











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El chile (Capsicum annuum L.), es una de las especies más cultivadas y producidas en todo el mundo (Demirkaya y Günes 2022). Entre las hortalizas es uno de los cultivos que más se consumen, y ocupa el sexto lugar en producción mundial después del tomate, la cebolla, la col, el pepino y las berenjenas. La producción a nivel mundial en 2020, fue de 40.3 toneladas métricas y México ocupó el segundo lugar en producción con más de 2.8 toneladas métricas, superado únicamente por China con 33.9 toneladas métricas (FAOSTAT 2022). México, se considera uno de los principales centros de domesticación y de mayor diversidad genética de C. annuum (Crosby 2008, Saba et al. 2020), este cultivo representa parte de la historia de este país y su aprovechamiento se remonta desde épocas prehispánicas (Cyphers et al. 2009).
El chile poblano se encuentra entre las tres variedades más cultivadas, después del jalapeño y el mirasol (SIAP-SAGARPA 2021), el fruto se consume en fresco o en seco y es muy degustado en platillos como el mole, chiles rellenos y tacos, además de elaboración de diversas variedades de salsas. Su producción ha decrecido en los últimos años, debido a la mala calidad de plántulas producidas (García et al. 2011), siendo el Damping off, una de las principales enfermedades causadas por hongos como Phytophthora capsici, Rhizoctonia solani, Fusarium spp., Pythium spp., y Alternaria spp. (Bautista-Calles et al. 2010); además se suman daños y enfermedades causadas por nemátodos, áfidos, ácaros y trips (Huerta et al. 2007, Crosby 2008). Muchos de estos problemas, se pasan a campo (como fuente de inoculo) al momento del trasplante y pueden ocasionar pérdidas considerables en la producción; según datos recopilados con los propios productores, los factores anteriores han provocado falta de interés en la continuidad del cultivo y se ha optado por producir el maíz, frijol y algunos frutales, lo que acrecienta el riesgo a la desaparición de las variedades locales (Pozo 1983).
El sistema de producción de plántulas en el cultivo de chile, depende de los agroquímicos, y su uso irracional puede provocar contaminación del suelo y contribuir al calentamiento global (Czaja et al. 2015). El uso de microorganismos nativos de la rizosfera, como los hongos micorrízicos arbusculares (HMA) y las bacterias promotoras del crecimiento vegetal (BPCV), son algunas alternativas para reducir el uso de productos químicos y producir de forma sostenible (Yadav et al. 2015, Etesami y Adl 2020). Los HMA pertenecen al phylum Glomeromycota (Fitter et al. 2011) y forman asociaciones simbióticas con más del 80% de las plantas terrestres (Smith y Reed 2008, Zhu et al. 2010), incluyendo al chile poblano (Almaraz-Suarez et al. 2021). Mejoran el crecimiento de las plantas, debido a que favorecen el traslado de nutrientes (principalmente de nitrógeno y fósforo), y agua hacia la planta, proporcionan mayor tolerancia al estrés ocasionado por factores bióticos (patógenos) y abióticos (sequia, suelos salinos, alcalinos o contaminados) (Sheng et al. 2008, Kong et al. 2020).
Las BPCV han sido clasificadas como biofertilizantes, fitoestimuladores y biopesticidas (Somers et al. 2004). Primero, por incrementar la disponibilidad de nutrientes para las plantas, entre ellos nitrógeno y fósforo; segundo, por promover el crecimiento de las plantas mediante la producción de fitohormonas, principalmente de auxinas y giberelinas; y finalmente, por el control de enfermedades mediante la producción de antibióticos y metabolitos antifúngicos (Sandhya et al. 2010, Ahemad y Kibret 2014, Dahiya et al. 2019, Ali et al. 2020).
Aplicar los HMA y las BPCV, en la producción de plántulas para el trasplante, podrían traer beneficios significativos en el crecimiento y por ende mayor calidad, lo que implicaría mayor probabilidad de pervivencia en campo. Por lo anterior, el presente trabajo, planteo como objetivo evaluar el efecto de la inoculación de HMA y BPCV, en la eficiencia fotoquímica del fotosistema II y en el crecimiento de plántulas de chile poblano.
Materiales y métodos
Establecimiento del experimento
El experimento se realizó en el invernadero del laboratorio de Microbiología de suelos, dentro de las instalaciones del Colegio de Postgraduados Campus Montecillo, Texcoco, Estado de México, con coordenadas geográficas 19° 30’ LN y 98° 53’ LO, a 2 220 msnm.
Se utilizaron semillas de chile poblano adquiridas con productores del estado de Puebla. Se emplearon semilleros de 200 cavidades (Marca AgroPlastiK®, Modelo CH-200), El sustrato que se utilizó en el experimento, fue una mezcla de Peat moss (Premier®), agrolita (Agrolita®) y vermiculita (Agrolita®) (relación 1:1:1). La mezcla fue esterilizada en autoclave a 20 libras de presión, durante tres horas diarias y un día de descanso, en total 9 horas, con la finalidad de eliminar microorganismos contaminantes como patógenos.
Antes de la siembra en semilleros, las semillas de chile poblano fueron desinfectadas durante tres minutos en agua con cloro al 1% y lavadas tres veces con agua destilada estéril. Las semillas fueron sembradas directamente en los semilleros; éstos fueron regados cada tercer día con agua destilada y cada 20 días con solución nutritiva Long Ashton, agregando baja cantidad de fosforo (11 mg L−1) para no afectar la colonización micorrízica (Hewitt 1966).
Material microbiológico y preparación de inóculos
Se utilizaron especies de HMA y una cepa bacteriana, colectadas en agroecosistemas de chile poblano, en el municipio de San Matías Tlalancaleca, estado de Puebla. Las especies de HMA fueron Funneliformis geosporum + Claroideoglomus spp. (FGC) y Claroideoglomus etunicatum (CE). Los HMA fueron propagados en cultivos trampa durante seis meses con pasto Ray-grass (Lolium hybridum) cultivado bajo el mismo sustrato del experimento; se utilizaron aproximadamente 7 (± 1.0) g de sustrato como inoculo para rellenar los semilleros, antes de la siembra, que contenían 140 (± 20) y 128 (± 15) esporas de FGC y CE, respectivamente, cuantificados mediante la técnica de tamizado y decantado en húmedo (Gerdemann y Nicolson 1963). El inoculo de la rizobacteria consistió en una cepa identificada como Arthrobacter spp. (AB), con número de accesión al GenBank, KX258420, el cual tiene la capacidad para producir auxinas (22.5 µg mL−1) y causar efectos antagónicos sobre el desarrollo de Phytophthora capsicí (45%) y Fusarium spp. (12%) (González-Mancilla et al. 2017). El inoculo bacteriano se preparó en medio liquido caldo nutritivo, incubándolo a 28 °C y en agitación constante por 72 horas, posteriormente se procedió a la inoculación, ocho días después de la germinación, agregando 1 mL de inoculo en medio caldo nutritivo que contenía 166 x 108 UFC mL−1.
Variables evaluadas
El experimento tuvo una duración de 80 días después de la germinación (DDG), bajo condiciones de invernadero. La altura y el número de hojas en plántulas, fueron determinados a los 10, 40 y 80 DDG. El área foliar, la producción de biomasa seca, la colonización micorrízica y la eficiencia fotoquímica del fotosistema II se evaluó al final del experimento (80 DDG). La altura se midió con una regla graduada en cm, desde la base del tallo hasta el meristemo apical de la plántula; el número de hojas se cuantificó de manera visual.
El área foliar se determinó con un medidor de área foliar marca LICOR (LI-3000, Inc.® Lincoln, Nebraska, USA), el método fue destructivo, al separar las hojas de los tallos. La producción de biomasa seca (raíz y aérea) se obtuvo al separar la parte aérea y radical de la plántula; las cuales se secaron en un horno marca FELISA®, Modelo 242-A (México) a 70 °C por 72 h; una vez finalizado el proceso, las partes de las plántulas se pesaron por separado en una balanza analítica (Sartorius®, Modelo Analytic AC 210S, Illinois, USA). La biomasa seca total, se calculó sumando los pesos secos de la raíz y de la parte aérea de las plántulas.
La colonización micorrízica en raíces de las plántulas de chile poblano, se evaluó utilizando la técnica de clareo y tinción con azul tripano propuesto por Phillips y Hayman (1970), y se expresó en porcentaje de colonización según el método de Biermann y Linderman (1981). La eficiencia fotoquímica del fotosistema II, que incluye la fluorescencia mínima (Fo), fluorescencia máxima (Fm), fluorescencia variable (Fv) y la eficiencia fotoquímica del fotosistema II (PSII) expresada en Fv/Fm y Fv/Fo, se midió con un fluorómetro OS-30p+ (Opti-Sciences®), considerando las lecturas en las hojas jóvenes y completamente desarrolladas.
Diseño experimental y tratamientos
Se utilizó un diseño experimental completamente al azar, con dos factores: tres niveles de HMA (FGC, CE y SM) y dos niveles de bacterias (AB y SB), teniendo seis tratamientos y 15 repeticiones cada uno, en total 90 unidades experimentales, siendo una unidad experimental cada plántula de chile poblano. Los tratamientos quedaron de la siguiente manera: T1) FGC + AB, T2) FGC + SB, T3) CE + AB T4) CE + SB, T5) SM + AB y T6) SM + SB (testigo sin microorganismos). Las variables fueron analizadas mediante el paquete estadístico SAS para Windows (SAS 2002), realizando un análisis de varianza y prueba de comparación de medias de Tukey (p ≤ 0.05).
Resultados
Crecimiento de plántulas
La inoculación de la micorriza arbuscular más la cepa bacteriana ejercieron efectos significativos (p ≤ 0.05) en altura de plántulas de chile poblano, según los diferentes tratamientos y diferentes fechas de evaluación. En la primera medición, 10 DDG, tres de los seis tratamientos evaluados [T1 (FGC + AB), T2 (FGC + SB) y T3 (CE + SB)], presentaron mayor altura de plántulas con 6.1 y 6.2 cm, no habiendo diferencias estadísticas significativas entre ellos. Después de los 40 DDG, los tratamientos T1 (FGC + AB), T2 (FGC + SB), T3 (CE + AB) y T4 (CE + SB) presentaron mayor altura (9.7, 9.5, 9.4 y 10 cm) siendo estadísticamente similares entre ellos y estadísticamente diferentes con los tratamientos T5 (SM + AB) y T6 (SM + SB), quienes obtuvieron una altura de 6.4 y 6.1 cm, respectivamente (Figura 1).
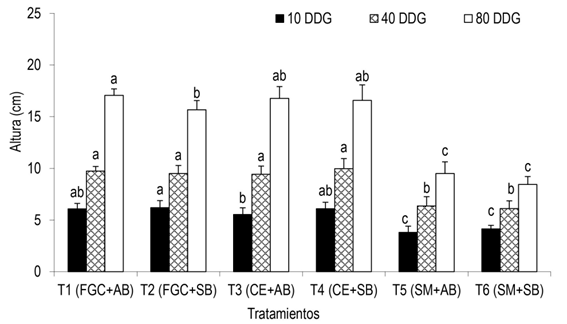
Figura 1: Altura en plántulas de chile poblano, inoculados con rizobacterias y hongos micorrízicos arbusculares. 10, 40 y 80 días después de la germinación (DDG). FGC = Funneliformis geosporum + Claroideoglomus spp.; CE = Claroideoglomus etunicatum; SM = Sin micorriza; AB = Arthrobacter spp.; y SB = Sin bacteria. n = 15, ± EE. Barras similares y con letras diferentes, indican diferencias estadísticas significativas según Tukey (p ≤ 0.05).
La altura de plántulas al final del experimento (80 DDG) presentó diferencias estadísticas significativas, el valor más alto se encontró en el T1 con 17.1 cm, seguido del T3 y T4 con 16.8 y 16.6 cm; los T5 y T6 presentaron en promedio 9.5 y 8.4 cm en altura, valores más bajos respecto a los demás tratamientos. El número de hojas por plántula a los 10 DDG fue relativamente similar, sobresaliendo el T4 con 4.2 hojas por plántula, a los 40 DDG sobresalieron las plántulas tratadas con el T1 con 8.3 hojas por plántula, este tratamiento presentó diferencias estadísticas significativas (p ≤ 0.05) respecto al número de hojas encontrado en el testigo (T6, 5.9 hojas plántula−1), no así con los demás tratamientos, quienes estadísticamente fueron similares (Tabla 1). A los 80 DDG, se observó mayores diferencias estadísticas significativas (p ≤ 0.05) entre los tratamientos, registrándose mayor número de hojas por plántula en los tratamientos T1 con 8.9 hojas plántula−1 y T2 con 8.1 hojas plántula−1; el tratamiento T6 obtuvo menor número de hojas, en promedio 5.3 hojas por plántula (Tabla 1).
Tabla 1: Número de hojas, área foliar y colonización micorrízica en plántulas de chile poblano inoculados con rizobacterias y hongos micorrízicos arbusculares.
| Tratamientos | Numero de hojas | Área foliar (cm2) | Colonización micorrízica (%) | ||
|---|---|---|---|---|---|
| 10 DDG | 40 DDG | 80 DDG | |||
| T1 = FGC+AB | 4.1(± 0.5) ab | 8.3(± 0.8) a | 8.9(± 0.7) a | 31.2(± 1.6) a | 16.7(± 2.3) a |
| T2 = FGC + SB | 4.0(± 0.0) ab | 7.7(± 0.9) a | 8.1(± 0.9) ab | 25.4(± 2.5) c | 7.9(± 1.3) b |
| T3 = CE + AB | 4.1(± 0.3) ab | 8.0(± 0.6) a | 7.8(± 1.3) b | 29.2(± 2.8) ab | 6.9(± 1.1) b |
| T4 = CE + SB | 4.2(± 0.5) a | 7.9(± 0.6) a | 7.0(± 1.0) bc | 26.6(± 5.1) bc | 5.1(± 1.3) b |
| T5 = SM + AB | 3.7(± 0.4) b | 7.5(± 0.9) a | 6.3(± 1.1) cd | 12.4(± 1.8) d | 0.0(± 0.0) c |
| T6 = SM + SB | 3.7(± 0.4) b | 5.9(± 0.8) b | 5.3(± 0.9) d | 8.6(± 1.8) e | 0.0(± 0.0) c |
FGC = Funneliformis geosporum + Claroideoglomus spp., CE = Claroideoglomus etunicatum, SM = Sin micorriza, AB = Arthrobacter spp., SB = Sin bacteria. n = 15, ± EE. Letras diferentes en las columnas, indican diferencias estadísticas significativas según Tukey (p ≤ 0.05).
El área foliar al final del experimento (80 DDG) fluctuó entre 8.6 y 31.2 cm2, los valores más altos (31.2 y 29.2 cm2) fueron localizados en T1 y T2, siendo estos estadísticamente similares (p ≤ 0.05) pero diferentes respecto a los demás tratamientos (Tabla 1). La menor área foliar, con 8.6 cm2, fue localizada en el T6 (Testigo), seguido del T5 con 12.4 cm2, éste último inoculados con Arthrobacter spp.
La producción de biomasa seca de raíz, parte aérea y biomasa seca total, en cada uno de los tratamientos, presentaron diferencias estadísticas significativas (p ≤ 0.05) (Figura 2). La mayor producción de biomasa seca radical (83 mg plántula−1), fue localizada en plántulas tratadas con T1, estadísticamente similar con los tratamientos T3 y T4, quienes obtuvieron 75 y 73 mg por plántula. Los tratamientos T5 y testigo T6 produjeron menor cantidad de biomasa seca radical, siendo 46 y 31 mg por plántula, respectivamente.

Figura 2: Producción de biomasa seca (radical, aérea y total) en plántulas de chile poblano, inoculados con rizobacterias y hongos micorrízicos arbusculares, 80 DDG. FGC = Funneliformis geosporum + Claroideoglomus spp., CE = Claroideoglomus etunicatum; SM = Sin micorriza; AB = Arthrobacter spp.; y SB = Sin bacteria. n = 15, ± EE. Barras similares y con letras diferentes, indican diferencias estadísticas significativas según Tukey (p ≤ 0.05).
La biomasa seca aérea fue mayor en los tratamientos T1 y T3, con 189 y 187 mg por plántula, siendo éstos, estadísticamente diferentes respecto a los demás tratamientos (Figura 2). Los valores más bajos fueron obtenidos en los tratamientos T5 y T6, con 70 y 52 mg de biomasa seca aérea por plántula. Los mismos tratamientos con T1 y T3, acumularon la mayor producción de biomasa seca total con 271 y 262 mg por plántula, mientras que el total en el tratamiento T6 fue de 83 mg plántula−1, y de 116 mg plántula−1 en las plántulas inoculadas con la cepa bacteriana (T5).
Colonización micorrízica
La colonización micorrízica, en las plántulas inoculadas, fluctuó entre 5.1 y 16.7% (Tabla 1). El T1 (FGC + AB) logró mayor colonización, siendo estadísticamente diferente respecto a los demás tratamientos, quienes presentaron 7.9 (T2), 6.9 (T3) y 5.1% (T4) de colonización micorrízica; mientras que los tratamientos T5 (SM + AB) y T6 (SM + SB) no presentaron colonización micorrízica.
Eficiencia fotoquímica del fotosistema II (PSII)
La medición de la eficiencia fotoquímica del PSII, basada en la fluorescencia de la clorofila, muestran diferencias estadísticas significativas entre los tratamientos (p ≤ 0.05) (Tabla 2). La Fluorescencia mínima (Fo) fluctuó entre 273 y 518, el valor más bajo fue localizado en el tratamiento con T1 y el más alto en el tratamiento con T5; la fluorescencia variable (Fv) osciló entre 454 y 661, el valor más alto fue encontrado en el T2, y los valores más bajos lo presentaron los tratamientos T6 (454) y T5 (484). La fluorescencia máxima (Fm) fue mayor en el T5 con 990 y la más baja se encontró en el T1 con 766. Finalmente, la eficiencia fotoquímica del PSII, expresada en Fv/Fm y Fv/Fo, fue más alta en el T2 con 0.70 y 2.32; seguido del T1 con 0.61 y 1.86. Los valores más bajos fueron localizados en los tratamientos T5 y T6 (testigo) con 0.49 y 0.54 de Fv/Fm y de 1.05 y 1.22 en Fv/Fo (Tabla 2).
Tabla 2: Eficiencia fotoquímica del fotosistema II en plántulas de chile poblano inoculados con rizobacterias y hongos micorrízicos arbusculares.
| Tratamientos | Fo | Fv | Fm | Fv/Fm | Fv/Fo |
|---|---|---|---|---|---|
| T1 = FGC + AB | 273(± 97) b | 492(± 227) a | 766(± 264) b | 0.61(± 0.14) ab | 1.86(± 0.84) ab |
| T2 = FGC + SB | 290(± 33) b | 661(± 40) a | 951(± 42) ab | 0.70(± 0.03) a | 2.32(± 0.37) a |
| T3 = CE + AB | 376(± 61) ab | 522(± 136) a | 898(± 160) ab | 0.58(± 0.06) ab | 1.41(± 0.38) b |
| T4 = CE + SB | 421(± 95) ab | 505(± 68) a | 927(± 75) ab | 0.55(± 0.08) ab | 1.30(± 0.52) b |
| T5 = SM + AB | 518(± 158) a | 484(± 135) a | 990(± 79) a | 0.49(± 0.13) b | 1.05(± 0.41) b |
| T6 = SM + SB | 391(± 76) ab | 454(± 87) a | 846(± 78) ab | 0.54(± 0.08) b | 1.22(± 0.38) b |
FGC = Funneliformis geosporum + Claroideoglomus spp.; CE = Claroideoglomus etunicatum; SM = Sin micorriza, AB = Arthrobacter spp., SB = Sin bacteria; Fo= Fluorescencia mínima; Fm = Fluorescencia máxima; Fv = Fluorescencia variable; Fv/Fm y Fv/Fo = eficiencia fotoquímica del PSII. n = 15, ± EE. Letras diferentes en las columnas, indican diferencias estadísticas significativas según Tukey (p ≤ 0.05)
Discusión
Crecimiento de plántulas
La inoculación de microorganismos nativos de la rizosfera pueden influir en el crecimiento de las plantas (Yadav et al. 2015). En este trabajo, las plántulas inoculadas con HMA y con la BPCV, solos o combinados, presentaron mayor crecimiento respecto a las plántulas no inoculadas; sin embargo, se observó mejor efecto cuando estos microorganismos se combinaron (T1 y T3). La inoculación de FGC + AB (T1), permitió mayor crecimiento de las plántulas de chile poblano, representando un incremento de 113, 68, 264 y 227% más en altura, número de hojas, área foliar y biomasa seca total comparado con el T6. Los efectos positivos de la inoculación con HMA y BPCV podrían estar relacionados con los mecanismos fisiológicos de ambos microorganismos, como favorecer el traslado de nutrientes (fósforo) y agua (Kong et al. 2020, Almaraz-Suárez et al. 2021), producir auxinas y solubilizar fosfatos (Sandhya et al. 2010, González-Mancilla et al. 2017). Diversos estudios han demostrado los efectos positivos al inocular HMA con BPCV, por ejemplo, FGC con Bacillus pumilus en chile pimiento incrementaron la altura de plántula, el área foliar y la biomasa seca (23%) (Angulo-Castro et al. 2021), Funneliformis mosseae con Ensifer meliloti incrementaron altura (28%) y biomasa seca (85%) en plantas de Vitis vinífera (Velásquez et al. 2020). Mientras que con F. mosseae, Acaulospora laevis y Pseudomonas fluorescens reportaron mayor altura (55%), área foliar (62%) y biomasa seca (133%) en plantas de chile bell pepper (Tanwar y Aggarwal 2014); A. longula, Glomus clarum y G. intraradiaces con Methylobacterium oryzae mejoraron la biomasa seca (17%) y altura (3%) de chile rojo (Kim et al. 2010). Mientras que con G. fasciculatum y Acinetobacter junii se reporta el incremento en la producción de biomasa seca (124%) en chile bell pepper (Padmavathi et al. 2015).
Colonización micorrízica
La colonización micorrízica en las raíces de las plántulas de chile poblano, se vio favorecida cuando se combinaron los HMA y la rizobacteria Arthrobacter spp. El T1 permitió un incremento de dos veces más el porcentaje de colonización micorrízica respecto al tratamiento solo con micorriza (T2), sin embargo, en el T3 dicho incremento fue de 1.4 veces más que el T4. Las rizobacterias producen diversos factores de crecimiento, mismos que pueden mejorar la colonización de las raíces por HMA, estimular la germinación de esporas y optimizar el desarrollo micelial (Frey-Klett et al. 2007). En otros trabajos, se ha observado un incremento de la colonización micorrízica cuando se inoculan juntamente con BPCV del género Bacillus, Pseudomonas, Bradyrhizobium, Burkholderia, Paenibacillus y Rhodococcus (Bending et al. 2006, Gamalero et al. 2008). El género micorrízico Funneliformis, puede asociarse con diversas comunidades bacterianas, principalmente con el género Arthrobacter del phylum Actinobacteria; dichas rizobacterias pueden desempeñar funciones específicas que afectan el rendimiento de los HMA, en términos de infectividad y eficiencia (Agnolucci et al. 2015). En este trabajo Arthrobacter spp., permitió incrementar el porcentaje de colonización en las plántulas con el T1, pero no fue efectiva con plántulas del T3.
Eficiencia fotoquímica del PSII
La medición de la eficiencia fotoquímica del PSII basada en la fluorescencia de la clorofila es una técnica efectiva para detectar daños en el PSII (Baker 2008). Los parámetros Fv/Fm y Fv/Fo predicen el desarrollo de la planta (Ruban 2015). En este trabajo, Fv/Fm y en Fv/Fo, en plántulas de chile poblano, fueron mejores en los tratamientos inoculados con HMA, solos o combinados con la rizobacteria Arthrobacter spp. El T2 presentó 30% más Fv/Fm, sin embargo, fue más baja cuando se combinó con Arthrobacter spp. (T1), al incrementar únicamente el 13% más que el T6. En cuanto a Fv/Fo, el incremento fue del 90% cuando se inoculo con T2, pero combinada con Arthrobacter spp. (T1), el incremento fue del 53%. Los valores altos en Fv/Fm y Fv/Fo posiblemente se relacionen con que las plántulas no hayan sufrido estrés y tengan un contenido nutrimental adecuado (Moreno et al. 2008). Existen pocos estudios sobre estas variables, referentes al uso de HMA y BPCV en cultivos de chile, al respecto, Angulo-Castro et al. (2018) en chile jalapeño, y al inocular (por separado) diversas especies de HMA (incluyendo FGC de este trabajo) y BPCV, no encontraron diferencias significativas y fue mejor un tratamiento fertilizado con más de 3 Fv/Fo, las plantas con FGC reportaron 2.6 Fv/Fo, valores más altos que los encontrados en este trabajo. Así mismo, Angulo-Castro et al. (2021) reportaron incrementos del 21.5% en fv/fo cuando se inoculó bell pepper con FGC, pero cuando se combinó FGC con Pseudomonas tolaasii fue 37% más el valor comparado con el testigo.
Conclusiones
El crecimiento de plántulas de chile poblano, a nivel semillero, se mejoró con la inoculación de los HMA y las BPCV, solos o en combinación, respecto a las plántulas no inoculadas. El tratamiento de Funneliformis geosporum + Claroideoglomus spp. + Arthrobacter spp. mejoró la altura, el número de hojas, área foliar, colonización micorrízica y producción de biomasa seca en plántulas. Los consorcios micorrízicos ejercieron efectos positivos en la eficiencia fotoquímica del fotosistema II, siendo mejor el tratamiento de Funneliformis geosporum + Claroideoglomus spp. Obtener plántulas sanas y de mejor crecimiento, incrementa la oportunidad de éxito cuando éstas son trasplantadas en campo, y al mismo tiempo inciden en las características de las plantas en etapas posteriores.

