicas en la vecindad de Tc, como, por ejemplo, la susceptibilidad magnética que registra los cambios en la magnetización ante variaciones de un campo magenético externo, esto se debe a relaciones conocidas como fluctuación-disipación. El estudio de los fenómenos críticos dio lugar al desarrollo de la teoría del grupo de renormalización, por la cual Kenneth Wilson obtuvo un Premio Nobel (Wilson 1975). Para los propósitos de esta discusión recalcamos que por debajo de Tc las estructuras se congelan, por arriba se pierden y en Tc se preservan.
Por otra parte, en los sistemas dinámicos, tales como sistemas de ecuaciones diferenciales no lineales o mapeos no lineales acoplados, que se utilizan para modelar dinámica de poblaciones ecológicas (ecuaciones de Lotka-Volterra, mapeo logístico), osciladores no lineales (ecuación de Van der Pol), dinámicas excitables (ecuaciónes de Hodgkin-Huxley y Fitzhugh-Nagumo), entre otros, presentan transiciones entre comportamientos dinámicos cualitativamente muy diferenciados que ocurren al variar algunos de sus parámetros. En sistemas dinámicos a esas transiciones se les denomina bifurcaciones. Como ejemplo de una bifurcación se tiene la transición de comportamiento regular periódico a uno caótico (dinámica determinista, con sensibilidad a condiciones iniciales, sin predictibilidad, con rasgos parecidos a los comportamientos azarosos) que se presenta a un valor “crítico” particular de su parámetro de bifurcación (May 1976). En ese valor, la dinámica es marginal, entre el orden y el caos, se presentan correlaciones temporales a todas las escalas, así como una invariancia de comportamiento ante cambios en la resolución temporal de observación. Las analogías con las transiciones de fase termodinámicas saltan a la vista; han sido y continuán siendo estudiadas formalmente.
En las redes que aquí estudiamos, la criticalidad se refiere al régimen entre dos fases dinámicas, y se cuantifica en términos de respuestas a alteraciones generadas por perturbaciones en condiciones iniciales. Para su estudio viene a la mente la terminología utilizada en el análisis de los sistemas dinámicos de una transición entre un comportamiento regular que elimina el efecto de las perturbaciones y otro caótico que las amplifica. Usualmente, en los estudios sobre dinámica caótica el sujeto primordial de análisis es el comportamiento asintótico de las trayectorias del sistema dinámico y su sensibilidad ante variaciones de condiciones iniciales. Aunque bajo ciertas condiciones, el criterio de Derrida se puede asociar con ese comportamiento de las trayectorias (Aldana 2003), su enfoque es sobre comportamientos globales, promedio, a un paso de tiempo. En este sentido, más que una transición caos-orden, parece ser más adecuada la nomenclatura de una transición expansión-contracción. Es más, bajo esta perspectiva, la proximidad a 1 de la susceptibilidad S definida con anterioridad, tiene rasgos reminiscentes de la divergencia de la susceptibilidad de compresión volumétrica en el punto crítico termodinámico.
Un asunto que merece ser recalcado es que, a diferencia de los casos como el magnético y la ecuación logística donde hay un parámetro de control (la temperatura y el parámetro de bifurcación, respectivamente) que sintoniza al sistema al punto crítico, en nuestro caso es la dinámica, per se, la que lleva al régimen crítico. Este tipo de comportamiento se conoce como “criticalidad auto-organizada” (Bak et al. 1987) y ha sido motivo de intenso estudio.
Enfatizamos que el régimen dinámico crítico de nuestra red preserva estructuras a todas las escalas tal como es el caso del magnetismo en punto crítico o la dinámica marginal de los sistemas dinámicos. Este comportamiento es indicativo de correlaciones y sensibilidad del sistema a todas escalas. Una propuesta, materia de controversia, es que en el régimen crítico la transferencia de información se optimiza. En la biología, una propiedad de esta índole tiene repercusiones profundas. Este comportamiento, aunado a la coexistencia de adaptabilidad y robustez bajo un régimen dinámico crítico, da sustento a la hipótesis de que la vida se desarrolla en un punto crítico. No es de extrañarse, por ello, que se haya manifestado en variedad de fenómenos biológicos (Muñoz 2017), en particular en diversos tipos de redes regulatorias: redes genéticas (Balleza et al. 2008; Krotov et al. 2014; Nykter et al. 2008), neuronales (Baggs y Plenz 2003; Beggs 2008; Tagliazucchi et al. 2012; Haimovici et al. 2013) y de señalización (Gupta et al. 2007; Kim et al. 2014; Nivala et al. 2012). Con nuestro trabajo mostramos que en un proceso biológico tan fundamental como lo es la fertilización, la criticalidad sale a relucir una vez más. ¿Será que algo similar ocurre en los fenómenos sociales?
6. Reflexiones sobre complejidad
En tiempos recientes el interés en el estudio y comprensión de los sistemas complejos ha ido en continuo ascenso. Aunque, comúnmente, se acepta que los procesos biológicos son per se complejos y los sociales aun más, reiteradamente se delibera sobre cuál es la definición de un sistema complejo (Thurner 2016). Más que definir, la cuestión es poder percibir un fenómeno complejo. El análisis que aquí se presenta sobre algunos aspectos de la fecundación abre la posibilidad de resaltar comportamientos que permitan distinguirlo como complejo. Cada uno de los atributos que a continuación se plantean, sin ser necesarios, ni suficientes, ayudan a caracterizar los fenómenos complejos. La sinergia que se establezca entre ellos va ligada a un aumento de complejidad.
Transdisciplinariedad. Este estudio es resultado de un intenso intercambio entre investigadores de varias disciplinas (biólogos, físicos y computólogos), tanto teóricos como experimentales. Las preguntas, los planteamientos y las pau tas a seguir son resultado de esa colaboración; se requirió del desarrollo de lenguajes y modos de pensamiento en comuún. Vale la pena puntualizar que, aunque el enfoque de los sistemas complejos proporciona una plataforma ad hoc para abordar aspectos de fondo de problemas multidisciplinarios (Cocho y Martínez-Mekler et al. 2011, 2014 y 2017), la complejidad no está supeditada a la transdisciplinaridad, existen problemas y situaciones propias de una sola disciplina que pueden dar lugar a comportamientos complejos.
Jerarquización y multiplicidad de escalas. El trabajo aquí presentado, junto con nuestras publicaciones relacionadas (Espinal-Enríquez et al. 2011, 2014 y 2017), integra distintos niveles de descripción e interacción (molecular, subcelular, celular) se abordan aspectos bioquímicos, fisiológicos, hidrodinámicos; comportamientos individuales y colectivos que se presentan a distintas escalas de resolución espacial y temporal. Eventualmente emergen comportamientos funcionales que en ocasiones pueden ligarse a procesos de auto-organización. Es importante resaltar que la riqueza de niveles de comportamientos temporales es tan importante como la presencia de estructuras espaciales a distintas escalas de descripción. La transmisión de información inter e intra nivel es un elemento crucial. Los desarrollos para poder integrar información y comportamientos de diferentes escalas es una de las características que ha despertado amplio interés en el enfoque de los sistemas complejos.
No linealidad y retroalimentación. En general, los sistemas complejos son multicomponentes con interacciones no lineales entre ellos. La no linealidad puede dar lugar a comportamientos tanto caóticos, en la práctica impredecibles, como regulares y robustos (Martínez-Mekler 1990). El que los efectos no sean proporcionales a las causas, introduce un elemento de sorpresa. Los procesos de retroalimentación son determinantes para el comportamiento de los sistemas complejos, este puede darse entre elementos de un nivel de descripción, o de distintos niveles de descripción. Es más, puede incluso presentarse con el entorno, en cuyo caso suele hablarse de adecuación. Llega incluso a suceder que, en ocasiones, cuando el número de componentes con interacciones no lineales tiende a infinito, el tiempo para llegar a un estado estable asintótico (conocido como atractor) también diverge; la dinámica siempre será transitoria, por más que se avance, nunca se llega (Livi et al. 1990).
En nuestro estudio de la fecundación las interacciones no lineales y los circuitos de retroalimentación se presentan en la vía de señalización de la concentración del Ca2+ y condicionan el nado del espermatozoide. Para el caso de mamíferos, considerar las relaciones entre diversos niveles espacio-temporales y con el entorno que se presentan a lo largo de la fecundación es determinante.
Colectividad y comportamientos emergentes. Al considerar sistemas de muchos componentes, sus interacciones y las dinámicas que presentan, es de interés de terminar, estudiar y comprender los comportamientos colectivos que surgen. En el ámbito de los estudios de complejidad se habla de comportamientos emergentes, que no pueden explicarse solo en términos de un conocimiento individual a fondo de sus componentes. La expresión de que en un sistema complejo el todo es mayor a la suma de las partes hace alusión a esta situación. En nuestro estudio podría pensarse que la quimiotaxis es una propiedad emergente que proviene de la interrelación entre la dinámica bioquímica intra-extra flagelar, y el nado por medio de motores moleculares en gradientes químicos provenientes del huevo.
Un formalismo de considerable atención recibida para el estudio de estos sistemas es el de redes. Una red consta de nodos (los elementos del sistema) y enlaces (las interacciones) entre los nodos. Los valores de los nodos a un tiempo dado definen el estado del sistema y la evolución temporal depende de las interacciones, en general no lineales. Como ya se mencionó, la red objeto de nuestro estudio corresponde a la vía de señalización del calcio en el flagelo y los enlaces están definidos por las funciones regulatorias. La situación se complica cuando se presentan procesos de retroalimentación, sean con el entorno o entre los mismos componentes, los cuales generan cambios en las funciones regulatorias. Se tiene entonces una evolución tanto de nodos como de enlaces. Para el caso de organismos marinos esta situación puede darse por los cambios en el entorno al explorar por medio del nado gradientes quimioatrayentes (por ejemplo, el speract) en tiempos conmensurables con los de la dinámica interna de los nodos. En el caso de los mamíferos, al recorrer el tracto femenino las concentraciones externas de varios estímulos cambian con su nado, lo cual modifica el comportamiento de las vías de señalización que a su vez afectan el nado, aunado a esto se tienen cambios en la viscosidad y la morfología del tracto. La concatenación de estas retroalimentaciones es un indicativo del grado de complejidad del sistema. Es aquí donde las relaciones temporales son cruciales. Si el entorno varía lentamente, la dinámica interna puede considerarlo como fijo; por otra parte, si el entorno cambia rápido en relación con los tiempos característicos internos, la consideración de un valor promedio de los estímulos es aplicable. El problema más agudo se presenta cuando los tiempos internos y los del entorno son conmesurables. En este caso no se cuenta con las condiciones para que se llegue a un estado asintótico estacionario. La dinámica del sistema corresponde a estados transitorios que se van modificando por el intercambio con el entorno, se transita entre estados transitorios. En lenguaje coloquial “por más que me acerco, nunca llego, pues todo el tiempo me andan cambiando la jugada”.1 El sistema está constreñido a encontrarse siempre fuera de equilibrio. Otro factor temporal que contribuye a la complejidad se genera si los tiempos característicos del cambio de los nodos son disímiles, la evolución temporal individual es entonces asíncrona, lo cual dificulta llegar a comportamientos globales coherentes. La determinación de condiciones que conduzcan a una sincronización global resulta fundamental.
Un desarrollo teórico reciente que contribuye a poder contender con las situaciones anteriores son las redes multicapa en las cuales los componentes del sistema pueden operar en varias redes que abordan distintos comportamientos, las cuales se encuentran interconectadas. Un mismo nodo puede participar en varias de las redes. Los desarrollos formales y aplicaciones de redes multicapa son materia de intenso estudio (Boccaletti et al. 2014; Kivela et al. 2014).
Inhomogeneidad. Buena parte de la complejidad de un sistema puede provenir de su inhomogeneidad. Bajo estas condiciones no se puede recurrir a simplificaciones de comportamientos promedio, efectivos. La homogeneidad es más patente a escalas de alta resolución. La falta de homogeneidad puede estar asociada con estructuras recurrentes bien definidas pero con frecuencia se presenta como un desorden. Los tratamientos estadísticos resultan reveladores, como es el caso de determinación de correlaciones e información mutua. Conceptos probabilísticos son requeridos y formulaciones estocásticas son recurrentes. En nuestros modelos de fecundación la resolución experimental aún no permite cuantificar la inhomogeneidad, una vez que se tengan los datos será un factor a considerar.
Estocasticidad. El ruido es un elemento recurrente en los fenómenos complejos; en fenómenos biológicos y sociológicos el entorno siempre es ruidoso. La inclusión del ruido como un factor determinante del desarrollo de los sistemas complejos que puede contribuir a su funcionalidad es un cambio paradigmático respecto a su concepción como un elemento de distorsión que debe ser eliminado. En biología, algunas estructuras y comportamientos coherentes afloran solo en presencia del ruido (Vázquez-Rodríguez et al. 2017). Algunos formalismos matemáticos que dan lugar a dinámicas con transiciones entre transitorios son estables gracias a la presencia de ruido de fondo (Muezzinoglu et al. 2010; D’Huys et al. 2016). Cabe aquí la reflexión del eminente matemático A. N. Kolmogorov: “De hecho, todo el valor etimológico de la teoría de la probabilidad se basa en que: fenómenos azarosos presentes a grandes escalas crean en su comportamiento colectivo una estricta regularidad no azarosa” (Gnedenko y Kolmogorov 1954), o dicho de otra manera: “Nada hay tan determinsita que un tambache de procesos azarosos, desordenados”.
Multifactorialidad. Esta útima característica se ha llegado a considerar definitoria. La dependencia de un proceso de múltiples causas que incluso pueden provenir de diferentes niveles de organización, aunque no cuestiona la causalidad, sí puede dar lugar a comportamientos totalmente inesperados. El avance extraordinario de capacidades de obtención y almacenamiento de información ha puesto de manifiesto una multifactorialidad/multiescala impresionante que requiere del desarrollo de metodologías de minería de datos masivos y aprendizaje profundo. La complejidad inherente a esta situación es evidente.
7. Colofón
Conceptual y metodológicamente queda claro que los sistemas dinámicos no lineales (discretos o continuos, deterministas o estocásticos) y la física estadística fuera de equilibrio son herramientas naturales para el estudio de la complejidad. El crecimiento vertiginoso de las capacidades de cómputo, almacenamiento y procesamiento de información, que permiten simulaciones cada vez más realistas, estudio de redes intrincadas, manejo y entendimiento de grandes bases de datos, provee un punto de partida para trabajos futuros. Se espera que esto, aunado a capacidades experimentales antes insospechadas y desarrollos teóricos de frontera en diversas áreas de conocimiento, permita que un entendimiento y solución de la problemática que el mundo actual nos presenta sea más accesible. Estos desarrollos son una respuesta a la transformación sociocultural que estamos viviendo, C.H. Waddington escribió en 1977 el libro Herramientas del pensamiento, el cual abrió nuevos senderos para el desarrollo del estudio de los sistemas complejos; ahora, el terreno está fértil para un libro sobre formas del pensamiento.

